
苍白球外侧部生理功能及其对运动调控作用研究进展
2015-12-16 01:13:39刘军
河北体育学院学报 2015年6期
刘 军
(西安体育学院健康科学系,西安 710068)
苍白球外侧部生理功能及其对运动调控作用研究进展
刘 军
(西安体育学院健康科学系,西安 710068)
众多研究提示,基底神经节主要通过直接通路、间接通路和超直接通路进行运动调控,运动疲劳是由于基底神经节与皮层之间三条通路的失衡导致皮层不能发放有效的神经冲动引起的。苍白球外侧部在基底神经节的功能核团中居于中心地位,就国内外关于苍白球外侧部的生理功能及运动疲劳时苍白球外侧部在运动调控方面的研究现状予以梳理,旨在为相关研究提供参考。
基底神经节;苍白球外侧部;运动调控;疲劳
苍白球(globus pallidus,GP)位于豆状核内侧部,是基底神经节(basal ganglia,BG)的重要组成核团之一,上面因为分布的血管稀少,在新鲜标本上呈苍白色而得名。苍白球可进一步通过内侧髓板分为外侧部(external segment of the Globus pallidus,GPe)和内侧部(Internal segment of the globus pallidus,GPi),两部分虽然位置邻近但功能有很大区别。啮齿类动物没有苍白球内侧部,其苍白球相当于灵长类动物的苍白球外侧部。为方便叙述,本文所指的苍白球外侧部在涉及啮齿类动物时只指苍白球。
1 基底神经节在动作控制中的作用
大脑皮层通过锥体系和锥体外系来调控躯体运动。基底神经节是锥体外系调控通路的重要组成部分,将来自皮层的信号加工处理后反馈给皮层,与运动控制、运动过程、感觉性和运动性冲动的整合作用、奖赏、动机和情感等功能密切相关[1-2],在运动调控中起着非常重要的作用。借助于本体感受反馈冲动,基底神经节还可以对皮质发出的运动方案进行校正,以实现更为精确的运动控制。早有研究表明,基底神经节功能紊乱导致的运动功能失调与帕金森疾病、精神分裂症、亨廷顿舞蹈病和药物成瘾等密切相关[3]。从功能角度可将基底神经节划分为纹状体(Str,尾核和壳核)、苍白球外侧部、苍白球内侧部—黑质网状部复合体(GPi-SNr)、黑质致密部(SNc)及丘脑底核(STN)等五个功能核团,通过与皮层组成复杂调节环路联络从而兴奋或抑制性影响运动系统[4-5]。
运动时,来自外周神经系统的感觉输入和来自中枢神经系统的动机输入同时到达中枢神经系统,前者激活抑制系统限制运动输出,而后者通过基底神经节激活易化系统增加运动输出[6](图1)。因此,基底神经节在运动的易化通路中占据中心地位。
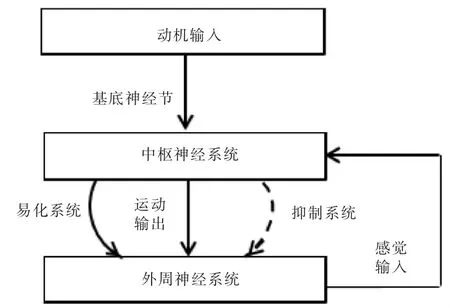
图1 动作输出通过易化和抑制系统调控模式图
2 苍白球外侧部在基底神经节调控通路中的中心作用
基底神经节通过直接通路(皮层—纹状体—苍白球内侧部/黑质网状部—丘脑腹外侧核/丘脑腹前核—皮层)、间接通路(皮层—纹状体—苍白球外侧部—丘脑底核—苍白球内侧部/黑质网状部—丘脑腹外侧核/丘脑腹前核—皮层)和超直接通路(皮层—丘脑底核—苍白球内侧部/黑质网状部—丘脑腹外侧核/丘脑腹前核—皮层)来整合和处理来自皮层的信息[7-8](图2)。研究发现,纹状体投射向直接通路的是表达有多巴胺(dopamine,DA)D1受体和P物质的中等棘状神经元(medium spiny neurons,MSNs),而投射向间接通路的是表达有DAD2受体和脑啡肽的MSNs。一般认为DA对两条纹状体苍白球通路有不同的效果:通过D1受体易化直接通路,通过D2受体抑制间接通路,这样一来,原来起兴奋作用的直接通路和起抑制作用的间接通路经过来自黑质的DA调节后均可以易化运动。
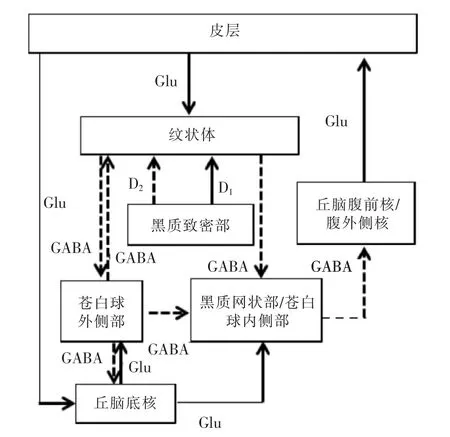
图2 皮质—基底神经节环路示意图
由上述的调控模式可知,在基底神经节中,苍白球外侧部绝不仅仅是传统观点认为的属于间接调控通路的一个中继核团,它同时接受基底神经节两大信息输入核团纹状体和丘脑底核的投射性纤维,并且发出纤维返回投射到纹状体,这些纤维投射与丘脑底核—苍白球外侧部之间的往返纤维投射平行[9]。此外,苍白球外侧部发出大量的纤维投射到苍白球内侧部。它不仅与基底神经节的输入结构纹状体和丘脑底核都相互联系,还是输出核团苍白球内侧部和黑质网状部的主要纤维支配来源,但并没有纤维投射向基底神经节以外的结构,只形成由丘脑底核和纹状体到苍白球外侧部再到苍白球内侧部和黑质网状部的单向信息流,因此,它可能是基底神经节各功能核团的中心(图3)[10]。
基底神经节另外一个重要特征是核团的体积和神经元的数量随着纹状体—苍白球外侧部—苍白球内侧部信息轴的传递而剧烈减少。人类背侧纹状体(包括尾壳核)的体积为9 700 mm2,这分别是苍白球外侧部(800 mm3)的12倍、苍白球内侧部(480 mm3)和黑质网状部(410 mm3)的20—25倍,而丘脑底核(160 mm3)的体积更是小到容易被忽略[10]。不过考虑到相比纹状体更小的体积和更大的神经元密度,这种体积的递减就变得合乎情理。由皮层到纹状体的投射几乎比纹状体本身神经元的数量至少多一个数量级[11]。灵长类动物中纹状体MSNs的数量约为10 000 000个,苍白球外侧部和苍白球内侧部神经元数量各为150 000 和50 000个[12];而在啮齿类动物中这样的漏斗状结构依然存在,MSNs数量为1 700 000—2 800 000,苍白球外侧部和苍白球内侧部神经元数量各为46 000和3 200个[13]。
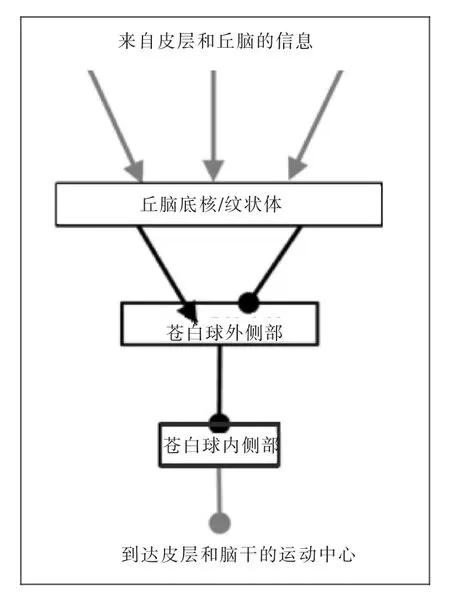
图3 苍白球外侧部在基底神经节信息流中的中心地位
3 苍白球外侧部纤维联系与神经元类型
3.1 苍白球外侧部的纤维投射
苍白球外侧部的树突都被来自于其他细胞的树突所形成的突触覆盖,在哺乳动物中,一个苍白球外侧部神经元上面树突所联系的平均突触数目估计有30 000—40 000个,这些突触的绝大多数(80%—90%)都是纹状体—苍白球外侧部GABA(γ-氨基丁酸)能神经末梢,除此之外,苍白球外侧部还接受约占总数10%的来自丘脑底核的兴奋性谷氨酸(Glu)能的投射纤维[14]。纹状体和苍白球外侧部之间的神经联系是交互型的,即相同的纹状体和苍白球外侧部神经元会分别接受很多苍白球外侧部和纹状体神经元的投射,形成信息交联[15]。苍白球外侧部神经元内部也有GABA能的局部轴突的侧支投射,形成回返性抑制。另外,苍白球外侧部神经元表面分布有DA和腺苷的受体亚型,说明除了GABA 和Glu能投射之外,来自黑质的DA能神经纤维也对苍白球外侧部神经元的生理功能具有调控作用。
苍白球外侧部神经元发出的纤维与基底神经节所有其他核团形成联系,除正常投射到其下游核团丘脑底核、苍白球内侧部/黑质网状部复合体之外,还和纹状体神经元形成大量突触联系,从而使苍白球外侧部的功能不再局限于间接通路的中继作用,成为对基底神经节信息接受核团纹状体和丘脑都会形成直接影响的中心核团。
3.2 苍白球外侧部神经元类型和放电特征
苍白球外侧部是通过基底神经节调整神经活性的整合中心,苍白球外侧部的不同区域有不同的功能,其中的每一种理论上都有相似的细胞类型。因此,理论上脊椎动物苍白球外侧部的神经元是一种快速放电的GABA能神经元,通过支配丘脑底核而行使特定功能[16]。根据放电的时间特征,苍白球外侧部神经元被划分为2个亚群,即GP-TI和GP-TA,前者是典型的GABA能神经元(75%)支配基底神经节下游的核团,神经元上面表达有GDA67和小清蛋白,然而后者(20%)却将大量的GABA/脑啡肽能神经元投射向纹状体。因此,和纹状体两类投射神经元一样,这两类神经元也在基底神经节环路中扮演各自的角色,功能互补。其余5%的神经元是胆碱能神经元,目前功能尚不清楚。与此相应,有85%左右的苍白球外侧部神经元在相对高频率的范围内(50—70 Hz)自发放电,并且会被可长达0.3—2 s的间隔打断,另外一类神经元在较低频率(<15 Hz)放电,伴随间歇性、较短的高频爆发式放电[17]。
4 苍白球外侧部与运动失调性疾病
运动失调性疾病主要是因为某种病理原因导致运动过多或过少,前者典型如亨廷顿病(Huntington disease,HD),后者如帕金森疾病(Parkinson disease,PD)、肌张力障碍(Dystonia)和运动障碍(Dyskinesia)等。这类疾病发病大多是因为基底神经节相关调控核团的功能异常。
4.1 苍白球外侧部和PD
PD是由英国医生James Parkinson于1817年首先描述的以中枢神经系统退行性病变为特征引发的运动障碍疾病,主要病理表现为黑质DA能神经元变性脱失,临床表现为静止性震颤、运动徐缓、肌张力增高和步态异常等。目前研究认为PD主要是因为黑质DA神经元变性死亡导致的黑质对纹状体的DA能投射调控异常,特别是在受D2受体负调控的间接通路中,因为DA对间接通路的去抑制作用减弱或丧失,造成基底神经节通过丘脑对皮层的驱动不足而导致运动不能症状。电生理学研究也证实,在PD发生时,苍白球外侧部神经元活性下降,而丘脑底核神经元的活性增加。另有研究发现,MPTP处理的PD模型猴子苍白球外侧部放电频率减少,但也有放电频率没有变化甚至相反的结果出现,证明基底神经节内各核团放电节律改变在PD发病中起了非常重要的作用,但肯定还有其他原因同时存在[18]。
此后人们对神经元放电形式即爆发式放电和常规放电等指标加以研究,在PD病人和PD模型动物中可观察到苍白球外侧部爆发式放电形式增加,这一度被作为PD中基底神经节异常放电的可靠判断指标,与PD时的僵直、震颤以及运动不能有密切关系,反之,如果对相应核团神经元进行电刺激而减少这种异常的爆发放电形式,则有利于缓解帕金森的症状[19]。放电形式还可以以重复性的振荡方式出现,在对PD患者和PD动物模型研究时都发现,在苍白球外侧部神经元放电同步化增加,并出现显著的β频率波段(大约10—35 Hz)的振荡活动,而且在丘脑底核也倾向于同时出现这种同步化放电[20],且能被左旋多巴抑制[21]。振荡产生的机理仍不清楚,推测和DA能神经元的丢失有关[22]。
4.2 苍白球外侧部和HD
HD是一种常染色体显性遗传疾病,主要病理改变是纹状体神经元的丢失,症状主要表现为异常的舞蹈样不自主动作、情绪改变和认知功能方面的障碍。由于纹状体神经元的变性死亡,控制GABA和乙酰胆碱合成的谷氨酸脱羧酶和胆碱乙酰化酶活性降低,由纹状体GABA能投射神经元和胆碱能中间神经元分别分泌的神经递质GABA和乙酰胆碱合成减少,而由黑质投射神经元产生的DA合成却相对增多,因此,导致舞蹈样的多余动作产生和认知功能下降。
在HD发生过程中,由纹状体投射向苍白球外侧部的间接通路上的MSNs出现病变被认为是早期主要病因之一[23]。这部分神经元变性丢失后,投射向苍白球外侧部神经元的GABA能抑制减少,导致苍白球外侧部神经元兴奋性增高而丘脑底核兴奋性降低,从而使间接通路对丘脑的抑制性输出不足,两条通路对皮层的反馈性调节失去平衡,因此产生运动过多的症状[24]。而且在HD病例中确有发现苍白球外侧部放电节律的上升现象,直接证实了上述理论推测的正确性[25]。
5 苍白球外侧部与运动疲劳的可能关系
关于疲劳和基底神经节的研究,主要集中在临床上如多发性硬化和小儿麻痹等疾病所导致的疲劳的中枢机制,也正是在对诸如PD、HD和肌张力障碍等疾病的研究中发现苍白球外侧部可能在动作输出方面具有重要作用[26]。一项对慢性疲劳综合症(chronic fatigue syndrome,CFS)开展的较新研究认为,CFS的疲劳产生可能由基底神经节输出的活性减少所致,并推测可能与苍白球到丘脑和皮层的投射紊乱有关[27]。
现在比较一致的认识是中枢神经系统发放的保护性抑制才是运动疲劳产生的根本原因,运动疲劳产生时,神经系统不能有效募集运动神经元,中枢不能发放或维持足够的神经冲动[28]。进一步研究提示,疲劳发生时,组成间接通路的纹状体和丘脑底核神经元兴奋性增高,基底神经节的输出核团苍白球内侧部和黑质网状部神经元兴奋性增强,随着运动疲劳的发生,皮层运动区神经元电活动被广泛抑制,均提示由纹状体发出的基底神经节皮层环路调控紊乱可能是导致运动疲劳发生的中枢机制[7]。而如前文所述,苍白球外侧部是联系基底神经节信息接受核团和信息输出核团的重要中介,对直接通路、超直接通路和间接通路均有调控作用,在基底神经节信息调控中居于中心地位[10]。
有研究通过在体玻璃微电极技术记录一次力竭运动后大鼠苍白球外侧部神经元自发放电活动的变化时发现,运动疲劳后大鼠苍白球外侧部低频爆发式放电神经元放电频率显著增加、比例显著增高,而高频间隔放电神经元比例显著降低,提示苍白球低频爆发放电神经元在运动疲劳中枢调控中发挥着重要作用,运动疲劳状态下,苍白球外侧部与基底神经节其他核团网络连接更加紧密,神经元放电趋于同步[29]。而在另外一项研究中,发现大鼠苍白球GABA、Glu分别通过GABAARα1和GluR2共同参与了一次力竭运动过程中大鼠苍白球神经元兴奋性的调节,并据此认为,力竭时大鼠胞外GABA的增多和相应GABAARα1表达上调而对间接通路激活过度,导致对皮层的抑制性冲动增多,是运动疲劳产生的原因之一[30]。进一步的研究证实,向苍白球外侧部直接注入GABA激动剂会严重妨碍运动功能,而注入拮抗剂则会有益于运动效果[31]。
6 结语
苍白球外侧部在基底神经节—皮层环路的调控中居于中心地位,苍白球外侧部通过调控基底神经节的信息接受核团和信息输出核团,影响基底神经节对丘脑的兴奋程度最终易化或抑制运动皮层。据此,已有少量研究开始关注苍白球外侧部在运动疲劳发生过程中的作用,但因为苍白球外侧部本身细胞构筑和纤维联系复杂,所以在运动疲劳发生过程中调控苍白球外侧部相应的递质、受体和信息传导过程中的关键信号分子都值得进一步关注。
[1]Lanciego J L,Luquin N,Obeso J A.Functional Neuroanatomy of the Basal Ganglia[J].Cold Spring Harb PerspectMed,2012,2(12):a009621.
[2]Graybiel A M.The basal ganglia:Learning new tricks and loving it[J].Curr Opin Neurobiol,2005,15(6):638-644.
[3]Graybiel A M.Neurotransmitters and neuromodulators in the basal ganglia[J].Trends Neurosci,1990,13(7):244-254.
[4]Nambu A,Tokuno H,Takada M.Functional significance of the cortico-subthalamo-pallidal‘hyperdirect’pathway [J].Neurosci Res,2002,43(2):111-117.
[5]Wall N R,De La Parra M,Callaway EM,et al.Differential innervation of direct-and indirect-pathway striatal projection neurons[J].Neuron,2013,79(2):347-360.
[6]Tanaka M,Watanabe Y.Supraspinal regulation of physical fatigue[J].NeurosciBiobehav Rev,2012,36(1):727-734.
[7]乔德才,刘军,刘晓莉.运动疲劳的中枢机制研究进展:基于基底神经节—皮层环路紊乱的视角[J].北京体育大学学报,2014,37(2):51-58,65.
[8]Nambu A.A new dynamicmodel of the cortico-basal ganglia loop[J].Prog Brain Res,2004(143):461-466.
[9]Bolam JP,Hanley JJ,Booth PA,et al.Synaptic organisation of the basal ganglia[J].Anat,2000,196(Pt4):527-542.
[10]Goldberg JA,Bergman H.Computational Physiology Of The Neural Networks Of The Primate Globus Pallidus:Function And Dysfunction[J].Neuroscience,2011(198):171-192.
[11]Sato F,Lavallée P,Lévesque M,et al.Single-axon tracing study of neurons of the external segment of the globus pallidus in primate[J].JComp Neurol,2000,417(1):17-31.
[12]Redgrave P,Coizet V,Comoli E,et al.Interactions between themidbrain superior colliculus and the basal ganglia[J].Front Neuroanat,2010(4):132.
[13]Haber SN,Knutson B.The reward circuit:linking primate anatomy and human imaging[J].Neuropsychopharmacology,2010,35(1):4-26.
[14]Chan C S,Surmeier D J,YungW H.Striatal information signaling and integration in globus pallidus:timing matters[J].Neurosignals,2005,14(6):281-289.
[15]Kita H.Globus pallidus external segment[J].Prog Brain Res,2007(160):111-133.
[16]Stephenson-Jones M,Samuelsson E,Ericsson J,et al.Evolutionary conservation of the basal ganglia as a common vertebratemechanism for action selection[J].Curr Biol,2011,21(13):1081-1091.
[17]Mallet N,Pogosyan A,Márton L F,et al.Parkinsonian beta oscillations in the external globus pallidus and their relationship with subthalamic nucleus activity[J].JNeurosci,2008,28(52):14245-14258.
[18]Soares J,Kliem M A,Betarbet R,et al.Role of external pallidal segment in primate parkinsonism:comparison of the effects of1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced parkinsonism and lesions of the external pallidal segment[J].J Neurosci,2004,24(29):6417-6426.
[19]Wichmann T,Soares J.Neuronal firing before and after burst discharges in themonkey basal ganglia is predictably patterned in the normal state and altered in parkinsonism[J].JNeurophysiol,2006,95(4):2120-2133.
[20]Hammond C,Bergman H,Brown P.Pathological synchronization in Parkinson’s disease:networks,models and treatments[J].Trends Neurosci,2007,30(7):357-364.
[21]Weinberger M,Mahant N,Hutchison W D,et al.Beta oscillatory activity in the subthalamic nucleus and its relation to dopaminergic response in Parkinson’s disease[J].JNeurophysiol,2006,96(6):3248-3256.
[22]Weinberger M,Dostrovsky JO.A basis for the pathological oscillations in basal ganglia:the crucial role of dopamine[J].Neuroreport,2011,22(4):151-156.
[23]Menalled L,ZanjaniH,MacKenzie L,etal.Decrease in striatal enkephalin mRNA in mouse models of Huntington’s disease[J].Exp Neurol,2000,162(2):328-342.
[24]Starr P A,Kang G A,Heath S,et al.Pallidal neuronal discharge in Huntington’s disease:support for selective loss of striatal cells originating the indirect pathway[J].Exp Neurol,2008,211(1):227-233.
[25]Fasano A,Mazzone P,Piano C,etal.GPi-DBS in Huntington’s disease:results onmotor function and cognition in a 72-year-old case[J].Mov Disord,2008,23(9):1289-1292.
[26]胡荣光,刘晓莉,侯莉娟,等.苍白球内侧部、黑质网状部与运动调控[J]河北体育学院学报,2013,27(3):69-73.
[27]Miller A H,Jones JF,Drake D F,etal.Decreased basal ganglia activation in subjects with chronic fatigue syndrome:association with symptoms of fatigue[J].PLoS One,2014,9(5):e98156.
[28]Secher N H,Quistorff B,Dalsgaard M K.The muscles work,but the brain gets tired[J].Ugeskr Laeger,2006,168(51):4503-4506.
[29]侯莉娟,林然,刘晓莉,等.一次性力竭运动对大鼠苍白球外侧部神经元自发放电活动的影响[J].西安体育学院学报,2015,32(2):196-200.
[30]乔德才,张吉敏,侯莉娟,等.大鼠苍白球GABA、Glu在力竭运动过程中的调控作用研究[J].沈阳体育学院学报,2015,34(2):82-86.
[31]Obeso JA,Rodriguez-Oroz M C,Javier Blesa F,et al.The globus pallidus pars externa and Parkinson's diseaseReady for prime time[J].Exp Neurol,2006,202(1):1-7.
Physiological Function and M odulation to Physical M ovement of External Segment of Globus Pallidus
LIU Jun
(Department of Health Sciences,Xi’an Physical Education University,Xi’an 710068,China)
Many studies show that basal ganglia regulates sportmovementsmainly by direct access,indirect access and ultra-direct access,sport fatigue happens when the imbalance between the three accesses leads to failure of the cortex to trigger nerve impulses.The external segment of globus pallidus is in central position of the functional groups of the basal ganglia.This article combs the previous researches at home and abroad about the physiological function of external segment of globus pallidus and itsmodulation to physicalmovement in sport fatigue,aiming to offer references for relevant studies.
basal ganglia;external segment of globus pallidus;modulation to physicalmovement;fatigue
G804.2
A
1008-3596(2015)06-0078-05
2015-08-11
西安体育学院院管课题(2012YB001);西安体育学院博士启动基金
刘 军(1980-),男,甘肃民勤人,副教授,博士,研究方向为运动疲劳的机制与调控。
猜你喜欢
影像科学与光化学(2022年5期)2022-09-26 06:17:34
中国临床医学影像杂志(2021年10期)2021-11-22 07:46:30
海南师范大学学报(自然科学版)(2020年1期)2020-04-18 06:18:46
山东医学高等专科学校学报(2018年4期)2018-03-18 05:51:58
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
动物营养学报(2016年2期)2016-03-10 22:12:05
中国组织化学与细胞化学杂志(2016年4期)2016-02-27 11:15:55
磁共振成像(2015年7期)2015-12-23 08:53:04
环球中医药(2015年4期)2015-02-27 15:01:51
医学信息(2014年13期)2014-07-09 19:50:40
