
山西省油松林生态系统碳氮磷化学计量特征
2015-12-16 08:15张有利王百田王瑞君
水土保持研究 2015年1期
王 宁,张有利,王百田,王瑞君
(1.黑龙江八一农垦大学 农学院,黑龙江 大庆163319;2.北京林业大学 水土保持学院,北京100083)
生态系统碳(C)、氮(N)、磷(P)等元素的循环是全球变化研究的热点之一,而且C与N,P等元素的循环过程是相互耦合的[1-2],所以,养分循环的改变会强烈影响生态系统碳循环过程[3]。在相对稳定的条件下,生态系统C贮量是由关键养分元素N,P的供应量控制的[4],N,P缺乏意味着C的相对过量(C∶N和C∶P上升),反之,N,P充足则意味着C的相对不足(C∶N和C∶P下降)[5-6],因此,植物的 C,N,P化学计量(stoichiometry)比值在一定程度上可以指示所在生态系统的C积累动态和 N,P养分限制格局[7-9]。
叶片作为植物的主要光合器官,其N∶P的大小经常被用来表明生态系统生产力受到哪种元素的限制作用,但这种限制关系会随着外界环境的变化而变化[6,8,10-12],从全球尺度来看,高纬度地区的温带森林更易受N的限制,而低纬度地区的热带和亚热带森林更易受 P限制[11,13-14],即使同区域的植物也可能因为外界环境与内在属性的差别,其受限制的营养元素也不相同[11,13,15]。植被的生产力受到其所处环境条件中N,P可利用量的控制,土壤中N和P含量的大小在一定程度上可调节植物C∶N和C∶P的大小[5-6,16]。因此,植物体的C∶N和C∶P在一定程度上可反映单位养分供应量所能达到的生产力,N∶P也可用来表示植物受 N,P养分的限制格局[8,13,17-18]。目前全球或区域尺度上的植被C,N,P化学计量特征已有较多 研 究[8,14,19-22],但 将 植 物—凋 落 物—土 壤 作为一个完整的系统加以研究,探讨C,N,P元素化学计量比在整个系统中的变化格局及其相互作用的报道不多,对油松林生态系统的C,N,P化学计量学特征的研究更少。
油松(Pinus tabulaeformisCarr)是暖温带湿润半湿润气候区的地带性植被[23]。山西省的油松林在黄土区和太行山土石山区都有分布,其面积占全省森林总面积的26.19%[24]。目前,对于山西省油松林森林生态系统C,N,P化学计量学特征方面尚缺乏研究。本文以山西省天然油松林生态系统和人工油松林生态系统为研究对象,利用标准样地的实测数据,通过对植物叶片、凋落物和土壤的C,N,P含量和化学计量进行研究,探索分析油松林生态计量指标随林龄、林分起源的变化及可能的影响机制。
1 材料和方法
1.1 研究区概况
山西省位于黄土高原东部和太行山西部,介于东经110°14′—114°33′,北纬34°34′—40°43′,南北长约682km,东西宽约385km,总面积156 806km2,大部分地区海拔在1 000~2 000m。冬季长而寒冷干燥,夏季炎热多雨,北中部春季日温差大,风沙多,南部春季日温差较小,风沙小,北部秋季短暂,气候温和,南部秋季漫长,属暖温带半干旱大陆性区,年平均气温3~14℃,全年无霜期120~220d,多年平均降雨量400~650mm,森林覆盖率18.03%,森林土壤类型主要是棕壤、褐土、黄绵土。油松天然林主要分布在中条山、吕梁山、太岳山、太行山、关帝山、管涔山、五台山和黑茶山8大国有林区,人工林在山西各地均有分布,油松林面积占全省森林总面积的26.19%,且90%的油松林属于中幼龄林[23],林下灌木主要有毛榛(Corylus mandshuricaMaxim.)、黄刺玫(Rosa xanthinaLindl.)、绒毛绣线菊(Spiraea dasyantha)、胡枝子(Lespedeza bicolorTurcz.)、金银木(Lonicera maackiiRupr.Maxim.)、六道木(Abelia biflora)等,草本主要有披针苔草(Carex lancifolia)、唐松草(Thalictrum aquilegifoliumL.var.sibiricum Regel)、费菜(Sedum aizoonL.)、地 榆 (Radix Sanguisorbae)、穿 山 龙(Dioscorea nipponicaMakino)等。

图1 山西油松林样地分布
1.2 试验设计和样本采集
本研究按起源和龄组将油松林生态系统划分为人工幼龄林(artificial young forest,AY)、人工中龄林(artificial middle-aged forest,AM)、天然幼龄林(natural young forest,NY)和天然中龄林(natural middle-aged forest,NM)4种类型,并于2011年7月在山西省境内共选取38块油松林标准样地(50m×20m)进行调查取样(样地概况见图1和表1)。
试验主要由3部分组成:首先,在各样地选择油松标准木3株,利用高枝剪在树冠东、西、南、北4个部位采集叶片,为减少工作量,现场将每株标准木所采集叶片在托盘中进行混匀,利用四分法选取300g左右叶片,装入透气网兜,带回实验室备用,同时选择林下灌木和草本优势种及伴生种,其叶片采集和处理方法同上,每个样地获得乔木叶、灌木叶和草本叶各3个样本;其次,在样地内沿对角线设3个1m×1m样方,收集每个样方内全部凋落物,混匀后取样;最后,在样地3个样方内用土钻分5层(0—10cm,10—20cm,20—30cm,30—50cm,50—100cm)钻取土样(深至1m或C层为止)。共采集植物样品456个,土壤样品486个。
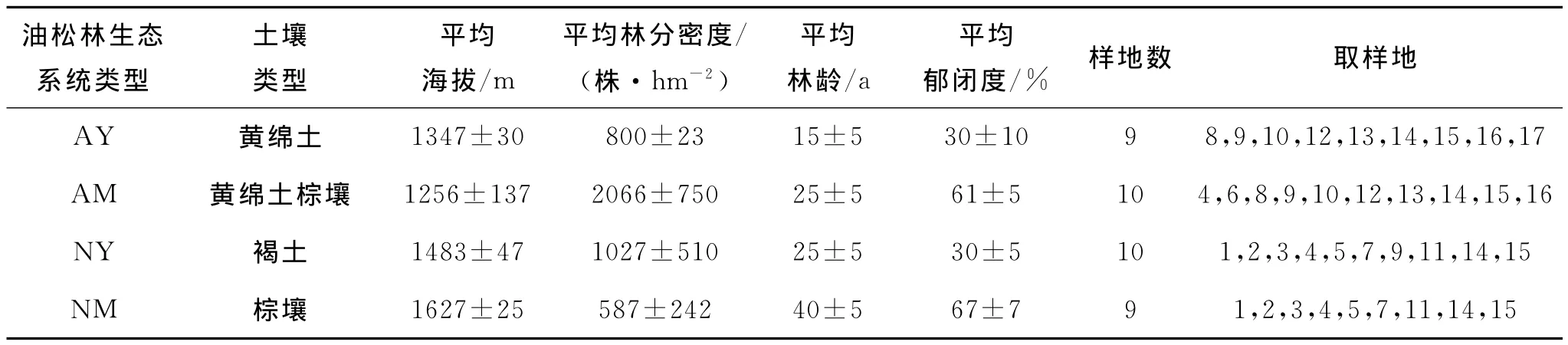
表1 油松林生态系统类型概况
1.3 室内测定、处理和化学分析方法
分析项目为植物叶片、凋落物和土壤的全碳、全氮、全磷含量。植物和凋落物样品在75℃烘箱中烘干至恒重,然后用高速万能粉碎机粉碎(FW100,天津)后装袋备测,土壤样品,经自然风干后碾碎并过0.25mm筛后装袋备测。所有样品的碳含量采用重铬酸钾法—浓硫酸消化硫酸亚铁滴定法,全氮含量采用凯氏定氮法(KDY-9820,北京),植物全磷含量采用钒钼黄比色法(UV-2550,日本)测定,土壤全磷含量采用钼锑抗比色法(UV-2550,日本)测定,每个样品重复测定3次。
1.4 数据分析
叶片、凋落物和土壤的C∶N∶P采用质量比表示。各生态系统叶片、凋落物和土壤的C,N,P含量和C∶N∶P为该生态系统内所有样地的均值。采用成对比较T检验判断油松林不同生态系统乔木叶、灌木叶、草本叶、凋落物、土壤间C,N,P含量与C∶N∶P的差异。利用单因素方差分析方法分别判断不同生态系统对叶片、凋落物和土壤的C,N,P含量和C∶N∶P的差别。采用Tukey检验进行各水平间均值的配对比较。
最后,采用Ⅱ类线性回归分析油松林生态系统植物叶片、凋落物和土壤C,N,P的化学计量关系,分析前,所有数值都经过以10为底的对数转换。在运算中,N,P含量与C含量间的关系,以及N,P含量间的关系用线性回归方程lgy=a+blgx来描述,其中,在分析前者关系时,x表示C含量,y表示N和P含量,分析后者关系时,x表示N含量,y表示P含量,方程中a表示截距,b表示斜率,当b=1时,表示两者存在等速比例关系,当b>1时,表示y的增加依赖于x的增加,当b<1时,表示y的增加不依赖于x的增加而增加。因单个生态系统样本数较少,所以本研究将4种油松林生态系统数据合并来说明油松生态系统叶片、凋落物和土壤C,N,P的化学计量关系。
以上所有数据处理应用Excel 2003和SPSS 18.0软件进行分析。
2 结果与分析
2.1 油松林生态系统叶片、凋落物和土壤碳氮磷含量
由表2可见,油松林生态系统的叶片、凋落物和土壤的C,N,P含量均随林龄增加而升高。4种油松林生态系统内乔木叶C含量均显著高于灌木叶和草本叶(p<0.05),乔木叶片N,P含量则显著低于灌木和草本叶片相应元素含量(p<0.05),乔木叶N,P含量最低,灌木叶N,P含量最高,叶片C,N,P含量都在植物生长的正常范围[24-25]。4种生态系统间乔木叶C,N,P含量差异不显著(p>0.05),AM 生态系统叶C,N,P含量最高,AY的叶C含量最低,NY的叶N,P含量最低,灌木叶C含量差异不显著(p>0.05),N含量在AY,AM,NM间差异不显著(p>0.05),P含量在AY,AM间差异显著(p<0.05),在NY,NM间差异不显著(p>0.05),草本叶C,N,P含量表现出林龄间差异显著(p<0.05),与林分起源无关。
AY,AM,NY,NM间凋落物C含量差异不显著(p>0.05),其变化范围为430.77~473.62g/kg,N含量在AY,AM间差异不显著(p>0.05),在NY,NM间差异显著(p<0.05),P含量在各系统间差异显著(p<0.05),天然林凋落物C,N含量高于人工林,而P含量低于人工林,不考虑林分起源条件下,凋落物C,N,P含量表现为中龄林>幼龄林,这与森林起源、林龄、林分郁闭度以及人为干扰强度有关[26]。

表2 油松林生态系统乔灌草叶片、凋落物和土壤的碳氮磷含量 g/kg
AY,AM,NY,NM 间土壤(0—100cm)C含量差异显著(p<0.05),表现为 NM(12.38g/kg)>NY(6.80g/kg)>AM(4.45g/kg)>AY(3.37g/kg),N含量在AY,AM间差异不显著(p>0.05),在NY,NM间差异显著(p<0.05),且天然林>人工林,P含量分别在AY,AM间和NY,NM间差异不显著(p>0.05),但在人工林和天然林间差异显著(p<0.05),天然林>人工林。土壤层各分层C,N,P含量分别见图2a—2c,C,N,P含量基本随土层深度增加而降低,C含量与土层深度的关系可用指数函数加以拟合(p<0.001),N,P含量与土层深度的关系均可用二项式函数加以拟合(p<0.001)。

图2 不同森林类型土壤碳氮磷含量剖面分布
2.2 油松林生态系统叶片、凋落物和土壤的C∶N∶P化学计量比
4种油松林生态系统叶片、凋落物和土壤的C∶N∶P化学计量比见表3。乔木、灌木叶片和凋落物的C∶N∶P均表现为天然林大于人工林,且差异明显(p<0.05),草本叶片的C∶N∶P表现为NM>AY>AM>NY,土壤层(0—100cm)的C∶N∶P则表现为AM>NM>NY>AY,土壤各分层C∶N∶P见表3。
乔木叶片的C∶N在4种生态系统间差异不显著(p>0.05),平均为35∶1,C∶P在人工林间差异不显著(p>0.05),平均为280∶1,在天然林间差异显著(p<0.05),平均为410∶1,N∶P在人工林间相同,为8∶1,天然林间差异不显著(p>0.05)。灌木叶片C∶N人工林间差异不显著(p>0.05),平均为17∶1,天然林间差异显著(p<0.05),C∶P天然林间差异不显著(p>0.05),平均为225∶1,人工林间差异显著(p<0.05),N∶P表现为 NM(15∶1)>AY(11∶1)>NY(9∶1)>AM(8∶1)。草本叶片C∶N在同龄林间差异不显著(p>0.05),幼龄林平均为26∶1,中龄林平均为22∶1,C∶P人工林间差异不显著(p>0.05),平均为206∶1,天然林间差异显著(p<0.05),N∶P人工林间差异不显著(p>0.05),平均为9∶1,天然林间差异显著(p<0.05)。
凋落物C∶N人工林间差异不显著(p>0.05),平均为47∶1,天然林间差异显著(p<0.05),C∶P生态系统间差异显著(p<0.05),表现为 NY(507∶1)>AY(347∶1)>NM(327∶1)>AM(246∶1),N∶P人工林间差异不显著(p>0.05),平均为9∶1,天然林间差异显著(p<0.05)。
土壤层(0—100cm)C∶N和N∶P各生态系统间差异不显著(p>0.05),平均为5∶1和6∶1,C∶P表现为 AM(37∶1)>NM(30∶1)>NY(28∶1)>AY(23∶1),生态系统土壤各分层C∶N表现基本从表层往下呈现降低的规律,但C∶P和N∶P表现规律不明显。
2.3 叶片、凋落物和土壤C,N,P的化学计量关系
在对叶片、凋落物和土壤C,N,P含量分别进行Ⅱ类线性回归分析后发现,乔木叶的N,P含量均与C含量间不存在显著线性关系(p>0.05)(见图3a),未反映出单位N,P含量的投入与C投入间的等速比例,乔木叶N含量和P含量间存在显著线性关系(p<0.05)(见图3b),斜率约等于1,表明乔木叶片建成过程中对N,P元素按比例投入的依赖,灌木叶N含量与C含量、N含量与P含量与间存在显著线性关系(p<0.05)(见图4a和图4b),斜率分别大于1和小于1,表明灌木叶C增加依赖于N的增加,N的增加不依赖于P的增加而增加,P含量和C含量间则不存在显著线性关系(p>0.05)(见图4a),草本叶C,N,P含量间均不存在显著线性关系(p>0.05)(见图5a和图5b),凋落物N含量和C含量间有显著线性关系(p<0.05)(见图6a),P含量和C,N含量间不存在显著线性关系(p>0.05)(见图6a和图6b),土壤(0—100cm)C,N,P含量间均存在显著线性关系(p<0.05)(见图7a和图7b),且 N,P含量的回归斜率约等于1,反映了土壤中单位P与N含量间的等速再吸收关系。

表3 油松林生态系统化学计量特征

图3 油松林生态系统乔木叶C,N,P的关系

图4 油松林生态系统灌木叶C,N,P的关系

图5 油松林生态系统草本叶C,N,P的关系

图6 油松林生态系统凋落物C,N,P的关系

图7 油松林生态系统土壤(0-100cm)C,N,P的关系
3 结论与讨论
人工林不同林龄乔木叶片间的C∶N∶P差异不显著(p>0.05),而天然林不同林龄的乔木叶片间差异显著(p<0.05),AY,AM,NY和 NM 乔木叶片的C,N和P的质量比率分别为276∶8∶1,283∶8∶1,458∶12∶1和362∶11∶1(折合成原子比分别为713∶18∶1,730∶18∶1,1 184∶28∶1和936∶23∶1),低于全球森林叶片的C∶N∶P原子比率(1 212∶28∶1)[27]和亚热带常绿针叶林叶片的 C∶N∶P原子比率(1 752∶31∶1)[14],4种类型乔木叶片的C∶N∶P平均原子比率(853∶21∶1)接近于温带针阔混交林叶片的C∶N∶P原子比率(829∶29∶1)[21]。AY,AM,NY 和 NM 灌木叶片的 C,N 和P的质量比率分别为181∶11∶1,132∶8∶1,228∶9∶1和221∶15∶1(折合成原子比分别为468∶23∶1,341∶18∶1,588∶19∶1和570∶32∶1),低于温带灌木叶片的C∶N∶P原子比率(752∶33∶1)[28]和亚热带常绿针叶林灌木叶片的C∶N∶P原子比率(1 656∶33∶1)[14]。AY,AM,NY 和 NM 草本叶片的C,N和P的质量比率分别为211∶8∶1,200∶9∶1,190∶8∶1和230∶11∶1,与中国大尺度草原区的研究结果相比[8,29],均显著低于草原区叶片的C∶N∶P平均水平(270∶15∶1),接近于温带草本叶片的C∶N∶P(202∶13∶1)[28]。本研究的叶片C∶N∶P与其他亚热带地区的研究有较大的分异,原因主要是本研究区域属半干旱地区,水分亏缺是限制该区域植物生长的重要因素[19,27],亚热带较温带地区温度高且水分充分,植物叶片有较高的C含量,在单位N,P养分条件下,其叶片建成效率显著高于本区域植物。
本研究4种油松林生态系统植物叶片的N∶P都小于14,前人研究表明叶片N∶P可用作氮饱和的诊断指标,并被用于确定养分限制的阈值[30],Tessier等[5]认为当植物叶片N∶P<14时存在氮限制,N∶P>16时则存在磷限制,即本研究植物叶片存在氮限制,结果支持研究得出的低纬度地区的植物更易受P限制,高纬度地区的植物更易受N限制的结论[11,31]。作为重要的生理指标,C∶N和C∶P可以反映植物生长速度[32],4种类型植物叶片C∶P均表现为乔木叶片>草本叶片>灌木叶片,表明油松林生态系统中乔木叶建成效率最高,而灌木叶建成效率最低。
AY,AM,NY和NM凋落物层的C,N和P的质量比率分别为347∶3∶1,246∶5∶1,507∶9∶1和327∶9∶1(折合成原子比分别为897∶16∶1,637∶12∶1,1 310∶20∶1和844∶20∶1),均显著低于全球森林凋落物的C∶N∶P原子比率(3 007∶45∶1)[27]、亚热带常绿针叶林凋落物的C∶N∶P原子比率(1 886∶31∶1)[14]和温带针阔混交林凋落物的C∶N∶P原子比率(1 426∶31∶1)[21],其与上述叶片表现出一致的规律,这与凋落物秉承植物活体的特性密切相关。
AY,AM,NY和NM土壤层(0—100cm)的C∶N∶P分别为59∶12∶1,95∶16∶1,71∶13∶1和77∶11∶1,而前人研究发现美国爱荷华州、巴西、苏格兰、新西兰和印度土壤的C∶N∶P(原子比)分别为78∶7∶1,161∶8∶1,87∶8∶1,59∶4∶1,67∶5∶1,76∶5∶1,[1]这种差异主要是由于土壤C∶N∶P主要受区域水热条件和成土作用特征的控制,由于气候、地貌、植被、母岩、年代、土壤动物等土壤形成因子和人类活动的影响,土壤碳氮磷总量变化很大,使得土壤C∶N∶P的空间变异性较大[33]。本研究4种生态系统类型土壤C∶N(4∶1~6∶1),低于中国湿润温带土壤中的C∶N(10∶1~12∶1)和热带、亚热带地区的红、黄壤的20∶1[34],主要是因为生态系统高密度部分有机质有着更低的C∶N[35],从土壤各分层来看,本研究4种生态系统土壤层随着土壤厚度的增加,C∶N降低,反映出土壤深层剖面腐殖质的年代和分层化[33]。
致谢:本文在外业调查取样过程中得到山西省林科院李新平院长,方山县林业局高海平工程师和马保明工程师,吉县张彦勤场长、文万荣书记,太岳林局王占勤总工,七里峪林场徐天红主任,灵空山保护区管理局王英杰局长,太行林局李康福局长和陈立强工程师,铁桥林场王景功工程师,以及北京林业大学生物学院曲红老师的大力帮助,在此一并谢过。感谢加拿大曼尼托巴大学University of Manitoba邓继峰博士对本文英文摘要上的润色。
[1] Stevenson F J,Cole M A.Cycles of soil carbon,nitrogen,phosphorus,sulfur,micronutrients[M].USA:John Wiley & Sons Inc,1999.
[2] Lal R.Soil carbon sequestration impacts on global climate change and food security[J].Science,2004,304(5677):1623-1627.
[3] Chapin F SⅢ,Matson P A,Vitousek P M.Principles of Terrestrial Ecosystem Ecology[M].Springer,New York.,Inc,2011.
[4] Hessen D O,A°gren G I,Anderson T R,et al.Carbon sequestration in ecosystems:the role of stoichiometry[J].Ecology,2004,85(5):1179-1192.
[5] Tessier J T,Raynal D J.Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation[J].Journal of Applied Ecology,2003,40(3):523-534.
[6] Güsewell S.N∶P ratios in terrestrial plants:variation and functional significance[J].New Phytologist,2004,164(2):243-266.
[7] 张丽霞,白永飞,韩兴国.N∶P化学计量学在生态学研究中的应用[J].植物学报:英文版,2004,45(9):1009-1018.
[8] He J S,Wang L,Flynn D F B,et al.Leaf nitrogen:phosphorus stoichiometry across Chinese grassland biomes[J].Oecologia,2008,155(2):301-310.
[9] 王绍强,于贵瑞.生态系统碳氮磷元素的生态化学计量学特征[J].生态学报,2008,28(8):3937-3947.
[10] Koerselman W,Meuleman A F M.The vegetation N∶P ratio:a new tool to detect the nature of nutrient limitation[J].Journal of Applied Ecology,1996:1441-1450.
[11] von Oheimb G,Power S A,Falk K,et al.N∶P ratio and the nature of nutrient limitation in Calluna-dominated heathlands[J].Ecosystems,2010,13(2):317-327.
[12] von Oheimb G,Power S A,Falk K,et al.N∶P ratio and the nature of nutrient limitation in Calluna-dominated heathlands[J].Ecosystems,2010,13(2):317-327.
[13] Wardle D A,Walker L R,Bardgett R D.Ecosystem properties and forest decline in contrasting long-term chronosequences[J].Science,2004,305(5683):509-513.
[14] 阎恩荣,王希华,郭明,等.浙江天童常绿阔叶林,常绿针叶林与落叶阔叶林的C∶N∶P化学计量特征[J].植物生态学报,2010,34(1):48-57.
[15] Davidson E A,de Carvalho C J R,Figueira A M,et al.Recuperation of nitrogen cycling in Amazonian forests following agricultural abandonment[J].Nature,2007,447(7147):995-998.
[16] Hogan E J,Minnullina G,Smith R I,et al.Effects of nitrogen enrichment on phosphatase activity and nitrogen:phosphorus relationships in Cladonia portentosa[J].New Phytologist,2010,186(4):911-925.
[17] Niva M,Svensson B M,Karlsson P S.Nutrient resorption from senescing leaves of the clonal plant Linnaea borealis in relation to reproductive state and resource availability[J].Functional Ecology,2003,17(4):438-444.
[18] A°gren G I.Stoichiometry and nutrition of plant growth in natural communities[J].Annual Review of Ecology,Evolution,and Systematics,2008,39:153-170.
[19] Han W,Fang J,Guo D,et al.Leaf nitrogen and phosphorus stoichiometry across 753terrestrial plant species in China[J].New Phytologist,2005,168(2):377-385.
[20] Richardson S J,Allen R B,Doherty J E.Shifts in leaf N∶P ratio during resorption reflect soil P in temperate rainforest[J].Functional Ecology,2008,22(4):738-745.
[21] 王晶苑,王绍强,李纫兰,等.中国四种森林类型主要优势植物的C∶N∶P化学计量学特征[J].植物生态学报,2011,35(6):587-595.
[22] 马露莎,陈亚南,张向茹,等.黄土高原刺槐叶片生态化学计量学特征[J].水土保持研究,2014,21(3):57-61.
[23] 吴刚,冯宗炜.中国油松林群落特征及生物量的研究[J].生态学报,1994,14(4):415-422.
[24] 冯建成.山西省森林植被的碳贮量研究[J].山西林业科技,2010,39(1):16-18.
[25] Marschner H.Mineral nutrition of higher plants[M].New York:Academic Press,1995.
[26] NiinemetsÜ,Kull O.Biomass investment in leaf lamina versus lamina support in relation to growth irradiance and leaf size in temperate deciduous trees[J].Treephysiology,1999,19(6):349-358..
[27] 方运霆,莫江明,彭少麟,等.森林演替在南亚热带森林生态系统碳吸存中的作用[J].生态学报,2003,23(9):1685-1694.
[28] McGroddy M E,Daufresne T,Hedin L O.Scaling of C∶N∶P stoichiometry in forests worldwide:implica-tions of terrestrial Redfield-type ratios[J].Ecology,2004,85(9):2390-2401.
[29] 韩文轩,吴漪,汤璐瑛,等.北京及周边地区植物叶的碳氮磷元素计量特征[J].北京大学学报:自然科学版,2009,45(5):855-860.
[30] He J S,Fang J,Wang Z,et al.Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China[J].Oecologia,2006,149(1):115-122.
[31] Güsewell S,Koerselman W,Verhoeven J T A.Biomass N∶P ratios as indicators of nutrient limitation for plant populations in wetlands[J].Ecological Applications,2003,13(2):372-384.
[32] Chapin III F S,Moilanen L.Nutritional controls over nitrogen and phosphorus resorption from Alaskan birch leaves[J].Ecology,1991,72(2):709-715.
[33] A°gren G I.The C∶N∶P stoichiometry of autotrophs:theory and observations[J].Ecology Letters,2004,7(3):185-191.
[34] Batjes N H.Total carbon and nitrogen in the soils of the world[J].European Journal of Soil Science,1996,47(2):151-163.
[35] 黄昌勇.土壤学[M].北京:中国农业出版社,2000.
[36] De Camargo P B,Trumbore S E,Martinelli L Z,et al.Soil carbon dynamics in regrowing forest of eastern Amazonia[J].Global Change Biology,1999,5(6):693-702.
猜你喜欢
山西林业(2021年2期)2021-07-21
农村百事通(2020年21期)2020-11-23
绿色中国(2019年19期)2019-11-26
绿色中国(2019年19期)2019-11-26
世界家苑(2018年9期)2018-09-18
乡村科技(2018年8期)2018-02-13
现代农村科技(2018年3期)2018-01-18
诗潮(2017年2期)2017-03-16
湖南林业科技(2017年1期)2017-02-06
