
高原2种草本植物的光合作用和叶绿素荧光参数日动态
2015-12-16 07:44:50高丽楠陈舒慧孙佳媛
四川师范大学学报(自然科学版) 2015年4期
高丽楠, 张 宏, 陈舒慧, 贺 琼, 孙佳媛
(1.成都大学旅游文化产业学院,四川成都610106;2.四川师范大学资源生态学研究中心,四川成都610066)
青藏高原属于气候变化的敏感区和生态脆弱带,在全球气候变化背景下呈现出暖化趋势[1].青藏高原的沼泽化草甸面积4.9×104km2,占整个青藏高原草地面积的1.96%,是青藏高原分布面积较广的草地生态系统之一[2].作为青藏高原较为典型的植被类型,高寒沼泽草甸是适应高原隆起和长期低温环境形成的特殊产物,也是牧民的主要牧草来源.但其生态系统极其脆弱,对人类干扰和由温室气体增加引起的全球气候变化极为敏感,一旦遭受破坏很难在短期内得到恢复,并可能导致草地退化和沙漠化发生,生态环境日趋恶化.在全球变化情景下,高寒沼泽草甸土壤层的碳释放和植被层的碳吸收对全球变化更为敏感,因而在区域生态系统碳平衡过程中起着极为重要的作用[3].然而,光合作用是植物生长发育的基础和生产力高低的决定性因素,光合作用过程中所吸收的CO2是陆地生态系统碳循环的入口,并对减缓全球温室效应具有重要作用[4],同时也是一个对外界环境变化非常敏感的生理过程.因此,深入研究高寒沼泽草甸的光合特性是当前急需开展的工作.近年来,高寒草甸植物光合作用的研究相对较少[5-6],并且尚未见到系统探讨青藏高原高寒沼泽草甸光合特性的报道.
川西北高寒沼泽草甸位于青藏高原东缘,是我国主要牧区之一.川西北高寒沼泽草甸不仅是四川省重要的水源地,而且是黄河上游重要的水源涵养区和集水区.目前川西北草地退化面积已达60%,出现草地生产力降低、水土流失严重等多种生态系统退化症状,生态系统失衡的问题十分突出[7],其中,四川嵩草和木里苔草是川西北高寒沼泽草甸植物群落的优势种,其良好生长对维持青藏高原的生态平衡起着重要作用,但对其生理生态学方面的研究较少.所以,本文重点讨论青藏高原东缘高寒沼泽草甸2种优势草本的光合作用和叶绿素荧光参数的日动态,以探讨高寒环境对植物光合作用的影响及高山植物光合特性,也为该地区植被恢复与重建提供理论依据.
1 材料与方法
1.1 实验样地 实验样地位于四川省阿坝州红原县(101°51'-103°23'E,31°51'-33°19'N,平均海拔3 600 m),地处青藏高原东部边缘.气候属大陆性高原气候,日温差大、霜冻期长,年平均气温1.1℃,年平均相对湿度60% ~70%;日照时间长,太阳辐射强,年均日照时间2 158.7 h,太阳辐射年总量为6 194 MJ/m2[8].草甸和灌丛是当地的主要植被类型,草甸分为带有高寒性质的亚高山灌丛草甸、亚高山草甸和沼泽草甸类型[9].区内土壤类型以亚高山草甸土、沼泽土和高山草甸土为主,其中亚高山草甸土占全县土地总面积的55.4%[10].
测定点位于红原县城以北10 km的平坦宽谷中,选取了一块约400 m2的沼泽草甸样地(102°32'E,33°03'N;海拔:3 514 m 左右),该草地为季节性积水,积水面积为30% ~40%,雨季积水2~5 cm.群落优势种主要有:四川嵩草(Kobresia setchwanensis Hand.-Mazz)、西藏嵩草(Kobresia tibetica Maxim)、木里苔草(Carex muliiensis Hand.-Mazz)、乌拉苔草(Carex meyeriana Kunth)等,主要物种见表1.样地群落总盖度在90%以上[11],土壤类型为泥炭沼泽土,成土母质以河湖沉积物为主.

表1 样地的主要物种组成Table 1 The composition of plant species at sampling sites
1.2 测定方法及数据处理
1.2.1 光合作用测定 野外实验选在草本生长旺季的7月8-16日进行.选择晴朗无云的天气,7:00-19:00,每2 h测定一次.分别随机选取8株生长良好的木里苔草和四川嵩草,选取植株的中上部向阳的成熟的叶片重复测定2次,共16个数据取平均值.由于无法在同一时间同时测定2种草本,所以采取每株植物轮流测定的方法,即第1次以木里苔草→四川嵩草的顺序来测定,第2次就以相反的顺序来测定,第3次的测定顺序又与第1次的相同,以此类推,以消除2种草本测定时间上的误差.
用LI-6400光合测定仪测定叶片的光合速率(Pn)和蒸腾速率(Tr),并同步记录光合参数:有效辐射强度(PAR)、大气 CO2浓度(ca,)、气温(Tair)、大气相对湿度(RH)、叶片气孔导度(Gs)、胞间CO2浓度(ci)和叶片温度(Tleaf).
最后根据以上参数,计算出叶片水分利用效率(WUE=Pn/Tr)[12]和气孔限制值 (Ls=1-ci/ca)[13].木里苔草的叶面积使用AM300叶面积仪进行测定;四川嵩草的叶面积采用几何计算方法进行测定,先使用游标卡尺测量针叶的平均直径和长度以获取总表面积,再乘以0.5得到半球表面积[14].
1.2.2 叶绿素荧光参数测定 与光合作用同步测定,测定方式同理于光合测定.利用 PAM-2100(Walz,Germany)便携式叶绿素荧光仪测定叶绿素荧光参数,叶片暗适应30 min后,打开弱测量光测定初始荧光(Fo),再打开一次饱和脉冲光(PFD为8 000 μmol·m-2·s-1,频率为 20 kHz,0.8 s,1 个脉冲)测定最大荧光(Fm).然后在自然光下,光下稳态荧光(Ft)稳定后,再打一次饱和脉冲光测定光下最大荧光(F'm).得出光系统II(PSII)的反应中心
原初光能转化效率为
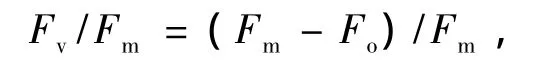
实际光化学反应效率为
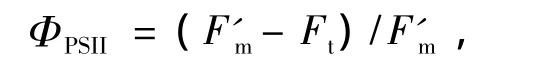
和电子传递速率[15]为
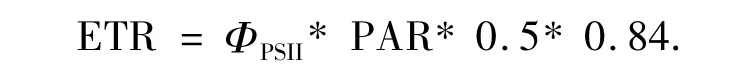
电子传递速率响应曲线的测量,7:00-19:00,每2 h测定一次.测量光为波长650 nm的红光,强度为0.1 μmol·m-2·s-1.光化光和饱和脉冲光均为卤素灯发出的白光,其中饱和脉冲光的强度大于5 000 μmol·m-2·s-1,脉冲时间 0.8 s.为避免外界光对植物生理状态的影响,作电子传递速率响应曲线时用小黑布罩住植物[16].荧光测量由电脑控制,步骤为:打开测量光并打开光化光,适应10 s后打开饱和脉冲光,升高光化光强度(升高1),适应10 s后再打开饱和脉冲,如此重复5次.
将测定的电子传递速率响应曲线通过拟合[17],得到最大电子传递速率(ETRmax)、光能利用效率(α)、半饱和光强(Ek).拟合方程式为
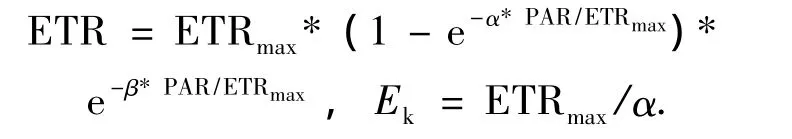
用SPSS12和Originpro 7.5软件对所得实验数据进行统计、分析和图形处理.
2 结果与分析
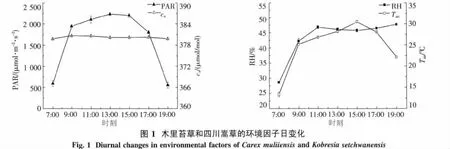
2.1 环境因子日变化规律 图1表明,光合有效辐射(PAR)的日变化呈单峰曲线,最高峰出现在13:00左右,此时PAR值达到2 237.43 μmol·m-2·s-1,日平均 PAR 为 1 634.43 μmol·m-2·s-1.大气CO2浓度(ca)基本保持不变,日均值为380.2 μmol/mol.相对湿度(RH)则在11:00达到峰值,此时RH为46.87%,其后随着光合有效辐射、气温的升高而逐渐下降,15:00降至最低,之后随着光合有效辐射和气温的下降又略有升高.气温(Tair)主要受PAR的影响,7:00左右为最低值(13.33℃),此后随着光合有效辐射的增加而迅速上升,到15:00达到峰值(30.50℃),其峰值比PAR略滞后,日平均气温为24.88℃(图1).
2.2 光合速率及其参数日变化特征
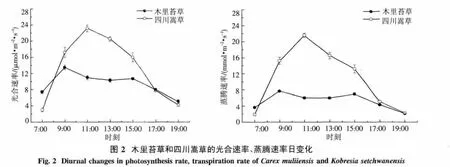
2.2.1 光合速率和蒸腾速率日变化特征 木里苔草的Pn日变化表现为典型的双峰曲线,有明显的“午休”现象,第一个峰值出现在9:00左右,为13.5 μmol·m-2·s-1,第二个峰值出现在15:00左右,为 10.7 μmol·m-2·s-1,在 13:00 左右出现低谷(图2).由此可以看出,木里苔草Pn的第二个峰值低于第一个峰值,且中午Pn下降的速率均大于下午Pn回升的速率,即Pn中午降低快而恢复慢.四川嵩草的Pn日变化表现为典型的单峰曲线,在11:00达到峰值,为 23.2 μmol·m-2·s-1,而后又迅速下降.木里苔草与四川嵩草的Pn日平均值之间存在极显著差异(P<0.01).2种草本所处的环境条件相同,而Pn日动态曲线表现出不同规律,四川嵩草Pn上升和下降速率均大于木里苔草,说明植物光合速率主要由本身的生理生态特性所决定.Tr日变化趋势和Pn日变化相似,木里苔草Tr表现为双峰曲线,而四川嵩草依然为单峰曲线,嵩草Pn上升和下降速率均大于木里苔草.木里苔草Tr分别在9:00及15:00左右出现峰值;而四川嵩草Tr出现在11:00左右,木里苔草与四川嵩草的Tr日平均值之间存在极显著差异(P<0.01).
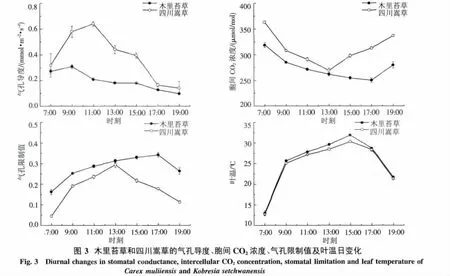
2.2.2 气孔导度和胞间CO2浓度日变化特征 四川嵩草Gs日变化表现为单峰曲线,从早晨开始随着PAR和Tair的增加,Gs逐渐增加,到11:00左右出现峰值,之后由于高温低湿的环境导致Gs迅速降低.但木里苔草Gs日变化也表现为微弱的双峰态势,9:00左右出现最高值,9:00-13:00呈递减趋势,Gs较低且日变化波动较小.四川嵩草的Gs日平均值极显著高于木里苔草(P<0.01).
四川嵩草的胞间CO2浓度(ci)在一天中呈早晚高、中午低的近“V”型变化趋势.7:00为363.0 μmol/mol,随着Pn的加强,叶片固定较多的CO2致使ci迅速下降;13:00左右达到低谷,之后一直呈上升趋势,到19:00达到336.9 μmol/mol.但木里苔草的ci的日变化规律表现出7:00最高,之后逐渐降低,在17:00达到全天的最低值.四川嵩草的ci日平均值极显著高于木里苔草(P<0.01).
2.2.3 气孔限制值及叶温日变化 四川嵩草和木里苔草的Ls日变化都呈现出单峰型曲线,2个峰值分别出现在13:00左右和17:00左右.木里苔草Ls在17:00之前保持缓慢上升,之后逐渐下降.而四川嵩草Ls在13:00之前急剧增大,之后迅速降低(图3).从Ls的日平均值的大小看,与ci表现出相反的趋势,木里苔草Ls显著高于四川嵩草(P<0.01).
木里苔草和四川嵩草的叶面温度日变化规律与气温日变化完全一致,日均叶温分别为25.5℃和24.8℃.木里苔草(r=0.998,P<0.01)和四川嵩草(r=0.891,P<0.01)的叶面温度都与大气温度呈极显著正相关,所以叶片温度主要受气温影响.
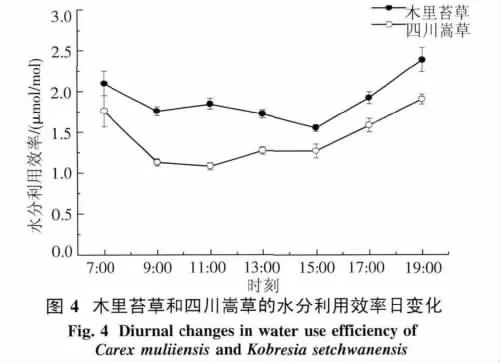
2.2.4 水分利用效率日变化 木里苔草WUE在7:00最大,为 2.1 μmol/mol,之后随着光合有效辐射和叶温的增加而降低,9:00-15:00,木里苔草WUE 在1.6~1.8 μmol/mol范围内波动,15:00之后又迅速上升.四川嵩草WUE日变化呈早晚高,中午低的趋势,7:00表现为最大,11:00降至最低,11:00-15:00都较为平稳的波动,15:00又迅速上升(图4).木里苔草的WUE日平均值极显著高于四川嵩草(P<0.01).
2.3 光合速率与生理生态因子的相关性分析
2.3.1 木里苔草净光合速率与生理生态因子的关系 木里苔草净光合速率与生理生态因子通径分析和相关分析结果(表2和表3)表明:从相关系数的大小看来,生理生态因子对木里苔草的Pn影响的大小顺序为:Tr>PAR>Gs>Tleaf>Tair>ca>ci>RH,其中,Tr、PAR、Gs、Tleaf、Tair、ca与 Pn呈极显著正相关,而ci与Pn呈显著的负相关.各生理生态因子对净光合速率直接通径系数的大小依次为:Tr>RH>Gs>PAR>ca>Tair>Tleaf>ci;Tr正向的直接作用远大于其对Tair和Tleaf的负向间接作用,故表现为其与Pn呈显著的正相关性.生理因子的决策系数顺序为:R2(Tr)>R2(Gs)>R2(PAR)>R2(ca)>R2(RH)>R2(ci)>R2(Tair)>R2(Tleaf),其中,R2(RH)、R2(ci)、R2(Tair)和 R2(Tleaf)均小于 0.所以,影响木里苔草Pn日变化规律的决定生理生态因子为Tr、Gs、PAR 和 ca,其中 Tr为主要决定因子;限制因子为 RH、ci、Tair和 Tleaf,其中,Tleaf为主要限制因子.
2.3.2 四川嵩草净光合速率与生理生态因子的关系 四川嵩草净光合速率与生理因子通径分析和相关分析结果(表3 和表 4)表明:PAR、ca、RH、Tair、Gs、Tr、Tleaf与 Pn呈极显著正相关,而 ci与 Pn呈显著的负相关.各生理因子对净光合速率直接通径系数的大小依次为:Tr>Gs>PAR>RH>ca>Tair>Tleaf> ci;尽管 ci通过 ca、RH、Tair和 Tleaf对 Pn的间接作用均为正值,但ci直接作用(-0.431)远高于这些间接作用,所以没有影响ci与Pn呈极显著负相关;Tleaf对Pn的直接作用不大(-0.138),但被Tr正间接作用(0.483)所掩盖而使其与Pn正相关.生理生态因子的决策系数顺序为:R2(Tr)>R2(ci)>R2(Gs)>R2(PAR)>R2(RH)>R2(ca)>R2(Tair)>R2(Tleaf),其中,R2(ca)、R2(RH)、R2(Tair)和R2(Tleaf)小于0.所以,影响木里苔草Pn日变化规律的决定生理生态因子为Tr、Gs、PAR和ci,其中,Tr为主要决定因子;Tleaf为主要限制因子.
2.4 叶绿素荧光参数
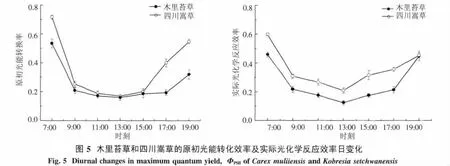
2.4.1 原初光能转化效率和实际光化学反应效率由图5看出,2种草本植物Fv/Fm和ΦPSII的日动态趋势相似,7:00最高,并随着光合有效辐射的增强而逐渐降低,在13:00降至最低,表明此时发生了光抑制现象;下午随着光强的减弱又逐渐升高,说明其光抑制到下午又逐渐得到恢复,19时2种草本Fv/Fm并未恢复到7:00的水平.木里苔草13时Fv/Fm相对于其7:00降低了70.3%,而四川嵩草13时Fv/Fm相对于其7:00降低了76.5%.早晚时,四川嵩草与木里苔草Fv/Fm之间存在极显著差异(P<0.01),表明四川嵩草叶片PSII的潜在活性和原初光能转换效率均高于木里苔草;而且四川嵩草ΦPSII日平均值极显著高于木里苔草(P<0.01).
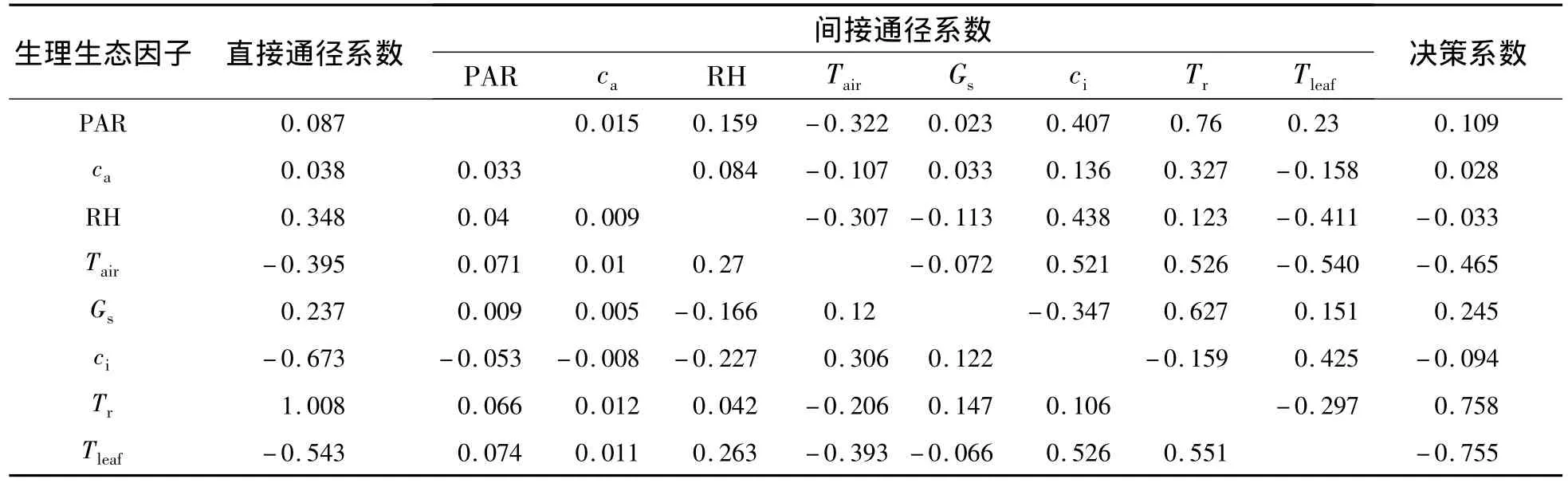
表2 木里苔草的净光合速率与生理生态因子通径系数Table 2 Path coefficients of Pnand eco-physiological factors in Carex muliiensis
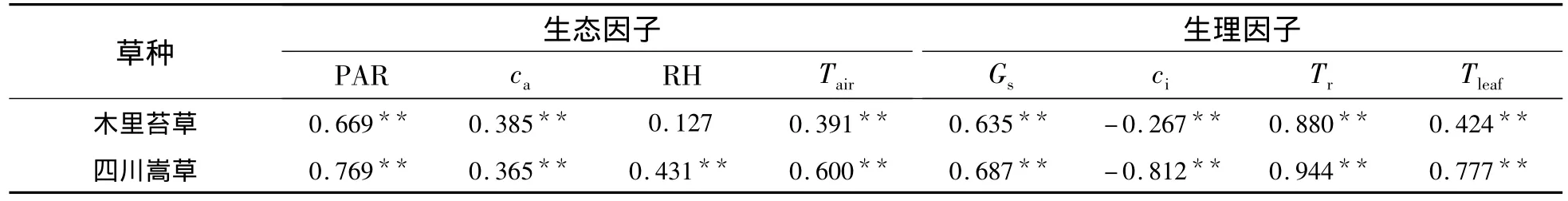
表3 净光合速率与生理生态因子相关系数Table 3 Correlation coefficients of Pnand eco-physiological factors
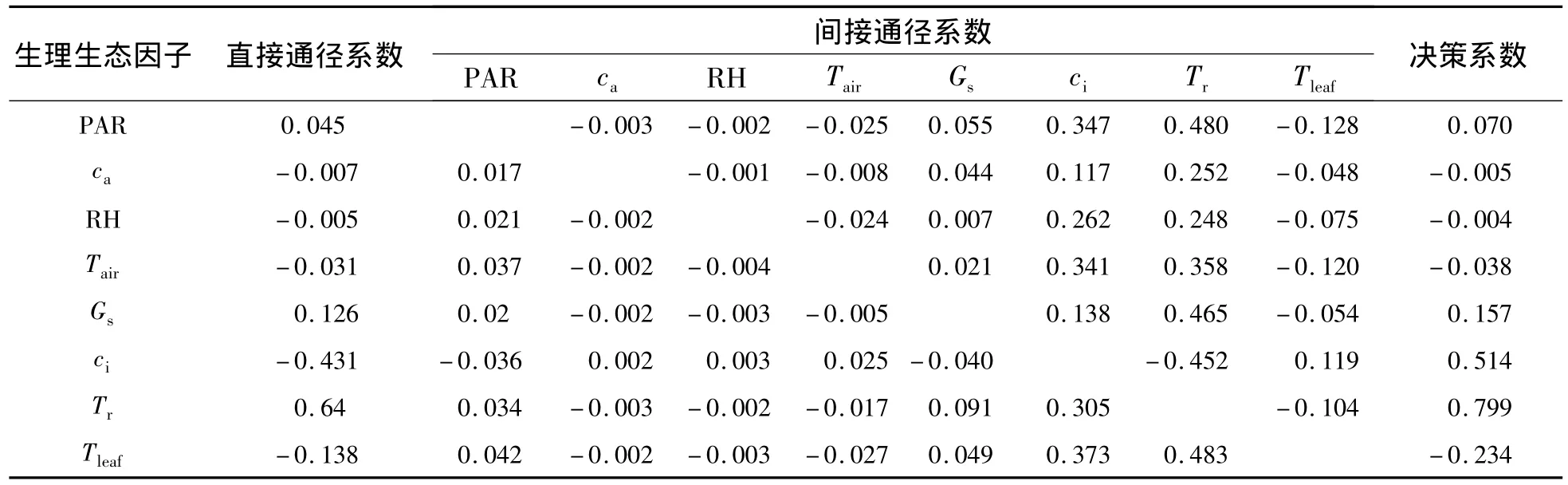
表4 四川嵩草的净光合速率与生理生态因子通径系数Table 4 Path coefficients of Pnand eco-physiological factors in Kobresia setchwanensis
2.4.2 快速光响应曲线 四川嵩草的ETRmax与Pn日变化相似,都呈单峰曲线,在11:00达到峰值,之后ETRmax逐渐降低.木里苔草ETRmax日变化呈微弱双峰曲线,第一个峰值出现在9:00左右,之后降低,在13:00左右出现低谷,随后略有回升,第二个峰值出现在15:00左右.另外,四川嵩草和木里苔草的Ek日变化都呈单峰曲线,分别在11:00和9:00达到峰值(图6).四川嵩草的ETRmax和 Ek日平均值都极显著高于木里苔草(P<0.01).
四川嵩草的α一天中呈早晚高、中午低的近“V”型变化趋势,11:00降至最低,之后逐渐上升.但木里苔草α日变化呈“W”型变化趋势,7:00-11:00较高且波动较小,之后降低,13:00左右出现低谷,随后再上升,15:00出现峰值,之后呈现再下降及上升的趋势.
3 讨论
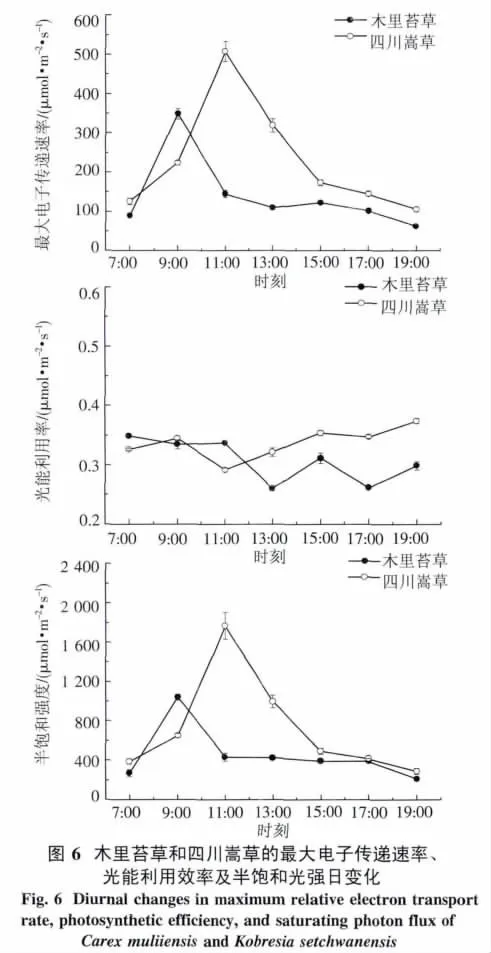
3.1 木里苔草与四川嵩草的光合特性 本试验四川嵩草Pn、Tr和Gs的日变化都呈单峰型曲线,最高峰均出现在11:00,由于11:00以前,土壤水分较充足,高原PAR强且增加较快,植物Pn迅速提高,植株通过增大气孔使外界CO2迅速向细胞内扩散以满足光合作用的需要,使得Gs增大,蒸腾速率加快;11:00以后随着光照强度的增强,过强的植物蒸腾和土壤蒸发量大[18],使植物体剧烈失水,而四川嵩草为浅根植物[19],植物根系得不到充足的供应,产生一定的信息物质(如脱落酸等),使叶片的气孔开张度减少蒸腾失水[20-21],Pn也随之下降,始终处于较低水平,所以四川嵩草Pn、Tr和Gs的日变化均呈单峰曲线,这与三江平原湿地毛苔草(Carex lasiocarpa Ehrh)研究结果相一致[22].但是,木里苔草Pn和Tr的日变化均呈典型的“双峰”曲线.日出之后,随着 PAR、Tair、Gs的逐渐增加,9:00左右达到最大值,之后强光照使叶片蒸腾失水过多,当在水分供应不充足时植物体多采用关闭气孔而减少水分散失,气孔的关闭造成ci降低,迫使光合速率下降[23],出现了光合“午休”现象.午后,由于光照减弱,使气孔导度略有回升,Pn在15:00达到第二次高峰,这也受益于它们具有发达的根系[24],与四川嵩草相比具有较高的WUE,可以充分吸收土壤水分补充蒸腾失水来满足生长需要,所以植物叶子的水分是对气孔的开闭有着强有力的控制作用.
木里苔草和四川嵩草Pn的日变化都呈现出光合“午休”现象,这与青藏高原东北隅矮嵩草(Kobresia humilis)[25]、老芒麦(Elymus sibiricus)[26]以及青海高原海北高寒草甸的垂穗披碱草(Elym usnutans)的研究结果相一致.光合作用的“午休”是2种草本植物在高寒沼泽草甸形成的对付环境胁迫的一种表现,这避免了植物的过度失水和保护了光合机构,并且加强了热耗散过程的运作[27].光合作用除了受到环境因素的影响外,其光合和蒸腾的大小其实还受到气孔导度的影响[28].植物叶片Pn中午降低的植物自身因素不外乎2个:即气孔因素和非气孔因素.前者是由于中午光照、温度和湿度等环境因子的变化引起植物气孔的部分关闭,CO2进入叶片受阻而使光合下降,而后者是由于叶肉细胞自身羧化酶活性的下降而引起Pn的降低[29].根据G.D.Farquhar等[30]的观点,只有当 Pn和 ci变化方向相同,两者同时减小,且Ls增大时,才可以认为Pn的下降主要是由Gs引起的,否则Pn的下降要归因于叶肉细胞羧化能力的降低.木里苔草在9:00-13:00期间Pn降低、ci下降、Ls增大,因此可以推断出引起光合“午休”的主要原因是气孔因素;午后15:00-17:00期间,Pn的下降主要也受气孔因素限制.而四川嵩草在11:00-13:00阶段,Pn与ci都下降,Ls上升,说明Pn的下降受气孔因素限制;13:00-19:00阶段,Pn的下降伴随着ci上升,Ls下降,说明Pn的下降主要受非气孔因素限制.研究表明,植物水分亏缺时光合速率的降低并不是由于水分供应不足直接引起的,而是由水分亏缺造成的气孔或非气孔因素限制,轻度水分亏缺以气孔限制为主,而较重水分胁迫下则以非气孔限制为主[31].说明气孔关闭对水分亏缺较敏感,木里苔草主要受到轻微水分胁迫,较低的Tr和较高的WUE显示出在受水分胁迫的高温低湿环境下的适应性,所以午后由于光照减弱,Pn再升高达到第二次高峰,日变化呈典型的“双峰”曲线.同理,四川嵩草主要受到较重水分胁迫,因受强烈PAR影响,植物蒸腾和土壤蒸发量大,而四川嵩草植物又多为浅根系植物,故为减少水分蒸发将气孔开度逐渐减小[32],Pn也随之下降,并难以恢复到较高水平,日变化呈典型的“单峰”曲线.所以,水分供应状况对2种植物的光合作用非常重要,而植物蒸腾是植物调节体内水分平衡的主要环节,植物通过调节蒸腾作用使光合作用最大化,减少水分散失并降低叶面温度、免受强光的灼伤[33],本研究结果也表明,2种草本植物光合作用Pn的主要决定因子是Tr.
与四川嵩草相比,木里苔草的WUE较高,说明生产相同量的物质消耗水分数量较少.木里苔草是沼泽湿地莎草科植物,多生长于沼泽草甸及低湿草地中,它兼具有水生和陆生植物共有的某些特征,适应性广,也有发达的根茎[34],长期适应沼泽草甸的环境中生长,可能使其具有较高的WUE.有研究表明,莎草科植物普遍具有较高的WUE[35],在生长过程中高效地使用了水分,进而保证了它们在沼泽湿地中作为优势种的地位[36].另外,本研究发现,2种草本植物WUE日变化均呈现早晚高,中午低的趋势,这是因为研究区早晨气温低,植物能以较低的水分代价得到更多的光合产物所致.随着温度升高,叶片气孔导度和蒸腾速率增加,这是对高温和强辐射的适应性策略,但却显著降低了光合速率和水分利用效率,这与内蒙古锡林浩特典型草原的羊草(Leymus Chinensis)具有相似的特征[37].
2种草本植物Pn、Gs和PAR的日变化趋势基本一致(图1~3),Pn与 PAR和 Gs呈正相关(表3).Pn与PAR的相关性大于与Gs的相关性(表3),而 Gs受光的诱导[38-39],可以认为生态因子PAR是影响2种草本植物光合作用的重要因子,这与青海高寒草甸的麻花艽(Gentiana straminea Maxim)研究结果相一致[40].低光强下随光强的升高Gs[39]、光合作用暗反应的酶活性升高[38],光合传递的电子增多(图6),光合作用加快.Pn与ci呈负相关,表明光合碳固定随光强升高而增加的速度大于Gs随光强增加的速度,导致ci随光强的升高而降低(图3),即强光下ci降低是由于光合作用利用CO2增加的结果.
3.2 木里苔草与四川嵩草的叶绿素荧光特性Fv/Fm反映了PSII反应中心潜在光能转化效率,即PSII反应中心处于开放状态时的量子产量.正常生长的植物的Fv/Fm值通常约为0.8~0.85之间[41],当植物受到水分胁迫、极端温度和盐胁迫等逆境胁迫后Fv/Fm值会不同程度下降,因此,Fv/Fm是逆境生理研究的重要指示性参数.本研究结果显示,清晨7:00,木里苔草和四川嵩草的Fv/Fm都低于上述范围,所以认为清晨环境对二者的生长构成抑制或胁迫,发生了长期的光抑制.这是因为,高原地区清晨的低温使植物的光合机构对光抑制更加敏感,低温下较低的光强也会引起光抑制[42].而且,四川嵩草7时Fv/Fm显著高于木里苔草,清晨时木里苔草光抑制程度更大,可能导致木里苔草的Pn上升速率较四川嵩草缓慢.四川嵩草Fv/Fm值高于木里苔草,也表明四川嵩草的光能转化效率也更高,适应环境变化的潜力更强,高的光化学转化效率有利于其维持较高的能量消耗和适应气温的急剧变化,因此四川嵩草的ETRmax较高.本研究结果表明,2种草本植物的Fv/Fm随日照时间的延长会不断降低,表明这2种植物遭受光抑制,光照越强光抑制现象越严重.13:00,四川嵩草Fv/Fm相对于其7:00降低了76.5%(图5),与木里苔草相比光抑制程度更大,植物水分胁迫较重,从而导致其光合速率的迅速降低.另外,19时2种草本Fv/Fm未恢复到7:00的水平,表明PSII反应中心可能发生了不可逆失活,或光合机构受到了破坏或损伤[43].这可能与青藏高原独特环境有关,低温和强光并存的环境条件往往造成植物较严重的光抑制[44].进一步说,在遭受低温的情况下,大多数植物的光抑制恢复较慢[45],是由于修复 PSII的 D-1蛋白质速率降低[46].本研究的通径分析也表明,2个草本植物的Pn,Tleaf为主要限制因子(表3和表4).
ΦPSII反映的是光系统II反应中心部分关闭情况下的实际光化学效率,与碳同化反应的强度密切相关.在中午强光、较高气温、低湿胁迫下,ΦPSII明显下降,午后又都有所恢复,说明强光胁迫使2种草本植物开放的光系统II反应中心部分关闭,光能转换效率下降,从而降低了电子传递能力,导致用于CO2同化的光能减少.其次,从7:00-19:00,四川嵩草的ΦPSII大于木里苔草,说明PSII的实际光能捕获效率较高,能够把所捕获的光能更多地用于光化学反应[47],因此四川嵩草的光合能力ETRmax较高.
PSII的相对光合电子传递速率(ETR)是反映实际光强下的表观电子传递速率,用于度量光化学反映导致碳固定的电子传递情况.将测定的电子传递速率响应曲线通过拟合,得到最大电子传递速率(ETRmax)、光能利用效率(α)和半饱和光强(Ek)的日变化.四川嵩草Ek显著大于木里苔草,说明其耐受强光.另外,2种草本植物ETRmax和Ek的变化趋势一致.这是因为Ek反映了植物耐受强光的能力[48],而耐受强光能力强的植物具有较高的最大电子传递速率ETRmax,所以四川嵩草有更大的光适应范围和较强的耐受强光能力,四川嵩草的ETRmax和Pn较木里苔草晚1 h到达峰值.由图6看出,从7:00-19:00,2个草本植物的Ek一直低于PAR,可见木里苔草一天中有12 h处在光饱和状态.本研究表明,2种草本植物光能利用效率α日变化趋势表现为清晨和傍晚较高,正午较低.这是因为,α反映了叶片捕光能力的高低,在中午光强太高,为了避免吸收的光能太多造成损伤,植物可以通过降低捕光能力α来保护自身,说明有调控光合作用和高效利用强光的能力.PAR最强13:00,四川嵩草α高于木里苔草,这说明上午光合有效辐射较强时,四川嵩草α较高,对强光耐受能力强.
4 结语
综上所述,川西北高寒沼泽草甸2种植物的光合生理特征之间有着较大的差异,这些差异在一定程度上是对外界变化生境适应的结果.木里苔草Pn和Tr的日变化呈双峰曲线,而四川嵩草则呈单峰曲线,两者都呈现光合“午休”现象,木里苔草的光合“午休”主要受气孔因素限制,但四川嵩草主要受非气孔因素限制.沼泽湿地植物木里苔草,它兼具有水生和陆生植物共有的某些特征,也有发达的根茎,长期适应沼泽草甸的环境中生长,较低的Tr和较高的WUE显示出在受水分胁迫的高温低湿环境下的适应性;但高寒草甸四川嵩草具有较高实际光化学反应效率(ΦPSII)和半饱和光强(Ek),表明四川嵩草对强光耐受能力强,高原沼泽草甸不同植物即使在相同环境条件下具有不同适应对策.2种草本植物光合作用Pn的主要决定因子是Tr,高寒沼泽草甸2种植物光合对环境的适应取决于水分状况,水分是植物生长的主要因子.过多的土壤水分在生长季节通过植物蒸腾的散失,从而使土壤含水量进一步减少,这在某种程度上加剧高寒沼泽湿地的退化.所以要保护和恢复湿地资源,合理利用湿地的植物资源,以及在已经退化的湿地中补充高WUE的土著湿地物种,都是在现有条件下保护和恢复湿地的策略.
致谢 成都大学校青年基金项目(2013XJZ23)对本文给予了资助,谨致谢意.
[1]Wu Q B,Li X,Li W J.The prediction of permafrost change along the Qinghai-Tibet highway,China[J].Permafrost and Periglacial Processes,2000,11(4):371-376.
[2]王俊峰,吴青柏.青藏高原沼泽草甸区植被退化对浅层寒冻土壤环境的影响[J].兰州大学学报:自然科学版,2011,47(6):39-45.
[3]徐洪灵,张宏,张伟.川西北高寒草甸土壤呼吸速率日变化及温度影响因子比较[J].四川师范大学学报:自然科学版,2012,35(3):405-411.
[4]张新时,周广胜,高琼,等.中国全球变化与陆地生态系统关系研究[J].地学前缘,1997,4(1/2):137-144.
[5]卢存福,卉桂英.矮嵩草光合作用与环境因素关系的比较研究[J].植物生态学报,1995,19(1):72-78.
[6]Cui X Y,Niu H S,Wu J,et al.Response of chlorophyll fluorescence to dynamic light in three alpine species differing in plant architecture[J].Environmental and Experimental Botany,2006,58:149-157.
[7]张宏,张伟,徐洪灵.川西北高寒草甸生长季土壤氮素动态[J].四川师范大学学报:自然科学版,2011,34(4):583-588.
[8]四川省红原县志编撰委员会.红原县志[M].成都:四川人民出版社,1996.
[9]刘长秀,张宏,泽柏.灌丛对川西北高寒草甸土壤资源的影响[J].山地学报,2006,24(3):357-365.
[10]田应兵,熊明标,宋光煜.若尔盖高原湿地土壤的恢复演替及其与养分变化[J].生态学杂志,2005,24(1):21-25.
[11]张伟,张宏.青藏高原东缘红原地区三种不同草甸土壤活性碳特征[J].山地学报,2008,26(2):205-211.
[12]张正斌,山仑.作物水分利用效率和蒸发蒸腾估算模型的研究进展[J].干旱地区农业研究,1997,15(1):73-78.
[13]许大全.光合作用效率[M].上海:上海科学技术出版社,2002.
[14]Wykoff W R.Measuring and modeling surface area of ponderosa pine needles[J].Canadian J Forest Research,2002,32(1):1-8.
[15]尹华军,赖挺,程新颖,等.增温对川西亚高山针叶林内不同光环境下红桦和岷江冷杉幼苗生长和生理的影响[J].2008,32(5):1072-1083.
[16]李畅,苏家乐,刘晓青,等.一品红不同品种叶片叶绿素荧光特性比较[J].园艺学报,2009,36(10):1519-1524.
[17]Platt T,Gallegos C L,Harrison W G.Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton[J].J Marine Research,1980,38:687-701.
[18]李英年,赵亮,徐世晓,等.祁连山海北高寒湿地植物群落结构及生态特征[J].冰川冻土,2006,28(1):76-84.
[19]周兴民.中国嵩草草甸[M].北京:科学出版社,2001.
[20]Zhang J W,Davies J.Increased synthesis of ABA in partially dehydrated root tips and ABA transport from roots to leaves[J].J Experimental Botany,1987,38:2015-2023.
[21]徐惠风,刘兴土,金研铭,等.沼泽植物泽泻气孔导度日变化的研究[J].生态科学,2003,22(3):218-221.
[22]栾金花.干旱胁迫下三江平原湿地毛苔草光合作用日变化特性研究[J].湿地科学,2008,6(2):221-228.
[23]李文瑞,冯金朝,江天然,等.沙冬青几种光合特性的季节性变化的研究[J].植物学报,1999,41(2):190-193.
[24]赵佐成.四川省红原县水生植物群落调查[J].武汉植物学研究,1996,14(3):213-222.
[25]师生波,韩发,卉桂英.高寒草甸矮嵩草群落光合作用的“午休”现象[J].植物生理学报,1997,23(4):405-409.
[26]王岩春,干友民,邱英,等.高寒地区“川草2号”老芒麦夏季光合生理生态特性的初步研究[J].草业与畜牧,2008(1):1-3.
[27]陈晓亚,汤章程.植物生理与分子生物学[M].北京:高等教育出版社,2007.
[28]陈模舜,柯世省,倪琼琼.田间珊瑚树净光合速率及生态因子的日变化[J].四川师范大学学报:自然科学版,2004,27(3):298-302.
[29]杜占池.羊草和大针茅光合作用午间降低与生态因子关系的研究[J].自然资源学报,1990,5(2):177-186.
[30]Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J].Ann Rev Plant Physiol,1982,33:317-345.
[31]薛崧,汪沛洪,许大全,等.水分胁迫对冬小麦CO2同化作用的影响[J].植物生理学报,1992,18(1):1-7.
[32]师生波,贲桂英,韩发.矮嵩草草甸植物群落的光合特性研究[J].植物生态学报,1996,20(3):225-234.
[33]Sparks J P,Black R A.Regulation of water loss in populations of populus trichocarpa:the role of stomatal control in preventing xylem cavitation[J].Tree Physiol,1999,19(7):453-459.
[34]赵佐成,魏泰昌,邱发英,等.若尔盖辖曼-阿西高原沼泽自然保护区沼泽植物特征[J].武汉植物学研究,1987,5(8):235-246.
[35]李宏林,徐当会,杜国祯.青藏高原高寒沼泽湿地在退化梯度上植物群落组成的改变对湿地水分状况的影响[J].植物生态学报,2012,36(5):403-410.
[36]Wang Y S,Chu C J,Maestre F T,et al.On the relevance of facilitation in alpine meadow communities:an experimental assessment with multiple species differing in their ecological optimum[J].Acta Oecologica,2008,33:108-113.
[37]许振柱,周广胜,李晖.羊草叶片气体交换参数对温度和土壤水分的响应[J].植物生态学报,2004,28(3):300-304.
[38]翁晓燕,蒋德安,陆庆.影响水稻光合日变化的酶和相关因素的分析[J].生物数学学报,1999,14(4):495-500.
[39]赵平,曾小平,彭少麟,等.海南红豆(Ormosia pinnata)夏季叶片气体交换、气孔导度和水分利用效率的日变化[J].热带亚热带植物学报,2000,8(1):35-42.
[40]吴建国.UV-B辐射增强对麻花艽叶片光合作用及相关生理参数的影响[J].环境科学研究,2010,23(11):1395-1404.
[41]Genty B,Briantais J M,Baker N R.The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence[J].Biochimica et Biophysica Acta:General Subjects,1989,990(1):87-92.
[42] Huner N P A,Oquist G,Hurry V M.Photosynthesis,photoinhibition and low temperature acclimation in cold tolerant plants[J].Photosynth Res,1993,37:19-39.
[43]Kolek J,Kozinka V.Physiology of the Plant Root System[M].Dordrecht:Kluwer Academic Publishers,1991.
[44]郭连旺,沈允钢,许大全,等.几种高寒草甸常见植物的光合特性及其光合作用的光抑制[C]//高寒草甸生态系统:第四集.北京:科学出版社,1995:65-74.
[45]Greer D H,Laing W A.Photoinhibition of photosynthesis in intact kiwifruit Actinidia deliciosa leaves,recovery and its dependence on temperature[J].Planta,1988,174:15-165.
[46]Franklin L A.The effects of temperature acclimation on the photoinhibitory responses of ulva rotundata blid[J].Planta,1994,192:324-331.
[47]张杰,杨传平,邹学忠,等.蒙古栎硝酸还原酶活性、叶绿素及可溶性蛋白含量与生长性状的关系[J].东北林业大学学报,2005,33(3):20-21.
[48]Ralph P J,Gademann R,Dennison W C.In situ seagrass photosynthesis measured using a submersible,pulse-amplitude modulated fluorometer[J].Marine Biology,1998,132:367-373.
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
科学(2022年4期)2022-10-25 02:43:00
青海草业(2022年2期)2022-07-23 09:34:58
干旱地区农业研究(2022年1期)2022-01-28 04:51:30
装备制造技术(2019年12期)2019-12-25 03:06:52
活力(2019年21期)2019-04-01 12:17:10
科技创新与品牌(2019年12期)2019-02-06 04:03:29
Coco薇(2016年5期)2016-06-03 09:17:41
考试周刊(2016年6期)2016-03-11 08:14:32
