
海南岛尖峰岭林区近50年的热量因子变化特征
2015-12-06 09:43周璋林明献李意德陈德祥骆土寿许涵
生态环境学报 2015年4期
周璋,林明献,李意德,陈德祥,骆土寿,许涵
中国林业科学研究院热带林业研究所//海南尖峰岭森林生态系统国家野外科学观测研究站,广东 广州 510520
海南岛尖峰岭林区近50年的热量因子变化特征
周璋,林明献,李意德*,陈德祥,骆土寿,许涵
中国林业科学研究院热带林业研究所//海南尖峰岭森林生态系统国家野外科学观测研究站,广东 广州 510520
全球气候变暖已得到充分证实。区域气候条件的变化必然影响该区域植物的生理活动,进而影响该区域植被的结构和功能。尖峰岭热带森林作为中国典型的热带森林生态系统之一,其长期的气候变化特征在评价森林对全球气候变化的响应与反馈中具有重要意义。采用国家野外科学观测研究站尖峰气象观测场1957─2005年的地面常规气象观测资料,选用气候趋势系数和气候倾向率定量分析了尖峰岭林区热量条件的变化特征,讨论热量条件的变化对主要热带森林植物的影响。结果表明,尖峰岭林区50年的多年平均气温和平均地温分别为24.7和29.0 ℃。50年来,林区热因子(平均温度和极端温度)均呈上升趋势,其中平均气温、平均最高地温、平均最低地温、极端最高气温、极端最低气温、极端最高地温升高趋势都显著(P<0.05),每10年分别增加0.17、0.29、0.35、0.30、0.53、1.93 ℃,尖峰岭热带林区气候变暖来自于最低气温和最高地温升高的贡献。1、2、8、10和11月的平均气温升高趋势显著(P<0.05),每10年分别增加0.27、0.28、0.14、0.12、0.24 ℃;1、2、3月的平均地温升高趋势显著(P<0.05),每10年分别增加0.47、0.82、4.56 ℃,旱季对年平均气温和地温的增温贡献值大于雨季。尖峰岭地处热带北缘,其有效积温远低于赤道地区,因此,变暖的气候条件将通过改变尖峰岭热带森林植物的物候期而影响其生长与繁殖。
热带森林;尖峰岭;气温;地温;气候趋势
IPCC(2013)第 5次报告指出,1880─2012年全球地表温度升高了0.65~1.06 ℃,1983─2012年可能是过去1400年中最暖的30年(中等信度),人为影响极有可能是造成观测到的 20世纪中叶以来气候变暖的主要原因。基于气候模型预测,与1986─2005年相比,2016─2035年期间全球平均表面温度可能升高0.3 ~0.7 ℃(中等信度。IPCC,2013)。
全球气候变化是不同区域气候变化的一个总体表现,因此区域气候变化是一个值得关注的问题。中国近 40年来的年均温变化趋势存在显著的转折点,全国平均升温开始于 1984年,增温率为0.058 ℃/a(Wang等,2010)。近50年来海南岛各气候区的温度总体呈上升趋势(增温率为0.024 ℃/a),并以东北区及西南区温度上升最为明显(许格希等,2013)。Malhi等(2002)的研究表明,热带森林地区气温平均升高 0.021~0.031 ℃/a。植物的各种生理活动,如光合呼吸作用、蒸腾作用、根系吸收水分和养分均受到温度的影响,同时温度变化能够引起环境其他因子,如湿度、土壤肥力的变化,从而影响植物的生长发育、产量和质量。温度是影响植被群落分布的主要因子,各气候带由于温度条件不同而分布着相应的树种和植被类型。温度通过影响生态系统的物质循环而影响生态系统的生产力。气候暖化作用势必造成气候植被带的迁移,造成植物群落结构、植物种类组成及植被生产力发生变化(Bonan等,2008)。
全球30%的陆地是森林,森林与气候之间存在着密切关系。大尺度上,森林水热环境决定于区域水热条件,并受气候变化的综合影响。植被与气候相关关系已经成为研究热点。森林气候资料不仅为森林生态学的各项研究如应用于森林树木生长的模拟(Fang等,2014)、水量平衡研究(Malhi等,2002)、物候研究(Gordo和 Sanz,2009)以及森林病虫害研究(Russo等,1993)等提供了基本数据,也为生态系统管理和生态系统模型的建立提供了基础数据(Kucharik等,2000),而且为全球变化对森林生态系统的影响及森林生态系统对全球变化的响应研究提供了基本依据。
目前对森林区短期气候分析(常杰等,1999;欧阳旭等,2014;冯静等,2012;张一平等,2002)及单气象要素分析较多(Morecroft等,1998;刘文杰等,2001;Hao等,2007),但对多年气候动态进行综合分析的资料却很少见,国内报道过小兴安岭五营红松林近50年(贺萍和孙勇,2008)、长白山阔叶红松林 22年(张弥等,2005)和西双版纳植物园50年的热量变化趋势研究(赵俊斌等,2012),而有关热带雨林区的长期气候动态研究极少,并且由于一般的气候资料来源于附近城市的气象站,不能充分地代表当地的森林气候。
热带森林是全球最重要的森林生态系统,其面积只占全球陆地面积12%,但是物种数占全球总物种数 50%以上;碳库占全球陆地生态系统的 40%(Phillips等,1998),占全球森林碳库的55%(Pan等,2011)。因此,热带雨林生态系统碳吸存的微小变化都将对生态系统乃至全球气候变化产生深刻的影响。尖峰岭地处热带北缘,被誉为“热带北缘生物物种基因库”(蒋有绪和卢俊培,1991),对气候变化敏感,是研究气候变化及其影响的理想天然实验室。本文对尖峰岭林区近 50年的热因子气候动态进行分析,补充该地区长期森林气候资料的不足,以期为揭示全球变化对陆地生态系统的影响过程与机理提供参考和依据。
1 研究地区与研究方法
1.1 自然概况
尖峰岭地区位于海南岛西南部的乐东和东方两县交界处,北纬 18°23′~18°50′,东经 108°36′~109°05′。尖峰岭林区面积为472.27 hm2,为海南岛5大林区之一。海南岛尖峰岭属低纬度热带岛屿季风气候,雨热同期,降水丰富,干湿两季明显(11月至翌年4月为旱季,5─10月为雨季)。成土母岩主要是花岗岩,土壤为砖红壤和砖黄壤。具有明显的气候、土壤和植被垂直带谱,地带性植被类型为热带常绿季雨林,而热带半落叶季雨林是尖峰岭地区海拔最低的森林植被类型(海拔60~350 m)。季雨林植被类型以龙脑香科(Dipterocaparceae)、大戟科(Euphorbiaceae)、茜草科(Rubiaceae)、蝶形花科(Papilionaceae)和桑科(Moraceae)等热带成分为主(李意德等,2002)。
1.2 研究方法
1.2.1 气象观测和气象要素统计
本文资料来自于海南岛尖峰岭森林生态系统国家野外科学观测试验站气象观测场 1957─2005年的地面常规气象资料(2006年后因站场移至其他地点而未统计其后的观测数据)。该气象观测场位于中国林业科学研究院热带林业研究所海南岛尖峰岭试验站内,海拔75 m,土壤为砖红壤。观测场按国家地面气象站标准设计:南北和东西边长 35 m×25 m,均质草皮地面,观测场四周距天然热带半落叶季雨林(次生林)2 m以上。
统计指标包括年平均气温、月平均气温、年平均最高和最低气温、年极端最高和最低气温、年积温、年平均地面温度、月平均地温、年平均最高和最低地温、年极端最高和最低地面温度以及地气温差等热量因子。为突出气候年代际的变化,对温度进行 5年滑动平均,以滤去年际振荡(魏凤英,1999)。
1.2.2 气候趋势系数和气候倾向率
有关气候趋势变化方面的分析,本文选用气候趋势系数(王绍武,1990)和气候倾向率(Jones,1998)进行定量分析,以研究气象要素在气候变化中升降的定量程度。气候要素趋势系数实际上为逐年的气候要素值(如温度)与年份序列数的相关系数。趋势系数为正(负)时,表示气候要素在所统计的时间内有线性增加(减少)的趋势。气候倾向率就是采用最小二乘法获得的气候要素值与时间的线性回归系数,其 10的倍数称为该要素的变化速率,其单位为要素单位/10a(例如:温度变化速率的单位为℃/10a)。
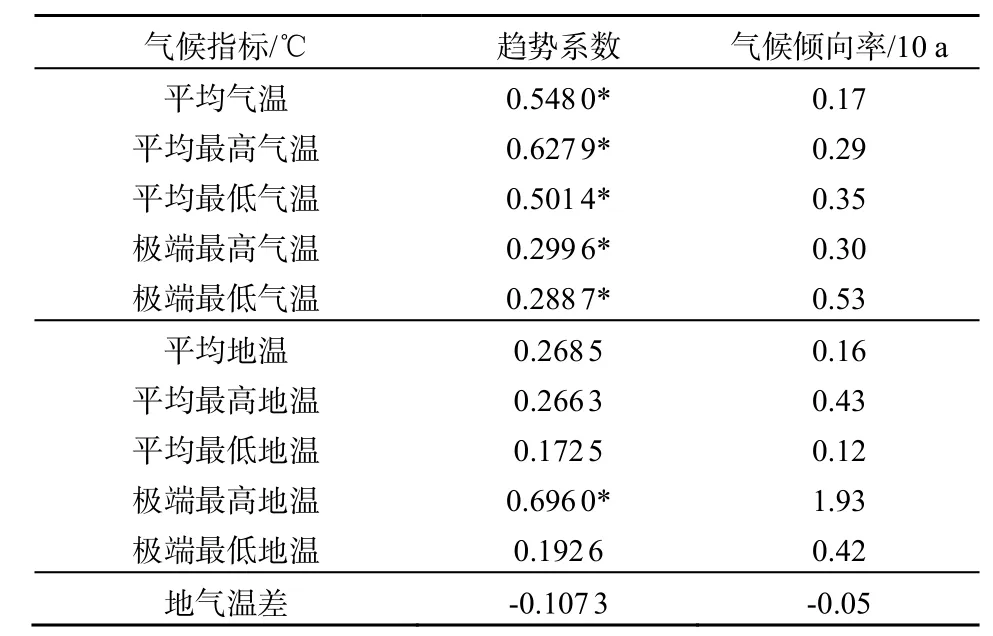
表1 尖峰岭热带山地雨林区热因子的趋势系数和气候倾向率Table 1 Climate trend coefficient and climate trend rate in Jianfengling tropical mountain rainforest
2 结果与分析
由表1可知,1957─2006年平均气温、平均最高气温、平均最低气温、极端最高气温、极端最低气温和极端最高地温总体都呈明显上升趋势(P<0.05),其中极端最高地温上升最明显,升温幅度为约1.93 ℃/10a。平均地温、平均最高地温、平均最低地温、极端最低地温都呈上升趋势,地气温差呈降低趋势,但均未通过P<0.05显著性检验。
2.1 气温变化
2.1.1 平均气温变化
如图1a所示,平均气温从1957年的24.3 ℃上升到2006年的25.4 ℃,其中上升幅度最大的是1997─2006年。尖峰岭林区50年的多年平均气温为24.7 ℃,其中1998年为平均气温最高的一年,其值为 25.9 ℃,这与海南岛和全国平均气温最高值出现年份一致;而 1971年为平均气温最低的一年,其值为23.7 ℃。
由图1a可知,1978年是很明显的一个转折点,在 1978年以前,该地区平均气温普遍比较低,为负距平,且始终在较小的范围内上下波动;以后气温就一直呈明显的上升趋势,除少数几年外,平均气温都为正距平,两阶段温差相距达 0.54 ℃。因此,近 50年海南岛尖峰岭林区地面气温的增暖主要发生在最近的25年内。
如图 1b所示,旱季平均气温年际波动明显比雨季剧烈,增温速率同样高于雨季,由此可见,旱季对年平均气温的增温贡献值大于雨季。
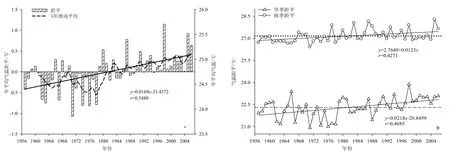
图1 1957─2006年年平均气温(a)、旱季雨季平均气温(b)的变化Fig. 1 Dynamics of annual mean air temperature(a), mean air temperature in dry and wet seasons(b) from 1957 to 2006
2.1.2 平均最高气温、最低气温变化
由表1可知,1957─2003年平均最高气温和平均最低气温都呈明显上升趋势,倾向率分别为0.29和0.35 ℃/10a,可见平均最低温度的上升幅度要大于平均最高温度,呈现非对称变化。
由图2可知,该地区近50年的年平均最高气温、最低气温的多年平均值分别为30.1、20.5 ℃。平均最高温度一直呈上升变化,而平均最低气温总体呈“V”型变化,在1977年为最低点,也是一个转折点,1977年之前大多为负距平,且一直呈下降趋势,特别60─80年代急剧下降;1977年之后大多为正距平,呈缓慢上升趋势,两阶段温度相差1.4 ℃,这说明了尖峰岭林区80年代后气温升高趋势明显加强。

图2 1957─2003年平均最高气温(a)、年平均最低气温(b)的变化Fig. 2 Dynamics of annual mean maximum air temperature(a) and mean minimum air temperature (b) from 1957 to 2003
2.1.3 极端最高气温、最低气温变化
由表1可知,1957─2005年极端最高气温和极端最低气温变化呈明显的升高趋势,气候倾向率分别为0.26和0.86 ℃/10a,极端最低气温上升速度较快。该林区 50年的年极端最高气温、最低气温的多年平均值分别为 36.7、8.8 ℃。年极端最低气温最低值出现在1974年,其值为 2.5 ℃;而年极端最高温最高值出现在1968年,其值为42.6 ℃。

图3 1957─2005年极端最高气温(a)、极端最低气温(b)的变化Fig. 3 Dynamics of annual extreme maximum air temperature(a) and extreme minimum air temperature (b) from 1957 to 2005
由图3可知,尖峰岭林区年极端最高、最低气温长期呈波动变化。除 1968年外,极端最高气温在年际间的变化都较为缓和,90年代以后大多数年份为正距平,表明年极端最高温度普遍偏高;而极端最低气温年际间波动比较剧烈,60─70年代普遍偏低,比多年平均偏低约3.0 ℃,80年代及其以后普遍偏高,比多年平均偏高约1.5 ℃。
2.1.4 月平均气温变化
由表2可知,月平均气温趋势系数和倾向率都为正值,表明月平均气温呈上升趋势。1、2、8、10和11月都表现出很强的正趋势,气温倾向率都在 0.2 ℃/10a左右,其中 8月上升幅度最大,达0.3 ℃/10a;5月的气温趋势系数和倾向率为最小,表现弱的正趋势;雨季的平均增温率略大于旱季。

表2 尖峰岭热带山地雨林月平均气温和月平均地温的趋势系数和气候倾向率Table 2 Climate trend coefficient and climate trend rate of monthly mean air and ground temperature in the forest region at Jianfengling
平均气温的季节变化整体呈抛物线趋势,1月是该地区一年中的最冷月,多年平均值为19.8 ℃;6月是最热月,多年平均值为28.3 ℃,为典型的热带季风气候。
2.2 地温变化
2.2.1 平均地温变化
由表1可知,1963─2003年平均地温的趋势系数和气候倾向率分别为0.2685和0.16 ℃/10a,说明 40年尖峰岭林区平均地温总体呈上升趋势,每10年上升0.16 ℃,平均气温增温速率接近。平均地温多年平均值为29.0 ℃,其中1972年为最低的一年,其值为 27.5 ℃,比年平均气温最低年份迟一年;与气温一样,1998年为年平均地温最高的一年,其值为30.9 ℃。
由图4a可知,1963─1978年(除1968年外),平均地温距平都低于或接近于 0,即平均地温处于较低时期,比多年平均值低出约0.6 ℃;80年代及其以后,平均地温处于较高的时期。与旱雨季平均气温变化一样,旱季对年平均地温升高的贡献大于雨季,如图4b。
2.2.2 平均最高地温、最低地温变化
由表1可知,1963─2003年平均最高地温和最低地温趋势系数分别为0.2663和0.1725,都呈上升趋势,增长率分别为0.43和0.12 ℃/10a,平均最高地温上升速度比平均地温高出0.27 ℃/10a,这说明尖峰岭林区地表温度的增加主要是由于最高地温升高造成的。
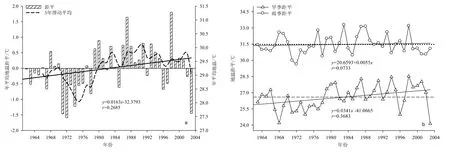
图4 1962─2003年年平均地温(a)、旱季雨季平均地温(b)的变化Fig. 4 Dynamics of annual mean ground temperature(a), mean ground temperature in dry and wet seasons(b) from 1962 to 2003
如图5所示,年平均最高地温和最低地温年际间波动都比较大。由 5年滑动平均线可以看出,1963─1976年除1964年外,平均最高地温都为负距平,表明该时期最高地温处于较低时期;1976年以后滑动平均距平大多在0以上,表明该时期最高地温处于高温时期。平均最低地温在1963─2003年之间呈现中间低两端高的变化特点,即70年代和80年代处于较低时期,60、90年代及后来处于较高时期。

图5 1963—2003年平均最高地温(a)、年平均最低地温(b)的变化Fig. 5 Dynamics of annual mean maximum ground temperature(a) and mean minimum ground temperature (b) from 1963 to 2003
2.2.3 极端最高地温、最低地温变化
由表1可知,1963─2003年极端最高地温变化呈显著上升趋势,每10年上升1.93 ℃;极端最低地温呈上升趋势,每10年上升0.42 ℃。40年间的年极端最高地温、最低地温的多年平均值分别为68.8、8.6 ℃。历年极端最高地温的最高值出现在1995年,其值为76.1 ℃,历年极端最低地温的最低值出现在1974年,其值为2.6 ℃(图6)。

图6 1963─2003年极端最高地温(a)、极端最低地温(b)的变化Fig. 6 Dynamics of annual extreme maximum ground temperature(a), and extreme minimum ground temperature (b) from 1963 to 2003
2.2.4 月平均地温变化
由表2可知,月平均地温的趋势系数在5月份为负值,每10年降低0.3 ℃;其余月份都为正值,即呈升高趋势,其中,1—3月呈显著上升趋势,尤其是3月份达4.5 ℃/10a。这说明了尖峰岭地表温度的升高是由于冬季地表温度的显著升高所造成的。平均地温的季节变化呈抛物线,最小值与气温一样,出现在1月,其值为17.4 ℃;最大值出现在5月,其值为34.2 ℃。
2.3 地气温差变化
从表1和图7a可知,1962─2003年地气温差呈微弱下降趋势,每10年下降0.05 ℃。地气温差在上世纪70年代初和2000年以后为较小时期,80和 90年代大多数年份为正值,即地温高于气温,热量是由地面输往大气;最大值和最小值分别出现在1987和2003年。
由气温ta、0 cm地温ts和地气温差ts-ta的多年平均值季节变化可以看出(图7b),气温和0 cm地温曲线都呈抛物线型,变化趋势相同,最高值出现在雨季的5─6月,最低值出现在旱季的12月至翌年1月。地气温差曲线明显不同于气温和地温,最大值和最小值分别出现在雨季的5月和8月,最大值出现的时间与全国(6─7月)相比,提前了2~3个月;最小值与全国(12月至翌年1月)相比,提前了3~4个月。

图7 尖峰岭热带山地雨林地气温差年代际变化(a),多年平均气温、0 cm地温和地气温差的季节变化(b)Fig. 7 The interdecadal changes (a) of (ts-ta) and the annual variation (b) of (ta, ts, and ts-ta) in Jianfengling tropical mountain rainforest
3 讨论与结论
3.1 讨论
本研究表明,在全球变暖背景下,尖峰岭林区也出现了温度升高的趋势,平均气温上升 0.17 ℃/10a,平均地温上升0.16 ℃/10a,均低于全球热带地区0.21~0.31 ℃/10a(Malhi和Wright,2004)、中国 0.25 ℃/10a(Wang等,2010)以及海南岛0.24 ℃/10a(许格希等,2013)的气温升高速率。中国南方地区增温率不如北方地区高是其原因之一(于淑秋等,2003);另外,尖峰岭林区由于比较偏远落后,工农业温室气体排放较少,且森林覆盖面积较大,故其增温率相对较低。根据已有的森林长期气候资料来看,尖峰岭林区气温增长率低于中国北方针叶林区的气温增长率(0.38 ℃/10a。王宇等,2008),略高于西双版纳植物园(0.13 ℃/10a。赵俊斌等,2012),然而,造成这种变化格局的机制,还有待于进一步深入研究。
地气温差的变化趋势基本上反映了地表感热通量(包括水汽和热量)的变化特征,而感热通量又是低层大气的主要热量来源之一,也是地面热量平衡的重要分量,因此,地气温差是决定小气候和区域气候的重要因子。有研究发现青藏高原春季地气温差对夏季的降雨有一定的指示意义,即地气温差越大则降雨量越大(江灏和王可丽,2000)。对西双版纳的研究表明,地气温差值在过去 50年呈下降趋势是导致该地区降雨量减少的原因(赵俊斌等,2012)。尖峰岭林区地气温差季节变化明显,在刚进入雨季的5月份达到最大;从年际间变化看,地气温差值在过去的半个世纪里呈下降趋势,这也是尖峰岭热量条件变化趋缓和波动变小的必然结果。地气温差的最大值和最小值都比全国平均出现时间要提前2~4个月,这可能是因为8月为该区降水最丰月(约占全年降水量的 20%左右),过多的云雨天气大大削弱了到达地面的太阳辐射强度,因而地温与气温也受到影响。其中,地温受其影响降温效应更显著,从而导致地温与地气温差最大值相差3.0 ℃左右。
从各热量因子变化趋势来看,尖峰岭林区最低气温增暖趋势高于平均气温,这与中国总体气温变化趋势一致(任国玉等,2005)。同时平均最低气温的上升幅度要大于平均最高气温,平均最高地温的上升幅度要高于平均最低地温,都呈现非对称变化。这一结果与其他一些研究所得结论一致(王菱等,2004;郑艳等,2005)。这也说明了尖峰岭林区气候变暖主要是最低气温和最高地温的显著升高造成的。
尖峰岭地处亚洲热带北缘地区,气温比典型的热带地区低,热带植物(尤其是东南亚典型热带龙脑香科植物)的生长与繁殖受该地低温影响较大(粟娟和王德祯,1994;许格希等,2014)。该研究表明,尖峰岭林区年极端最低气温和平均最低地温均呈现显著上升的趋势(分别为 0.53、0.35 ℃/10a),这对该地区热量不足起到一定的补偿作用。
气温上升必然对生物的各种活动与行为产生影响,例如,在很多地区发现植物春季物候提前、秋季物候期推迟、生长季延长(Chen等,2005;Ma和Zhou,2012;Piao等,2006)。许格希等(2014)对海南岛尖峰岭热带树木园(本研究的气象观测场附近)的 12种热带常绿阔叶乔木展叶期与开花期对气候变化的响应研究表明,多数树种展叶始期受展叶前冬季及春季气温影响显著,所有的树种展叶始期与开花始期的发生都受到气温和降水的共同影响。由此可推断,如果仅从热量因子的变化趋势看,尖峰岭林区的热量因子变化趋势有利于热带植物的生长与繁殖,但由热量所导致的其他环境因子的变化(如降雨量减少),则可能对尖峰岭林区生长的植物造成一定的负面影响(许格希等,2014)。全球不同生态系统增温实验综合分析得出,通过2~9年增温 0.3~6.0 ℃,土壤呼吸增加 20%,氮矿化速率增加46%,植物生产力增加19%(Rustad等,2001)。
3.2 结论
对于尖峰岭林区近 50年热量因子变化趋势,从年际变化来看,平均气温、平均最高地温、平均最低地温、极端最高气温、极端最低气温、极端最高地温升高趋势显著(P<0.05),且最低气温和最高地温增温幅度最大,说明尖峰岭热带林区气候变暖来自于最低气温和最高地温升高的贡献。从季节变化来看,1、2、8、10、11月的平均气温升高趋势显著(P<0.05),1、2、3月的平均地温升高趋势显著(P<0.05),说明旱季对年平均气温和地温的增温贡献值大于雨季。
影响尖峰岭林区近 50年热量因子变化的因素有外部因素和内部因素,外部因素主要是指太阳活动的影响,内部因素主要是大气环流(包括ENSO事件、台风等)和人为干扰(尖峰岭上世纪 80年代的森林砍伐、人工林的种植等),这些因素往往在尖峰岭气候变化中占主导地位。然而,这些因素是如何影响尖峰岭林区气候变化的,还有待于今后的进一步研究。
BONAN G B. 2008. Forest and climate change: forcings, feedbacks, and the climate benefits of forests [J]. Science, 320(5882): 1444-1449.
CHEN X Q, HU B, YU R. 2005. Spatial and temporal variation of phonological growing season and climate change impacts in temperate eastern China [J]. Global Change Biology, 11(7): 1118-1130.
FANG J Y, KATO T, GUO Z D, et al. 2014. Evidence for environmentally enhanced forest growth [J]. Proceedings of the National Academy of Sciences, 111(26): 9527-9532.
GORDO O, SANZ J J. 2009. Long-term temporal changes of plant phenology in the Western Mediterranean [J]. Global Change Biology, 15(8): 1930-1948.
HAO Y B, WANG Y F, HUANG X Z, et al. 2007. Seasonal and interannual variation in water vapor and energy exchange over a typical steppe in Inner Mongolia, China [J]. Agricultural and Forest Meteorology, 146(1-2): 57-69.
IPCC, 2013: Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change [M]. Cambridge: Cambridge University Press.
JONES P D. 1988. Hemispheric surface air temperature variations: recent trend and an update to 1987 [J]. Journal of climate, (1): 654-660.
KUCHARIK C J, FOLEY J A, DELIRE C, et al. 2000. Testing the performance of a dynamic global ecosystem model: water balance, carbon balance, and vegetation structure [J]. Global Biogeochemical Cycles, 14(3): 795-825.
MA T, ZHOU C. 2012. Climate-associated changes in spring plant phenology in China [J]. International journal of biometeorology, 56(2): 269-275.
MALHI Y, WRIGHT J. 2004. Spatial patterns and recent trends in the climate of tropical rainforest regions [J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 359(1443): 311-329.
MALHI Y, PEGORARO E, NOBRE A D, et al. 2002. The water and energy dynamics of a central Amazonian rain forest [J]. Journal of Geophysical Research: Atmospheres, 107 (D20): LBA 45-1–LBA 45-17.
MORECROFT M D, TAYLOR, M E, OLIVER H R. 1998. Air and soil microclimates of deciduous woodland compared to an open site [J]. Agricultural and Forest Meteorology, 90(1-2): 141-156.
PAN Y D, BIRDSEY R A, FANG J Y, et al. 2011. A large and persistent carbon sink in the world's forests [J]. Science, 333(6045): 988-993.
PHILLIPS O L, MALHI Y, HIGUCHI N, et al. 1998. Changes in the carbon balance of tropical forests: Evidence from long-term plots [J]. Science, 282(5388): 439-442.
PIAO S L, FANG J Y, ZHOU L M, et al. 2006. Variations in satellite-derived phenology in China's temperate vegetation [J]. Global Change Biology, 12(4): 672-685.
RUSSO J M, LIEBHOLD A M, KELLEY J G. 1993. Mesoscale weather data as input to a gypsy moth (Lepidoptera: Lymantriidae) phenology model [J]. Journal of Economic Entomology, 86(3): 838-844.
RUSTAD L E, CAMPBELL J L, MARION G M, et al. 2001. A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming [J]. Oecologia, 126(4): 543-562.
WANG S P, WANG Z H, PIAO S L, et al. 2010. Regional differences in the timing of recent air warming during the past four decades in China[J]. Chinese Sci Bull, 55(19): 1968-1973.
常杰, 潘晓东, 葛滢, 等. 1999. 青冈常绿阔叶林内的小气候特征[J]. 生态学报, 19(1): 68-75.
冯静, 段文标, 陈立新. 2012. 阔叶红松混交林林隙大小和林隙内位置对小气候的影响[J]. 应用生态学报, 23(7): 1758-1766.贺萍, 孙勇. 2008. 小兴安岭五营林区近50a的气候动态[J]. 黑龙江气象, 25(S1): 1-3.
江灏, 王可丽. 2000. 青藏高原地表热状况的卫星资料分析[J]. 高原气象, 19(3): 323-330.
蒋有绪, 卢俊培. 1991. 中国海南岛尖峰岭热带林生态系统[M]. 北京:科学出版社.
李意德, 陈步峰, 周光益, 等. 2002. 中国海南岛热带森林及其生物多样性保护研究[M]. 北京: 中国林业出版社.
刘文杰, 张克映, 王昌命, 等. 2001. 西双版纳热带雨林干季林冠层雾露形成的小气候特征研究[J]. 生态学报, 21(3): 48-49.
欧阳旭, 李跃林, 张倩媚, 等. 2014. 鼎湖山针阔叶混交林小气候调节效应[J]. 生态学杂志, 33(3): 575-582.
任国玉, 郭军, 徐铭志, 等. 2005. 近 50年中国地面气候变化基本特征[J]. 气象学报, 63(6): 942-956.
粟娟, 王德祯. 1994. 海南岛尖峰岭热带树木园主要树种的物候研究[J].林业科学研究, 7(3): 294-300.
王菱, 谢贤群, 苏文, 等. 2004. 中国北方地区 50年来最高和最低气温的变化和影响[J]. 自然资源学报, 19(3): 337-343.
王绍武. 1990. 近百年我国及全球气温变化趋势[J]. 气象, 16(2): 11-16.
王宇, 周广胜, 贾丙瑞. 2008. 1954—2005年中国北方针叶林分布区的气候变化特征[J]. 应用生态学报, 19(5): 942-948.
魏凤英. 1999. 现代气候统计诊断与预测技术[M]. 北京: 气象出版社: 42-59.
许格希, 郭泉水, 牛树奎, 等. 2013. 近50a来海南岛不同气候区气候变化特征研究[J]. 自然资源学报, 28(5): 799-810.
许格希, 罗水兴, 郭泉水, 等. 2014. 海南岛尖峰岭 12种热带常绿阔叶乔木展叶期与开花期对气候变化的响应[J]. 植物生态学报, 38(6): 585-598.
于淑秋, 林学椿, 徐祥德. 2003. 中国气温的年代际振荡及其未来趋势[J]. 气象科技, 31: 136-139.
张弥, 关德新, 韩士杰, 等. 2005. 长白山阔叶红松林近 22年的气候动态[J]. 生态学杂志, 24(9): 1007-1012.
张一平, 刘玉洪, 马友鑫, 等. 2002. 热带森林不同生长时期的小气候特征[J]. 南京林业大学学报: 自然科学版, 26(1): 83-87.
赵俊斌, 张一平, 宋富强, 等. 2012. 西双版纳热带植物园热量变化趋势[J]. 南京林业大学学报:自然科学版, 36(1): 47-52.
郑艳, 张永领, 吴胜安. 2005. 海口市气温变化及最高最低气温的非对称变化[J]. 气象, 31(7): 28-31.
Change Characteristics of the Heat Factors in Tropical Forests in Jianfengling, Hainan Island during the Past 50 Years
ZHOU Zhang, LIN Mingxian, LI Yide, CHEN Dexiang, LUO Tushou, XU Han
Research Institute of Tropical Forestry, Chinese Academy of Forestry//Jianfengling Tropical Forest Ecosystem Station, Guangzhou 510520, China
Global climate warming has been confirmed, and region climate change would impact on the physiological activities of plants, and then have an influence on the structure and functions of vegetation. Tropical forests in Jianfengling, Hainan Island is one of the typical tropical forest ecosystems. Its long term climatic dynamics is important to assess the response and adaption of forest to global change. Long-term data (1957─2005) of air temperature and soil surface temperature in Jianfengling tropical forest area were used to analyze the inter-annual trends of heat condition and discuss the influences on the tropical plants. The results showed that, annual mean air/ground temperature during past 50 years was 24.7and 29.0 ℃. Annual mean air/ground temperature, annual mean maximum and minimum air/ground temperature, annual extreme maximum and minimum air/ground temperature, and annual accumulated temperature have been rising in the past 50 years over the Jianfengling forest. Annual mean air, annual mean maximum/minimum air temperature, extreme maximum/minimum air temperature and extreme maximum ground temperature increased significantly, having a increase of 0.17, 0.29, 0.35, 0.30, 0.53, 1.93 ℃ per 10 year .These facts indicate that the change of the minimum temperature is more sensitive than that of the maximum temperature. Monthly mean air temperature in January, February, August, October and November increased significantly, having a increase of 0.27, 0.28, 0.14, 0.12, 0.24 ℃ per 10 year. Monthly mean ground temperature in January, February and March increased significantly, having a increase of 0.47, 0.82, 4.56 ℃per 10 year. It is concluded that the main contributor of climate warming in Jianfengling tropical rainforest is the rising of the minimum air temperature and maximum ground temperature. The rising temperature in dry seasons made a greater contribution to the increase of air temperature and ground temperature. As Jianfengling is located in the north verge of tropical region, its rising temperature will benefit the tropical plants living there.
tropical forests; Jianfengling; air temperature; ground temperature; climatic variation tendency
10.16258/j.cnki.1674-5906.2015.04.005
X16;S716
A
1674-5906(2015)04-0575-08
周璋,林明献,李意德,陈德祥,骆土寿,许涵. 海南岛尖峰岭林区近 50年的热量因子变化特征[J]. 生态环境学报, 2015, 24(4): 575-582.
ZHOU Zhang, LIN Mingxian, LI Yide, CHEN Dexiang, LUO Tushou, XU Han. Change Characteristics of the Heat Factors in Tropical Forests in Jianfengling, Hainan Island during the Past 50 Years [J]. Ecology and Environmental Sciences, 2015, 24(4): 575-582.
国家生态系统观测研究网络运行服务(2014-2015);国家林业公益性行业科研专项(201404201);国家林业科技创新平台运行补助(2014)
周璋(1983年生),男,博士,主要从事生态气候和森林生态系统碳氮循环研究。E-mail:zhouzhang315@126.com *通信作者
2015-03-23
猜你喜欢
山西林业(2021年2期)2021-07-21
新农民(2019年2期)2019-08-16
天津科技(2019年3期)2019-03-30
安徽农业科学(2019年6期)2019-03-27
红土地(2018年8期)2018-09-26
农家科技下旬刊(2017年12期)2018-04-16
现代园艺(2018年2期)2018-03-15
现代农业科技(2018年22期)2018-01-15
农家科技下旬刊(2017年9期)2017-11-12
初中生学习·高(2016年8期)2016-05-14
