
河北坝上地区坡面尺度根系生物量空间变异
2015-12-16 08:15王雪妍张卓栋罗梦婷廖家仪
水土保持研究 2015年6期
王雪妍,张卓栋,罗梦婷,覃 昕,廖家仪
(北京师范大学 地理学与遥感科学学院,北京100875)
根系是一株植物全部根的总称。根系在完成植物体本身结构功能之外,对水土保持、农业生产两大领域都具有重大意义。植物根系能有效保持坡面土壤稳固[1-2],还能够作为土壤酸碱度、肥力等特性的指示剂,以及牧草养护、利用的基本依据[3],对理论研究和实际应用都具有重要的意义。
河北坝上地区是温带山地落叶小叶林和温带草甸的交错地带[4],是京津地区与北部浑善达克沙地之间生态屏障[5],其生态环境的优劣,直接影响着京津的大气环境和水源安全。坝上草原地区根系的分布对坝上地区及其周边生态环境将起到重要作用。国内外学者已对不同地区植物根系空间分布有过大量研究,其研究对象包括新疆旱生芦苇根系与土壤水分的关系[6]、黄土区草地植被细根垂直分布与土壤环境的关系[7]、贺兰山油松根系空间分布特征[8]、意大利东北部Legnaro的甜菜根系[9]等。这些研究对于各地土壤中植物根系空间分异与土壤理化性质的探索具有推动作用,然而有关坝上草原的根系研究目前还少见报道。有限的坝上地区根系研究成果主要集中在根系的生物量与坡向因素的关系等[10],对坡面根系空间变异的研究还不够深入,无法充分掌握较大尺度的植物根系空间分布规律与空间变异程度。
相比于景观、流域[11]等水土保持研究尺度,坡面是水土流失研究的重要单元。本研究以坡面为研究尺度,通过根钻法实地采样,取得坝上草原研究区坡面上样点各个深度层次的根系生物量,通过空间插值等方法,构建坡面各层次根系生物量空间分布图,从而得到各土层根系生物量在坡面的分布特征。相比以往单点、单样带的研究,本研究中完整坡面的根系空间变异,更能反映坡面根系生物量分布的实际情况。同时,本研究在掌握坡面尺度上坝上地区根系的分布规律后,还可与小区、景观、流域等尺度研究结合,为全面多尺度掌握坝上植物根系分布情况提供有价值的参考,为坝上地区更为精细化的水土保持工作提供建议。
1 研究区和方法
1.1 研究区概况
丰宁坝上草原地处内蒙古高原东南缘,燕山山脉北部。海拔高度为1 500~1 800m。区域年平均气温约为0.6℃,年均降水量400mm 左右[4],雨热同期,属大陆性季风气候。由历史演变及气候环境,这里成为北方典型的温带农牧林交错区,植被以草本灌木为主。虽植被覆盖较好,根系繁多,然而单一的植被结构,短小的生物链使本地生态环境极为脆弱。数十年内,草原发生了严重的退化现象,土壤贫瘠,蒿类繁盛,风灾增多,侵蚀严重[5]。
本研究选取样地为河北省丰宁县大滩镇二道河村火石沟南侧一东坡,位置41.534°N,116.112°E,坡向88°,东西跨度280m,南北跨度200m,总面积约27 000 m2。坡面坡度14.8°,平缓延绵,属于典型的坝上草原丘陵山地地形[4]。坡面各样点优势种包括中华隐子草、散穗早熟禾、金狗尾草等典型温带草甸的植被类型[4]。坡面土地利用以放牧为主,坡面南北二百米外均为耕地,土地利用类型具较强的坝上地区代表性[5]。因此,该坡面具有典型坝上草原坡面特征。
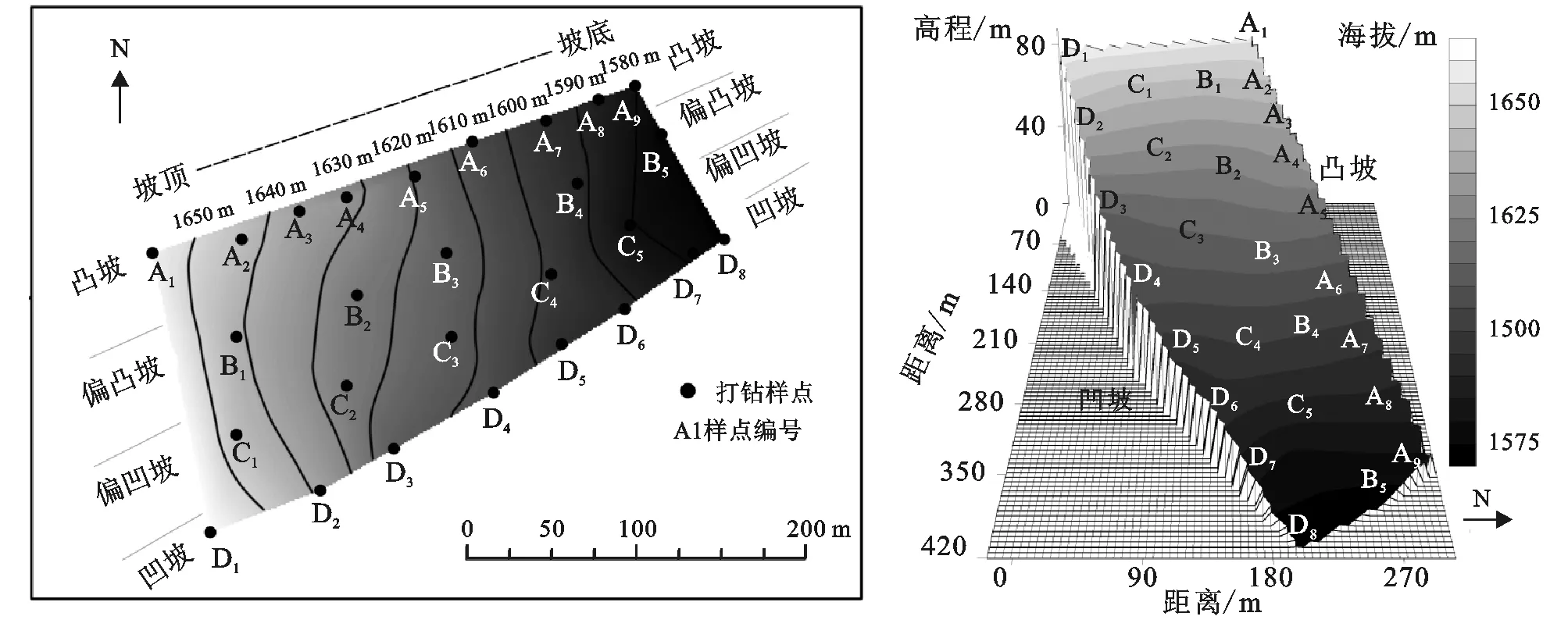
图1 样地数字高程模型及三维模型
1.2 研究方法
本研究以根系生物量为指标,使用Eijkelkamp根钻Edelman型钻头,采集直径8cm、长15cm土柱。在实地考察时,将研究地坡脊及坡谷,分别作为凸坡和凹坡样带,并以此为样地边缘。随后根据研究区实际情况,将本研究区分为四个部分,包括凸坡、偏凸坡、偏凹坡和凹坡。沿样带位置采集样点,其中凸坡9个样点,凹坡8个样点,两个过渡地带各5个样点。每样点分8个层次(0—30cm段分层间隔5cm,30—60cm段分层间隔15cm)。为便于下文叙述,将27个样点分别依照不同坡型样带分类编号,如图1所示。
在实验室对土壤根系样品进行清洗,分拣操作。将经过粗处理的根系置于60℃烘箱内烘8h至恒重,再使用研钵、镊子、有机玻璃棒等工具将根系与附着的土壤颗粒分离。最后使用精确至0.000 1g的电子天平对处理后的根系进行称重并记录。
利用SPSS软件对数据进行统计分析,完成可靠性检查后,进行根系生物量随深度变化的回归分析,得到指数回归方程。同时,将各样本根系生物量数据及点位空间信息导入ArcGIS 10.2软件,运用泰森多边形法改良的自然邻近插值法(Natural Neighbor)对根系数生物量进行空间插值,得到根系生物量在不同深度的根系密度的空间插值图,并进行交叉验证,对插值结果进行可靠性评价。该插值法具有使用便捷、参数简单,运算速度快等特点。
2 结果与分析
2.1 根系生物量数据统计特征
本次采样中,共得到206份土样。统计特征如表1所示。
由表1可知,凸坡平均根系生物量最高,达到4.68g/L。偏凸坡相差不大,数值为凸坡的91%,而凹坡及偏凹坡平均根重较少,分别只达到凸坡的79%和58%。在各个坡形中,偏凹坡土样之间根重差距较大,从最小值到最大值,增大了将近300倍,其变异系数也远高于其他坡形。

表1 不同坡形的样本统计特征

表2 不同深度样本根系生物量密度统计特征
由表2可知,从表层0-5cm段15.59g/L至底层的30—45cm段0.88g/L,根系生物量平均值减少为原先的二十分之一。各个深度层次土柱的变异系数相差不大,均在0.55左右,其中25—30cm深度的变异系数相对较大,20—25cm深度的变异系数最小。
2.2 根系生物量的空间分布特征
由自然邻近插值法(Natural Neighbor)法,得到不同深层根重插值图(图2)所示。根系生物量的极大值即各图中颜色比周围区域更深的点。插值得到的极大值结果无论属于样点上还是样点之间,大多都位于样地的边缘,即坡度变化较大的地形转变处。例如,凸坡边缘即为坡面与坡脊的转变处,出现根系生物量极大值样点7个(图2c中A2,A6两点、图2d中A6点、图2e中A1,A6两点、图2f中A7点,图2i中A7点,具体点编号位置信息见图1);凹坡即坡面与沟谷的转变处,出现根系生物量极大值的样点有10个(图2b中D4点、图2c中D1点、图2d中D6点、图2e中D5与D6之间一点、图2f中D1,D6两点、图2g中D4,D6两点、图2h中D4点、图2i中D6点)。
除位于南北两边缘的凸坡和凹坡的坡型极值外,东西两边缘坡底和坡顶的海拔极值处也因“地形转变”而出现根系生物量极大值。坡顶即坡面与山顶平面的地形转变处,出现根系生物量极大值的样点有5个(图2b中A1点、图2c中D1点、图2e中A1点、图2f中D1点、图2h中D1点);坡底即坡面与山底平面的地形转变处,出现根系生物量极大值的点约有2个(图2b中B5点、图2f中B5点)。在四条边缘线上,或由于坡面形状原因,坡顶和坡底出现的根系生物量极大值次数较少,凸坡和凹坡的根重极大值样点出现次数更大。在 ArcGIS 10.2中采用 Geostatistical Wizard功能对各深层插值结果进行Cross Validation(交叉验证)。在Prediction Error窗口中得到Mean(误差平均值)、Root-Mean-Square(误差均方根)、Mean Standardized(误差标准平均值)、Root-Mean-Square Standard(误差标准均方根)、Average Standard Error(平均标准误差),结果如表3所示。
各深层插值平均相对误差最小为10.2%,最大为35%,平均值20.8%。表层由于少量土样位置靠近植株,根系生物量异常大,导致平均标准误差加大。随层次加深,平均标准误差略有有减少趋势。总体插值结果较为理想。
2.3 根系生物量密度随深度分布
根系生物量密度随深度增加,迅速下降并渐渐平缓趋于零值,具有指数为负时的指数函数变化特征。将根系生物量密度(g/L)取自然对数形式,和剖面深度进行回归,方程形式如下:
lny=a-bx
其中纵截距a代表表层土壤根系生物量水平,斜率的负值b表达随深度增加时,根系生物量密度降低的速率[8]。绘制根系生物量密度随深度变化散点图及拟合线如下(图3)。

图2 不同层次根系生物量插值图
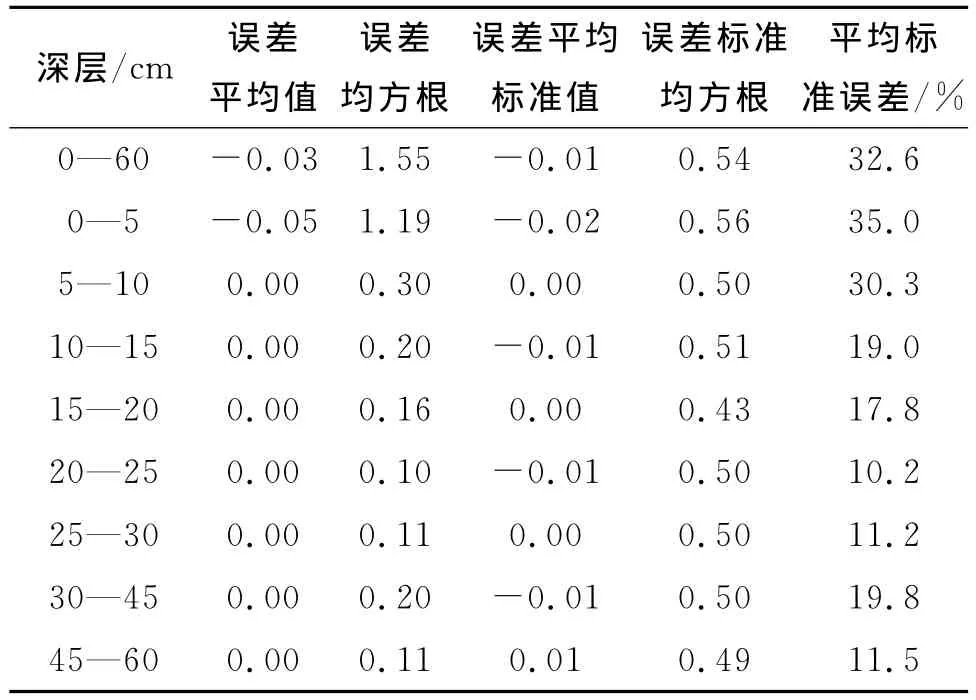
表3 插值模型交叉分析结果
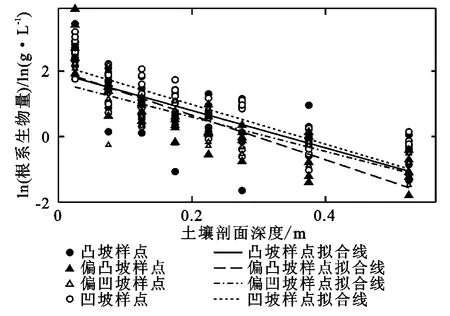
图3 各坡型根系生物量密度随深度变化散点图及回归曲线图
比较不同坡型散点图,其中凸坡、凹坡都存在若干根系生物量密度极大值,散点分布较为规律;偏凹坡数值较低且散点分布较为分散;偏凸坡样点中有根系生物量最大超过50g/L,为所有样品中最大值,散点分布非常集中。比较四条回归拟合曲线,凹坡纵截距最大,表层根系生物量水平最高;凸坡其次,与总平均情况相当。偏凸坡斜率最高,其根系生物量随深度增加而降低速率最快;偏凹坡斜率最低,其生物量降低速率最缓。将各坡型散点图分别进行回归(表4),均符合指数递减关系。在0.05置信水平下,结果均显著。进一步由不同坡形的回归方程可知,在本研究中,由凸坡到凹坡的四种坡形的b值分别为6.05,6.80,5.24,5.70,表明不同的地形单元的植物根系,其生物量随深度增加而降低的速率有所不同。

表4 各坡向根系生物量和剖面深度回归分析结果
3 讨 论
3.1 根系生物量空间插值规律
在坡面尺度上,地形的分布决定了水热再分配,从而决定了植物根系生物量在空间上的分布。干旱少雨是坝上地区生态环境恶化的主导因素[11],坡面水分条件从坡底到坡顶渐差,因此植物根系量渐低。孙中锋指出,在坡面不同立地条件的天然灌木林分的生物产量一般由坡下至坡上递减[12],本研究中由0—60cm根系生物量插值结果(图2a)可知,与之结论相同。减少趋势中的波动,或与牲畜放养和人为因素有关。坡底属于三道河一级河流阶地,又恰好有乡间土路穿过,牲畜觅食踩踏、人类的走动、农用车的行驶容易对下坡的植被产生影响。从单个层面而言,不同坡形的根系生物量情况有所不同,根重极大值出现在样地边缘。其中,凸坡根重是对土壤侵蚀不利条件的逆反馈。由于土壤侵蚀严重,细颗粒少,砾石较多且空隙大。冷蒿根系紧密攀附于砾石上,填补了砾石之间的空隙。而凹坡的根重情况,是充足水分条件的正反馈。凹坡本身可以汇聚雨水及山坡上冲刷下来的土壤颗粒,水土条件优越,植被覆盖度高,单位面积植株数量多,所以根重多出现极值。
3.2 根系生物量随深度变化规律
除水平分布外,根系生物量在垂直方向上的分布与土壤固土保水性能有密切联系。根系对土壤的加固是由于它们的拉伸强度和摩擦或粘附性质[13]。其中垂直于土壤表面延伸的根对土壤的加固,是通过增加根土体在剪切面上的的剪切强度实现的[14]。根系生物量随深度增加减少的速率,决定了其剪切强度。
本研究中根系生物量密度与剖面深度符合指数回归关系lny=a-bx。已有研究指出,指数回归式中b值越大,根系生物量随土层深度增加而减少的速率越大,剪切强度越差,保水性能越差[15]。王诗楠等在坝上草原同一地区以同样公式分析不同坡向的根系生物量随深度的变化,得到lny=2.540-0.039x的回归方程。本研究在此基础上,将根系生物量质量替换为根系生物量密度,解决了不同体积土样相互比较的问题,使方程更加严谨。在本研究中,由凸坡到凹坡的四种坡形的b值分别为6.05,6.80,5.24,5.70,说明凸坡及偏凸坡的根系生物量随深度增大而减少的速率大,深层次根系生物量较总量相比相对较少,对深层次水分吸收利用能力较弱,保水性能差;而凹坡及偏凹坡的由于b值相对较小,根系生物量随深度增大而减少的速率小,深层次根系生物量相对较多,能更多吸收利用深层次的水分,保水性能好,这与杨巧丽等人对于本公式b值与根系保水性能的阐释相符合。王巧利曾指出黄土高原沟谷地、近地表1 mm层根系生物量较大[16],本研究通过以上指数回归结果,发现坝上草原的根系生物量随深度变化也有类似现象。
但在这样的趋势下,保水性能并非沿坡型单调变化,而是存在波动。偏凸坡保水性能比“凸特征”更明显的凸坡保水性能差,偏凹坡的保水性能比“凹特征”更明显的凹坡更好。这样非单调的变化与样地地下钙积层及回归方法带来的误差有关。钙积层是土壤淋溶作用所致,其结构紧实,通透性差,影响根系向深层发展[17]。凸坡在中坡以上的部分样点,由于达到钙积层,根系生物量降为零无法参与指数回归,现实中其凸坡b值或许大于偏凸坡。其次本研究坡面地形复杂,一条弧形冲沟位于偏凹坡样带中下坡位置上,使偏凹坡某些位置汇水聚土能力较凹坡更佳,从而利于根系保水。所以本文认为,在保水性能自凸坡向凹坡逐步改善的趋势下,钙积层位置导致土壤保水性能降低,沟壑位置有助于土壤保水性能提高,这些客观因素也对根系生物量的分布起到作用。
3.3 对草原坡面植被保护的建议
在草原坡面上,加强植物根系是最经济高效的水土保持手段。其中水保效果好,利于根系生长的区域应该进行重点抚育保护,以解决时间紧、任务重、资金短缺、成活率不高等问题[18]。根系对土壤锚固效果在很大程度上取决于空间密度和深度[19],所以了解根系生物量在空间密度及各深度的分布,对于草原坡面植被分布具有很大意义。王诗楠等在坝上草原同样区域,采用类似方法研究了不同坡向上根系分布情况,得出南坡保水性能最佳[8]的结论。本文在此基础上,进一步研究了同一坡向不同坡形的根系空间分异。
在空间密度上,应利用坡面地形转折处对植被根系发展的优势,继续于凸坡脊、凹坡沟、坡顶、坡底等重点区域实施植被抚育。使之在适宜的条件下,形成不同深度贯通的根系网格,从而增加地表粗糙度,拦截坡面径流,削减流速,防止坡面侵蚀[20]。程龙飞曾提出,坡顶风化强烈,发生暴雨时其水流拖曳力及水流上升力会加剧水土流失[21]。这一结论与本研究对坡面网格的重点区域选择观点相同。同时在深度分布数据中,由于沟壑处根系生物量随土层深度增加而减少的速率最小,保水性能最佳,所以在沟谷充分利用水分条件,加强植被覆盖,其成功率较高。此外,坡底是坡面水土流失的最后一道屏障,相关部门应加强管理,减少牲畜觅食、人类通行踩踏、机动车碾压等等对坡底植被的破坏。综上所述,通过四周植被“框”、沟壑植被“带”以及坡底人为保护等措施,迅速建立起第一层植被防护,将有利于完成草原坡面进一步的更为全面的植被保护。
4 结 论
本研究利用根钻实地采样并分析,得到坝上草原坡面根系生物量在空间密度及深度上的分异规律。在空间密度方面,根系生物量的极值多分布于坡面区域的边缘,即地形转折处,其中凸坡和凹坡极大值出现最多,中下坡根系生物量相对较多。在深度方向上,根重密度随深度呈指数函数变化的特征,在凸坡和偏凸坡根系生物量随深度减少速率较大,保水性能较差,凹坡和偏凹坡根系生物量随深度减少速率较小,保水性能好的趋势下存在小范围波动,即含有钙积层的位置根系保水作用更差、沟壑处根系保水作用更佳。
当水土保持缩小到坡面研究尺度时,影响根系分布的因素主要为微地形因素。因此在恢复草原坡面植被时,应抓住地形特点,有重点地分阶段开展工作。首先应利用上述根系生长的有利地形,如地形转折处,在坡面边缘迅速覆盖第一批植被网格。同时利用沟壑保水有利条件,加强沟壑及周边植被覆盖,此后再逐步镶嵌填充,完成整个坡面的植被恢复与稳固。
致谢:北京师范大学地理学与遥感科学学院张科利教授对本研究进行了理论指导,高晓飞高级实验师对实验室烘干等操作提供了详细指导,董卫华副教授提供为本研究提供了Tremble GeoExplorer 6000 XT亚米级GPS仪器,本科生张东宇、董钊永在野外采样及样品处理过程中做出了贡献,在此一并致谢。
[1] 周云艳,陈建平,王晓梅.植物根系固土护坡机理的研究进展及展望[J].生态环境学报,2012,21(6):1171-1177.
[2] 刘定辉,李勇.植物根系提高土壤抗侵蚀性机理研究[J].水土保持学报,2003,17(3):34-37.
[3] 孙洪仁,武瑞鑫,李品红,等.紫花苜蓿根系入土深度[J].草地学报,2008,16(3):307-312.
[4] 中国地理图集编纂委员会.中国地理图集[M].中国地图出版社,2009.
[5] 李志祥,田明中,武法东,等.河北坝上地区生态环境评价[J].地理与地理信息科学,2005,21(2):91-93.
[6] 李修仓,胡顺军,李岳坦,等.干旱区旱生芦苇根系分布及土壤水分动态[J].草业学报,2008,17(2):97-101.
[7] 韦兰英,上官周平.黄土高原不同演替阶段草地植被细根垂直分布特征与土壤环境的关系[J].生态学报,2006,26(11):3740-3748.
[8] 成文浩,陈林.贺兰山油松林根系空间分布特征研究[J].水土保持研究,2013,20(1):89-93.
[9] Vamerali T,Guarise M,Ganis A,et al.Analysis of root images from auger sampling with a fast procedure:A case of application to sugar beet[J].Plant and Soil,2003,1(255):387-397.
[10] 王诗楠,张卓栋,韩玥,等.河北坝上地区不同坡向植物根系分布特征研究[J].水土保持研究,2014,21(2):152-157.
[11] 陈端吕,彭保发,李际平.植被水土保持效应尺度性研究综述[J].湖北农业科学,2012,51(3):433-437.
[12] 孙中峰,张学培.晋西黄土区坡面尺度土壤水分分布规律研究[J].水土保持通报,2014,34(2):25-30.
[13] Ekanayake J C,Marden M,Watson A J,et al.Tree roots and slope stability:a comparison between Pinus radiata and kanuka[J].New Zealand Journal of Forestry Science,1997,27(2):216-233.
[14] Zhou Y,Watts D,Li Y,et al.A case study of effect of lateral roots of Pinus yunnanensis on shallow soil reinforcement[J].Forest Ecology and Management,1998,103(2):107-120.
[15] 杨丽雯,张永清.4种旱作谷类作物根系发育规律的研究[J].中国农业科学,2011,44(11):2244-2251.
[16] 王巧利,贾燕锋,王宁,等.黄土丘陵沟壑区自然恢复坡面植物根系的分布特征[J].水土保持研究,2012,19(5):16-22.
[17] 闫培君,吴彤,王琰.乌兰察布市土壤钙积层对林木根系分布和林木生长影响情况研究[J].内蒙古科技与经济,2011(9):63-64.
[18] 王闰平,陈凯.中国退耕还林还草现状及问题分析[J].水土保持研究,2006,13(5):188-192.
[19] Reubens B,Poesen J,Danjon F,et al.The role of fine and coarse roots in shallow slope stability and soil erosion control with a focus on root system architecture:A review[J].Trees,2007,21(4):385-402.
[20] 余德亿,卢和顶.福建高速公路生物护坡草坪的建植与养护[J].草业科学,2001,18(5):45-49.
[21] 程龙飞,李林燕.库区坡面植被水土保持生态建设模式[J].水土保持研究,2010,17(5):251-253.
猜你喜欢
煤炭科学技术(2022年10期)2022-11-02
今日农业(2022年13期)2022-09-15
湖北植保(2022年4期)2022-08-23
科海故事博览·下旬刊(2022年4期)2022-05-07
食品安全导刊·中旬刊(2022年3期)2022-04-15
土壤(2021年1期)2021-03-23
西南石油大学学报(自然科学版)(2018年6期)2018-12-26
数学物理学报(2018年4期)2018-09-14
现代园艺(2017年23期)2018-01-18
土壤学报(2017年5期)2017-11-01
