
一种扩繁烟蚜茧蜂的载体植物适合性评价
2015-12-09 09:12黄凤霞张礼生陈红印
环境昆虫学报 2015年4期
黄凤霞,周 阳,张礼生*,陈红印
(1.中国农业科学院植物保护研究所,国家农业生物安全中心,中国-美国生物防治实验室,北京 100081;2.湖南农业大学植物保护学院,长沙 410128)
烟 蚜 Myzus persicae (Sulzer) 属半翅Hemiptera 蚜科Aphididoe,分布于世界各地,我国南北各烟区普遍发生。其食性比较杂 (Emden et al.,1969;Blackman,1974;Weber,1985),我国已知烟蚜的寄主植物达170种以上。世界上已报道的寄主多达50 个科400 多种(Weber,1985),主要为茄科、十字花科、菊科和豆科植物。该虫除直接刺吸植株汁液,造成植株水分和营养失调而萎缩外,还可传播多种病毒病,其分泌蜜露可导致煤烟病,是烟草上的一种重要害虫(韩晓东等,1985;陈宝善等,1986;李凡和陈海如,2001;古勤生,2002;Veronique et al.,2010)。烟蚜的天敌种类很多,主要的类群有瓢虫、蚜茧蜂、食蚜蝇、草蛉、猎蝽、姬蝽、花蝽等。常见的种类有烟蚜茧蜂Aphidius gifuensis Ashmaed、七星瓢虫Coccinella septempunctata Linnaeus、异色瓢虫Harmonia axyridis (Pallas)、龟纹瓢虫 Propylea japonica (Thunberg)、中华草蛉Chrysoperla sinica Tjeder、大草蛉 Sympetrum Croceolum、丽草蛉Chrysopa formosa Brauer 等,对抑制烟蚜有较大作用。蚜茧蜂是专寄生蚜虫的天敌昆虫,以烟蚜茧蜂在我国主要烟草种植区应用最为广泛,在自然条件下有过防治桃蚜的成功例子 (忻亦芬,1982)。利用简易薄膜温室饲养蚜虫繁蜂并田间散放防治示范试验,表明在烤烟生长期,连续释放烟蚜茧蜂可有效地控制烟蚜的发生在防治水平以下。其效果比喷施45%乐果乳剂1000 倍液还优越(赵万源等,1980)。
烟蚜在不同寄主植物(桃树、烟草、油菜和甘蓝)上的存活率,若、成蚜发育历期,产仔率动态及内禀增长力均有明显差异 (刘绍友等,1999)。我国对烟蚜茧蜂种群的研究,先后有福建农学院赵修复、陈家骅,中科院昆明动物所赵万源、丁垂平,中国农科院植保所张礼生等,他们对烟蚜茧蜂的生物学、生态学、生理学以及应用等都曾有报道(赵修复,1978;陈家骅等,1996;赵万源等,1980;李玉艳等,2013;张礼生,2009,2014)。
平卧菊三七Gynura procumbens (Lour.)菊目,菊科三七属,俗称富贵菜、神仙草。平卧菊三七植株生长旺盛,分枝再生能力强,抗逆性强,生长适温20-30℃,较耐荫。在适温及阳光充足环境下,生长快速健壮。平卧菊三七的茎节部易生不定根,适宜扦插繁殖,从健壮的母茎上取成熟嫩茎扦插,插条长约10 cm 去除基部叶片插于苗床,遮荫保湿,10-15 d 可成活,以春秋两季容易生根(俞仁强和朱徐燕,2011)。目前常用来扩繁烟蚜茧蜂的寄主植物有烟草Nicotiana tabacum、茄子Solanum melongena、二年生萝卜Raphanus sativus 等,它们的生长周期均比较长。另外,烟草叶片表面常有烟蚜分泌的蜜露,全体被腺毛,对于后期的僵蚜收集带来了麻烦;茄子叶片耐蚜能力较弱,使用周期不长;二年生萝卜是指萝卜贮藏后次年盆栽发出的植株,其发出的叶片一般比正常生长的叶片小,且耐蚜能力较弱,使用周期较短。鉴于此,我们尝试寻找替代寄主植物,从而达到快速大量扩繁烟蚜茧蜂的目的。平卧菊三七生长习性符合我们对新寄主植物的要求,只是对扩繁烟蚜茧蜂的能力尚未见报道。为评估平卧菊三七作为载体扩繁烟蚜茧蜂的适合性,测试了烟蚜在平卧菊三七、以及适宜寄主植物烟草、茄子和二年生萝卜共4种寄主植物上的发育历期和繁殖力等,比较烟蚜在4种寄主植物上存活率和繁殖力的差异及不同寄主植物对于烟蚜茧蜂的扩繁效果。
1 材料与方法
1.1 材料
1.1.1 供试植物
供试植物为茄科的烟草(普通烟草)、茄子(园杂八号)、十字花科的二年生萝卜(卫青萝卜)和菊科的平卧菊三七。在温室育苗盘内培育烟草和茄子幼苗,待其长至3-4 片真叶时,移栽至营养钵(直径9 cm,高10 cm)内,采用新型混合肥料继续培育(方法下同),每盆一株。将贮藏于土窖中的萝卜,转栽至营养钵(直径15 cm,高20 cm)内,长出5 片青叶时待用。平卧菊三七为常年扦插繁殖,截取长势良好的10 cm 嫩茎,去掉下部叶片移至营养钵(直径9 cm,高10 cm)内,成活后待用。分别放置于养虫笼(40 cm×40 cm×50 cm)内作为供试植物,试验选用植物(5-8叶龄期)的上部叶片。
1.1.2 供试烟蚜
采自中国农业科学院植物保护研究所廊坊基地实验室烟草苗绿色型烟蚜,在温室内饲养烟蚜数代后,将足量均一化的烟蚜转接到四种寄主植物上连续饲养数代,待其完全适应后,烟蚜发育至3-4 龄时待用。
1.1.3 供试烟蚜茧蜂
烟蚜茧蜂采自中国农业科学院植物保护研究所廊坊基地实验室烟草苗,供试时在温室内以烟蚜作为寄主连续饲养10 代以上。
1.2 试验方法
1.2.1 不同寄主植物对烟蚜的发育历期和繁殖的影响试验
采用单头饲养法:饲养方法参照一种叶碟法(汤清波,2007),用供试植株苗期的幼嫩叶片饲养单头成蚜,24 h 后除去成蚜。将叶片背面朝上放入底层覆盖有湿润滤纸片的透明塑料培养皿(直径9 cm,高1 cm)中,用软毛笔将1 龄若蚜转移到叶片上,每皿1 头,每个寄主30 个重复。每天8∶00 和20∶00 各观察1 次,详细记载烟蚜的蜕皮、产仔量、死亡情况,直至所有试虫死亡为止。每次观察均剔除蜕皮和新产若蚜,隔天加水并换新鲜叶片。
1.2.2 不同寄主植物对烟蚜扩繁速度的影响试验
分别取4种寄主植物上的4 头烟蚜成蚜接在同类寄主植物上,为了便于计数蚜量,将四种寄主植物均去除心叶,只留3-4 片叶,并用养虫笼罩严,每2 d 观察1 次,连续观察10 d。统计每株苗上蚜虫总数,每处理调查5 株。
1.2.3 不同寄主植物对烟蚜茧蜂发育的影响试验
烟蚜茧蜂按1∶1 (雌∶雄)配对,按照1∶250 的蜂蚜比,释放在装有四种寄主植物的养虫笼内。从植株上出现僵蚜开始,逐日将其挑下,装入指形管内。取不同寄主植物上产生的24 h 内的烟蚜茧蜂僵蚜,称量百头质量。羽化后的成蜂继续配对,接在有烟蚜寄生的寄主植物上,置于温室内扩繁。同期,选取廊坊基地试验田的局部小麦田进行封闭管理,接种麦二叉蚜,后期释放适量烟蚜茧蜂,获取部分僵蚜,作为对照。
1.3 种群参数的计算
以d (x)为单位间隔,构建烟蚜在不同寄主植物上的实验种群生命表,按照以下公式计算各生命表参数:
净增殖率R0=∑lxmx;
平均世代周期T=∑xlxmx/∑lxmx;
内禀增长率rm=lnR0/T;
周限增长率λ=erm;
种群加倍时间t=ln2/rm=0.6931/rm
式中:x 为特定年龄,d;lx为在x 期间的存活率;mx为在x 期间平均每雌产仔数。
因蚜虫以孤雌生殖方式繁衍下一代,因此不设计性比这一栏目。
1.4 数据处理
试验数据统计分析均采用Excel 2003 和SAS 9.1 版本,进行ANOVA 方差分析,差异显著性检测采用邓肯氏新复极差法。
2 结果与分析
2.1 不同寄主植物对烟蚜发育历期的影响
由表1 可知,不同寄主植物对烟蚜的发育历期有显著影响。在4种寄主植物平卧菊三七、二年生萝卜、茄子和烟草上,1 龄若虫历期在平卧菊三七(2.24 d)上明显长于其他三种寄主;2 龄若虫历期在4种寄主植物上相差最大的为烟草(1.91 d)和茄子(1.85 d),平卧菊三七趋于平均值;3 龄若虫历期在平卧菊三七上最长为2.04 d,茄子上最短为1.71 d,相差0.33 d;4 龄若虫历期平卧菊三七上最短为1.75 d,而在二年生萝卜(1.97 d)和烟草(1.95 d)上相对比较长;总结若蚜期历期平卧菊三七上最长(7.92 d)与最短的茄子上(7.32d)相差0.59 d。烟蚜生殖前期在4种寄主植物上的差距较大,最长的为二年生萝卜(1.5 d),最短的为烟草(0.72 d);生殖期除在茄子(9.93 d)上最短外,在平卧菊三七、二年生萝卜和烟草上均超过13 d,分别为13 d、13.79 d、13.09 d;生殖后期在4种寄主植物上均为1.5 d 左右;成虫寿命在二年生萝卜(15.143 d)上最长,茄子(11.6 d)上最短。
综上,烟蚜在四种寄主植物上的世代历期,茄子上最短为18.92 d,而其他3种寄主植物上差异不显著,均为22 d 左右。

表1 4种寄主植物上烟蚜的发育历期(d)Table 1 The developmental duration of Myzus persicae (Suizer)on different host plants

图1 烟蚜在不同寄主植物上的产仔量Fig.1 The reproduction of Myzus persicae (Suizer)on different host plants
2.2 不同寄主植物对烟蚜繁殖力的影响
由图1 知,不同寄主植物对烟蚜产仔量的影响差异显著。在4种寄主植物上单头烟蚜的总平均产仔量在二年生萝卜上最多为63 头,其次在平卧菊三七上为52.33 头,茄子上和烟草上之间没有差异分别为42.67 头和41.27 头。

图2 烟蚜在不同寄主植物上日平均产仔量Fig.2 The progeny dynamic of Myzus persicae (Suizer)on different host plants
从图2 可以看出,从开始产仔的16 d 里,烟蚜在4种寄主植物上的日平均产仔量都有一个高峰期,其中平卧菊三七为3-10 d,二年生萝卜是3-11 d,茄子为3-9 d,烟草为4-12 d。烟蚜在4种寄主植物上产仔高峰期内日平均产仔量表现差异显著,平卧菊三七为5.10 头,二年生萝卜为5.79 头,茄子为4.81 头,烟草为4.62 头。综合比较产仔高峰期出现的时间早晚和产仔高峰期内日平均产仔量发现,平卧菊三七都趋于中间值,二年生萝卜最适合烟蚜寄生,茄子和烟草次之。
2.3 不同寄主植物对烟蚜扩繁速度的影响
从表2 中可以看到烟蚜在不同寄主植物上的繁殖速度差异显著。3 d 后的数据显示烟草更适合烟蚜初期产仔;5 d 后则是平卧菊三七上的烟蚜量最多,表明烟蚜在其上的适口能力较强;7 d 及之后的数据都表明二年生萝卜是最利于烟蚜快速繁殖的寄主植物,而和烟草相比,平卧菊三七上的烟蚜繁殖速度较快,茄子上的烟蚜繁殖速度较慢。
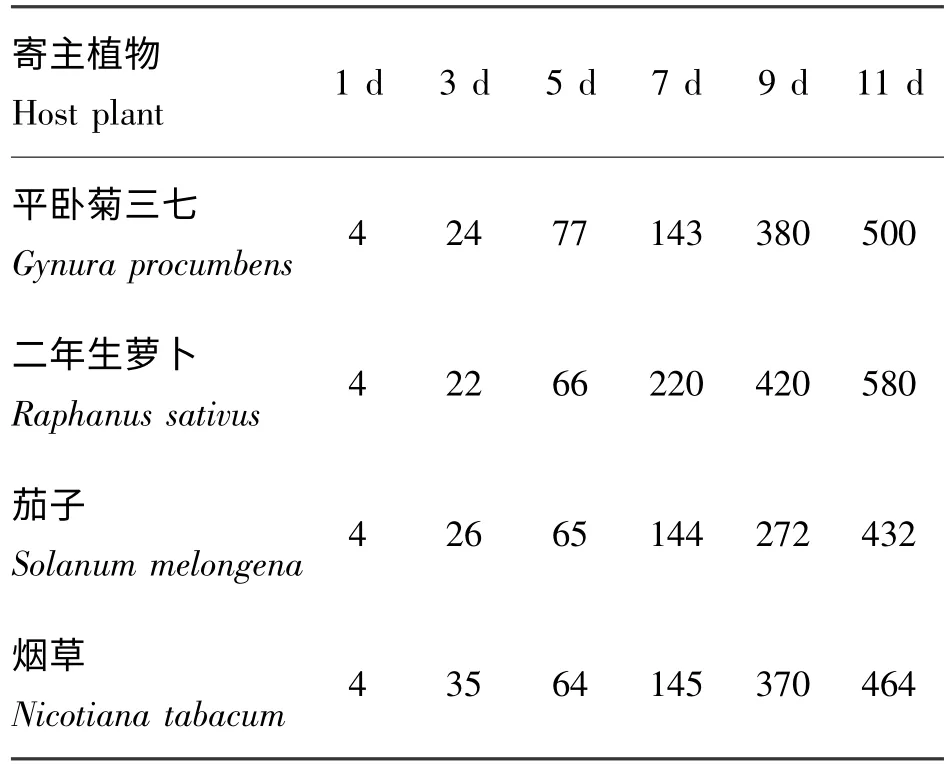
表2 烟蚜在不同寄主植物上的繁殖速度Table 2 The reproduction rate of Myzus persicae (Suizer)on different host plants
2.4 不同寄主植物对烟蚜茧蜂个体发育的影响
从图3 可以看出,不同的寄主植物对烟蚜茧蜂个体发育产生的影响差异显著。首先是小麦上的僵蚜明显比其他四种寄主植物上的僵蚜质量高,与二年生萝卜差距最大有两倍之多。其次是其他四种寄主植物之间对烟蚜茧蜂的发育也存在明显差异,主要是二年生萝卜上的烟蚜茧蜂个体发育较小,其他三种寄主植物上的烟蚜茧蜂个体发育之间差异不显著。

图3 不同寄主植物上取得烟蚜茧蜂的百头僵蚜质量Fig.3 The weight of one hundred mummies of Aphidius gifuensis taken from different host plants
2.5 主要生命表参数
由表3 可知,净增殖率R0,在二年生萝卜上最大(69.267),其次是平卧菊三七 (52.334),烟草上最小(41.262)。周限增长率λ 均大于1,表明烟蚜的种群呈几何型增长,按其值大小排序为茄子>二年生萝卜>平卧菊三七>烟草,其中平卧菊三七和二年生萝卜仅差0.001。内禀增长率rm表示烟蚜对寄主植物的适应度和嗜食性,其与λ值的趋势一致,表明相同条件下,烟蚜更易于取食茄子、二年生萝卜和平卧菊三七,该实验条件下的烟草有可能是抗虫品种。平均世代周期T 则是茄子最短(13.28 d),其他三种均大于14 d,种群加倍时间均在2.5 d 左右。综合评价,平卧菊三七在各个参数上都处于平均水平,可以作为替代载体植物来扩繁烟蚜及烟蚜茧蜂。

表3 烟蚜在不同寄主上的种群生命表参数Table 3 Life table parameters of Myzus periscae (Sulzer)on different host plants
3 结论与讨论
本研究结果表明,不同寄主植物对烟蚜发育和繁殖的影响存在显著差异,对烟蚜茧蜂的个体发育也存在显著差异。在不同的寄主植物上,烟蚜较喜欢取食二年生萝卜,这与忻亦芬等(2001)的研究结果一致,但是其上被寄生的烟蚜茧蜂的个体发育却不如其他3种寄主植物;烟蚜在茄子上的发育历期最短,但是其大量繁殖的速度最慢,这与宋春满等(2002)的研究结果一致;烟草是烟蚜最常见的易于扩繁烟蚜的寄主植物,马丽娜等(2006)研究结果表明烟蚜嗜食烟草强于十字花科。烟草生长周期长,从播种到接蚜虫往往需将近2 个月时间,其次,烟草生长需要较大空间,生产上常用的繁蜂小棚占地面积9 m2,一般可栽烟苗28 株(邓建华等,2006),另外,烟草叶片表面常有烟蚜分泌的蜜露且全体被腺毛,不利于后期僵蚜的收集;而比较4种寄主植物上烟蚜的发育历期时长为二年生萝卜>平卧菊三七>烟草>茄子,单头烟蚜的总平均产仔量为二年生萝卜>平卧菊三七>茄子>烟草,内禀增长率rm值为茄子>二年生萝卜>平卧菊三七>烟草,二年生萝卜上的烟蚜茧蜂个体平均发育较小,其他三种寄主植物上的烟蚜茧蜂个体发育之间差异不显著。综上说明,平卧菊三七可以用作大量扩繁烟蚜及烟蚜茧蜂的载体植物。
烟蚜对寄主植物的选择性是长期进化的结果。不同寄主植物上烟蚜的染色体核型有显著差异,染色体相对长度有一定差异(杨效文等,2000)。植物产生的次生代谢物对烟蚜的寄主选择性有显著影响,如烟草的次生代谢物——烟碱,与烟蚜的生长发育、存活率和繁殖成负相关(吴海军等,2003)。烟蚜对不同寄主的选择性也可能与不同寄主植物表皮的物理结构以及体内含有的营养物质有关,不同寄主植物的表皮结构及体内的营养物质有较大差异。这些都会直接或间接影响烟蚜的取食、生长发育、繁殖、种群动态、遗传基因等,进而影响烟蚜茧蜂的寄生、生长发育、遗传基因等。
平卧菊三七是一种易被蚜虫寄生的菊科植物,可满足蚜虫生长的营养需求;其有效承载叶片面积大,单株载蚜量高;叶片表面光滑,利于僵蚜收集;最重要的是,该植物可采用扦插方式进行扩繁,新枝条存活率高,成本低廉,易于操作,能节约大量的成本。故此,具备了大量扩繁烟蚜茧蜂时做为载体植物的潜力。本研究结果对于指导实验室用平卧菊三七快速大量扩繁烟蚜及烟蚜茧蜂有着开创性的启发,对于获取大量的烟蚜茧蜂僵蚜用于大田烟蚜的生物防治,这也是一种优良方法。
References)
Blackman RL.Life-cycle variation of Myzus persicae (Sulzer)in different part of the world,in relation to genotype and environment[J].Bulletin of Entomological Research,1974,63:595-607.
Chen BS,Gao QW,Luo XH.Identification of the causal viruses of tobacco mosaic in Guangdong Province [J].Chinese Journal of Virology,1986,2:166-174.[陈宝善,高乔婉,骆学海.广东烟草花叶病病原病毒的鉴定[J].病毒学报,1986,2:166-174]
Chen JH,Guan BB,Zhang YZ.A study of the relationship between Myzus persicae and Aphidius gifuensis[J].Acta Tabacaria Sinica,1996,1:8-12.[陈家骅,官宝斌,张玉珍.烟蚜与烟蚜茧峰相互关系研究[J].中国烟草学报,1996,1:8-12]
Deng JH,Wu XF,Song CM,et al.Rearing effect of Aphidius gifuensis with a nylon-net covered cages in tobacco fields[J].Journal of Southwest Agricultural University,2006,28 (1):66-73.[邓建华,吴兴富,宋春满,等.田间小棚繁殖烟蚜茧蜂的繁蜂效果研究[J].西南农业大学学报,2006,28 (1):66-73]
Emden HFV,Eastop VF,Hughes RD,et al.The ecology of Myzus Persicae[J].Annual Review Entomology,1969,14:197-270.
Gu QS.Advances in studies of cucurbits viral disease control[J].China Watermelon and Muskmelon,2002,4:115-117.[古勤生.葫芦科作物主要病毒病防治研究进展[J].中国西瓜甜瓜,2002,4:115-117]
Han XD,Chen RT,Li LS.An important virus of china nicotianatabacum-potato Y virus [J].Shanghai Agricultural Science and Technology,1985,2:20-21.[韩晓东,陈瑞泰,李林森.我国烟草一个重要病毒—马铃暮Y 病毒[J].上海农业科技,1985,2:20-21]
Li F,Chen HR.A new nicotianatabacum disease in growing nicotianatabacum areas where the west of Yunnan province —nicotianatabacum am disorder diseases[J].Acta Phytophylacica Sinica,2001,27 (4):50-51.[李凡,陈海如.云南西部烟区一种新的烟草病害—烟草丛枝症病害[J].植物保护学报,2001,27 (4):50-51]
Li YY,Zhang LS,Chen HY,et al.Temperature and photoperiodic response of diapause induction in Aphidius gifuensis[J].Chinese Journal of Applied Entomology,2013,50 (3):718-726.[李玉艳,张礼生,陈红印,等.烟蚜茧蜂滞育诱导的温光周期反应[J].应用昆虫学报,2013,50 (3):718-726]
Liu SY,Hou YM,Zhou JH,et al.A study on the adaptability of body-colour biotypes of the greenwich aphid,Myzus percise (Slugger)to host plants [J].Acta Agriculturae Boreali-Occidentalis Sinica,1999,8 (4):1-4.[刘绍友,侯有明,周靖华,等.桃蚜不同体色生物型的寄主适应性[J].西北农林学报,1999,8 (4):1-4]
Ma LN,Liu YH,Wang YJ,et al.Development and reproduction of Myzus persicae (Sulzer)as inflenced by different host plants[J].Journal of Southwest Agricultural University,2006,28 (1):74-76.[马丽娜,刘映红,王雅静,等.寄主植物对烟蚜生长发育和繁殖的影响[J].西南农业大学学报,2006,28 (1):74-76]
Song CM,Gao JH,Deng JH,et al.Effects of host plants on the activity of detoxicating enzymes and the target enzyme in Myzus persicae[J].Journal of Southwest Agricultural University,2002,24 (3):241-243.[宋春满,高家合,邓建华,等.寄主植物对云南烟蚜解毒酶和靶标酶活力的影响[J].西南农业大学学报,2002,24 (3):241-243]
Tang QB,Wang CZ.Leaf disc test used in caterpillar feeding preference study.[J].Chinese Bulletin of Entomology,2007,44 (6):912-915.[汤清波,王琛柱.一种测定鳞翅目幼虫取食选择的方法—叶碟法及其改进和注意事项[J].昆虫知识,2007,44 (6):912-915]
Veronique B,Maryline U,Baptiste M,et al.Aphids as transport devices for plant viruses[J].Comptes Rendus Biologies,2010,333:524-538.
Weber G.Genetic variability in host plant adaptation of the green peach aphid,Myzus persicae [J].Entomologia Experimentalis et Applicata,1985,38:49-56.
Wu HJ,Li YL,Pei SF.Influence of nicotine of the livability and growth rate of Myzus persicae [J].Journal of Shanxi Agricultural University,2003,(3):230-231.[吴海军,李友莲,裴淑芳.烟碱对桃蚜存活率和生长速率的影响[J].山西农业大学学报,2003,(3):230-231]
Xin YF,Li XR,Wang HP,et al.Studies on the use radish seedling to propagate peach aphid for mass rearing Aphidius gifuensis [J].Chinese Journal of Biological Control,2001,17 (2):49-52.[忻亦芬,李学荣,王洪平,等.用萝卜苗作桃蚜植物寄主繁殖烟蚜茧蜂[J].中国生物防治,2001,17 (2):49-52]
Xin YF.Researchprogress of Aphidius gifuensis Ashmead[J].Journal ofShenyang Agricultural University,1982,1:107-114.[忻亦芬.蚜茧蜂研究的进展[J].沈阳农学院学报,1982,1:107-114]
YangXW,Zhang SF,Zhang XY.Karyotype of green peach aphids from different host plants [J].Chinese Journal of Applied and Environmental Biology,2000,6 (1):56-60.[杨效文,张素方,张孝羲.不同寄主植物上烟蚜的染色体组型研究[J].应用与环境生物报,2000,6 (1):56-60]
Yu RQ,Zhu XY.Artificial cultivation techniques of rare vegetable-Gynura procumbens (Lour.)[J].Shanghai Vegetables,2011,2:77-78.[俞仁强,朱徐燕.保健珍稀蔬菜富贵菜人工栽培技术[J].上海蔬菜,2011,2:77-78]
Zhang LS,Chen HY,Zhang J,et al.Maternal effect of diapause in Aphidius gifuensis Ashmead [J].Chinese Journal of Applied Entomology,2014,51 (1):35-44.[张礼生,陈红印,张洁,等.滞育烟蚜茧蜂的亲代效应评价[J].应用昆虫学报,2014,51 (1):35-44]
Zhang LS.Application ofdiapause and hibernation in insects breeding.In:Zeng FR,Chen HY,eds.The Systems Engineering for Rearing Insect Natural Enemies [M].Beijing:China Agricultural Science and Technology Press,2009,54-89.[张礼生.滞育和休眠在昆虫饲养中的应用.见:曾凡荣,陈红印,主编.天敌昆虫饲养系统工程[M].北京:中国农业科学技术出版社,2009,54-89]
Zhao WY,Ding CP,Dong DZ,et al.The bionomics of Aphidius Gifuensis Ashmead and it's utilization for the control of tobacco aphid Myzus Persicae Sulzer[J].Zoological Research,1980,1 (3):405-414.[赵万源,丁垂平,董大志,等.烟蚜茧蜂生物学及其应用研究[J].动物学研究,1980,1 (3):405-414]
Zhao XF.A study on Chinese braconid wasps of the tribe spathiini(Hymenoptera:Braconidae,Doryctinae).Ⅰ[J].Acta Entomologica Sinica,1977,20 (2):205-216][赵修复.中国柄腹茧蜂记述(膜翅目:茧蜂科:矛茧蜂亚科:柄腹茧蜂族).Ⅰ[J].昆虫学报,1977,20 (2):205-216]
Zhao XF.A study on Chinese braconid wasps of the tribe spathiini(Hymenoptera:Braconidae,Doryctinae).Ⅱ[J].Acta Entomologica Sinica,1978,21 (2):173-184.][赵修复.中国柄腹茧蜂记述(膜翅目:茧蜂科:矛茧蜂亚科:柄腹茧蜂族).Ⅱ[J].昆虫学报,1978,21 (2):173-184]
猜你喜欢
湖北农业科学(2021年22期)2021-12-13
河南农业科学(2017年6期)2017-07-05
江苏农业科学(2017年7期)2017-05-23
西南农业学报(2017年4期)2017-05-17
中国稻米(2017年2期)2017-04-28
环境昆虫学报(2016年4期)2016-08-23
中国茶叶(2016年7期)2016-01-18
山东农业科学(2014年9期)2015-01-07
中国烟草学报(2012年3期)2012-04-10
中国烟草学报(2012年3期)2012-04-10
