
基于拮抗试验的杏鲍菇菌株分类研究
2015-12-02 04:59:53刘盛荣张维瑞柯斌榕刘新锐吴小平
食药用菌 2015年1期
刘盛荣 张维瑞 柯斌榕 刘新锐 吴小平*
基于拮抗试验的杏鲍菇菌株分类研究
刘盛荣1张维瑞1柯斌榕2刘新锐2吴小平2*
(1.宁德师范学院生物系,福建 宁德 352100;2.福建农林大学生命科学学院 福建农林大学菌物研究中心,福建 福州 350002)
利用拮抗反应对全国各地收集的81株杏鲍菇进行分类研究。结果显示,81个菌株可分为11个类群,最大群包含25个菌株,最小群仅含1个菌株。从中显示我国杏鲍菇菌株既具有丰富的遗传多样性,又可能存在严重的同物异名现象。
杏鲍菇;拮抗反应;分类;遗传多样性
杏鲍菇()又名刺芹侧耳,属担子菌亚门、层菌纲、伞菌目、侧耳科、侧耳属,是原产于欧洲南部、北非、中亚等地的一种品质优良的大型伞菌[1]。常发生于伞形花科刺芹属刺芹枯死的枝和根的周围。在我国新疆、四川、青海等地有少量分布。杏鲍菇具有味道鲜美、鲜品货架期长等优点。我国于20世纪90年代引种并进行试验性栽培,目前除利用自然季节规模栽培外,工厂化设施栽培在上海、广东、福建、山东等地区也已较为普遍。
科研人员对杏鲍菇的营养生理[2,3]、工厂化栽培[4~6]、瓶栽[7]、育种[8~10]等方面进行了深入研究,其遗传多样性研究也有不少报道。如尚晓东等[11]利用RAPD分析19株杏鲍菇的遗传关系,贺冬梅等[12]通过酯酶同工酶确定不同杏鲍菇的亲缘关系,冯伟林等[13]以ISSR指纹图谱研究杏鲍菇遗传差异。但这些研究所收集的菌株数量较少,难以真实反映我国杏鲍菇整体情况。因此收集大量菌株进行系统研究十分必要。为评估杏鲍菇菌种的遗传多样性,本研究依托福建农林大学菌物研究中心的菌种优势,利用拮抗反应对从全国各地收集的81个杏鲍菇菌株进行分类研究,以期为杏鲍菇的遗传研究、育种以及菌种管理提供参考。
1 材料与方法
1.1 供试材料
供试菌株及来源见表1。采用PDA培养基:马铃薯200 g、葡萄糖20 g、琼脂20 g、水1 000 mL,121 ℃灭菌15 min。
1.2 方法
(1)菌种活化。将4 ℃冰箱保存的供试菌株取出后重新接种于新鲜斜面,于28 ℃生化培养箱培养7天,作为活化菌株用于后续实验。
(2)拮抗对峙培养。将融化至温度约50 ℃的PDA培养基15~20 mL倒入直径9 cm的培养皿,凝固后随机抽取1个菌株作为测试菌株,用接种针从活化的斜面中挑取黄豆大小的琼脂块菌丝,面朝下接种至培养皿中部;相距1 cm处接种另一菌株,封口倒置于28 ℃生化培养箱中培养,约7 天后观察菌丝接触后两者的拮抗反应并拍照。
与测试菌株无拮抗反应或仅有微弱拮抗反应的菌株作为一个类群(包括测试菌株在内),不转入下一轮拮抗实验;与测试菌株有拮抗反应的所有菌株转入下一轮实验,从这些菌株中随机抽取1个菌株作为测试菌株,与其余菌株进行拮抗对峙培养,依次类推,直至将所有菌株都作出分类(图1)。
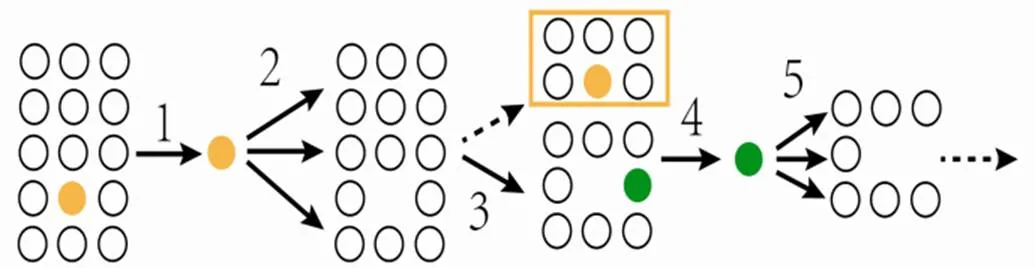
图1 拮抗实验示意图
注:1,随机选取1个菌株作为测试菌株;2,测试菌株与其余菌株分别一一配对;3,与测试菌株无拮抗反应或呈微弱反应的为一群(包含测试菌株),其余菌株进入下一轮实验;4,再从中随机选取1菌株为测试菌株;5,测试菌株与其余菌株分别一一配对,依次类推直至所有菌株分群完。
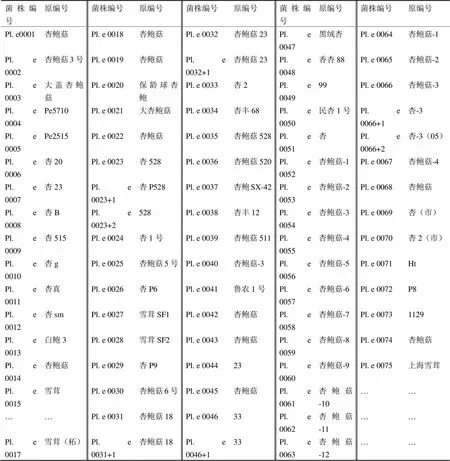
表1 供试菌株
注:菌株来源Pl. e 0001-5(福建省轻工业研究所);Pl. e 0006-15、17(福建农科院蚕桑所);Pl. e 0018[李晓宇赠(日本产品分离)];Pl. e 0019(深圳罗湖食用菌厂);20、21(东北食药用菌研究所);Pl. e 0022(姜堰市富康食用菌研究所);Pl. e 0023、24(江都市天达食用菌研究所);Pl. e 0023+1、26-29(贵州习酒食用菌研究所);Pl. e 0023+2、46+1(湖南农业大学食用菌研究所);Pl. e 0025、30(华中农业大学菌种中心);Pl. e 0031、31+1、32(古田戴维浩);Pl. e 0032+1、44、45(古田科兴食用菌研究所);Pl. e 0033(保定冀微覃源);Pl. e 0034-41(寿光市食用菌研究所);Pl. e 0042(四川省农科院微生物实验室);Pl. e 0043(泌阳县食用菌研究所);Pl. e 0046-49(嘉鱼县环宇食用菌研究所);Pl. e 0050(古田科协为民食用菌场);Pl. e 0051(锦州亚泰食用菌研究所);Pl. e 0052-63(龙海九湖食用菌研究所);Pl. e 0064-67(三明真菌研究所);Pl. e 0066+1、66+2、68-71(福州市农科所食用菌室);Pl. e 0072、73(锦州亚泰食用菌研究所);Pl. e 0074、75(福建省农科院植保所)。
2 结果与分析
2.1 杏鲍菇菌株间的拮抗反应
经7~15 天培养,所有供试杏鲍菇菌株的菌丝表现洁白、长势好。不同菌株间菌丝接触后的典型拮抗反应有3种表现(图2):(1)菌丝间无拮抗反应;(2)拮抗反应微弱(少数);(3)拮抗反应明显,菌丝间形成明显沟状。笔者将与测试菌株无拮抗反应或反应微弱的菌株视为同一类群。

图2 典型的杏鲍菇菌株间拮抗反应
2.2 81个杏鲍菇菌株的分群
拮抗反应依次以8、35、50、5、52、23、71、40、67、4为测试菌株,进行了10轮实验,结果供试的81株杏鲍菇可分成11个类群,各群所包含的菌株数相差悬殊,其中最大的类群包含25个菌株,最小的群有3个,各自仅含1个菌株,详细分群结果如表2所示。

表2 81株杏鲍菇分群
注:“a”本轮测试菌株。
3 讨 论
真菌的拮抗反应是体细胞不亲和的直接体现[14,15]。食用菌是具有重要经济价值的一类真菌,体细胞的不亲和性现象同样普遍存在。由于拮抗分类鉴定具有简单、快速、直观等优点,因此是最常用的食用菌菌种鉴定方法。诸多研究显示,该方法具有较高的可靠性,如黄伞的拮抗反应分类结果与以酯酶同工酶的分类方法相一致[16],金针菇拮抗分类与RAPD分析结果吻合[17]。此外,白灵菇[18]、真姬菇[19]、姬松茸[20]、香菇[21]、灵芝[22]等拮抗分类鉴定结果与其他鉴定方法的分类结果也相吻合。
本研究依据拮抗反应将81株杏鲍菇分为11个类群。福建农林大学菌物研究中心林骥[23]采用SCAR分子标记技术结合拮抗反应对其中72株杏鲍菇进行分类,结果供试菌株分成了59类,显示出更高的鉴别能力。此外,研究者认为其中13株为同种异名菌株(占所有菌株的18.06%),这与本研究结论有一定相似性。总之,杏鲍菇菌株在我国具有丰富的遗传多样性,通过基因工程、细胞融合或杂交等技术将不同的种质进行合理化组合,可预期获得符合特定需求的杏鲍菇新菌株。
值得注意的是,有3个群各自仅含有1个菌株,推测可能的原因如下:(1)这些菌株不是主要的商业性栽培品种,使得相互间引种以及被菌业工作者进行组织分离的可能性小,其在菌种市场的流通较少;(2)这些菌株与其他大多数杏鲍菇菌株相比,存在独特的遗传差异。此外,本实验其中一个类群包含25个菌株,这个群可能存在较多同物异名菌株,是由于主栽品种相互引种、组织分离、随意命名所致。
[1] Obatake Y, Murakami S, Matsumoto T, et al. Isolation and characterization of a sporeless mutant in[J]., 2003, 44: 33-40.
[2] 马璐, 杜双田, 金凌云, 等. 杏鲍菇营养生理研究[J]. 西北农林科技大学学报(自然科学版), 2010, 38(9): 129-134.
[3] 王振河, 武忠伟, 王斌, 等. 杏鲍菇菌丝营养生理特性研究[J]. 河南师范大学学报(自然科学版), 2007, 35(4): 139-143.
[4] 王瑞娟, 郭力刚, 刘朝贵, 等. 工厂化栽培杏鲍菇优良菌株筛选[J]. 食用菌学报, 2006, 13(3): 19-21.
[5] 姜性坚, 王春晖, 胡汝晓 ,等. 杏鲍菇工厂化生产关键技术的研究[J]. 中国食用菌, 2011, 30(1): 23-25, 41.
[6] 徐全飞, 郭亮, 充娜, 等. 杏鲍菇工厂化栽培优良菌株的筛选及亲缘关系评价[J]. 食用菌学报, 2010, 17(2): 19-21.
[7] 李玉, 于海龙, 周峰, 等. 工厂化瓶栽杏鲍菇疏蕾研究[J]. 上海农业学报, 2011, 27(1): 52-54.
[8] 刘宇, 陈文良, 王丽珍, 等. 杏鲍菇13号菌株选育研究[J]. 食用菌学报, 2004, 11(3): 61-64.
[9] 潘慧, 徐章逸, 边银丙. 灰色关联度法在杏鲍菇优良杂交菌株利筛选中的应用[J]. 食用菌学报, 2010, 17(2): 22-25.
[10] 王俊玲, 李明, 田景花, 等. 6个杏鲍菇菌株及其杂交子代的酯酶同工酶分析[J]. 河北农业大学学报, 2004, 27(3): 29-32.
[11] 尚晓冬, 宋春艳, 沈学香, 等. 杏鲍菇菌株RAPD指纹图谱分析[J]. 食用菌学报, 2009, 16(3): 1-4.
[12] 贺冬梅, 高君辉陈明杰, 等. 杏鲍菇菌株遗传差异的研究[J]. 食用菌学报, 1999, 6(4): 7-10.
[13] 冯伟林, 蔡伟明, 金群力, 等. ISSR分子标记分析杏鲍菇菌株遗传差异研究[J]. 中国食用菌, 2009, 28(1): 47-49.
[14] 陈强, 李翠新, 李辉平, 等. 真菌营养不亲和研究进展[J]. 食用菌学报, 2007, 14(1): 73-77.
[15] Smith ML, Gibbs CC, Milgroom MG. Heterokaryon incompatibility function of barrage-associated vegetative incompatibility genes () in[J]., 2006, 98(1): 43-50.
[16] 訾惠君, 刘连强, 周永斌, 等. 黄伞菌株拮抗实验与酯酶同工酶分析[J]. 天津农业科学, 2011, 17(5): 12-14.
[17] 王守现, 刘宇, 耿小丽, 等. 北京地区8个白金针菇主栽菌株的鉴别研究[J]. 华北农学报, 2009, 24(1): 219-222.
[18] 李冠喜,王珏, 邵世光, 等. 白灵菇品种的拮抗与品比鉴定[J]. 中国食用菌, 2006, 25(2): 17-18.
[19] 刘蕾, 宁丽, 郭立忠, 等. 12个真姬菇菌株拮抗实验及部分同工酶分析[J]. 食用菌, 2008, 1: 12-14.
[20] 郭倩, 潘迎捷, 周昌艳, 等. 姬松茸菌株种质资源多样性的初步研究[J]. 食用菌学报, 2004, 11(1): 12-16.
[21] 贾定洪, 王波, 郑林用, 等. 应用拮抗及ISSR方法鉴定香菇菌株[J]. 西南农业学报, 2013, 26(2): 832-834.
[22] 王磊, 宿红艳, 王仲礼, 等. 八种灵芝菌株的比较及筛选[J]. 食用菌学报, 2008, 15(4): 53-56.
[23] 林骥. 杏鲍菇、白灵菇、阿魏蘑标准菌株库的建立[D]. 福州: 福建农林大学硕士学位论文, 2010.
Study on classification ofstrains based on antagonism test
Liu Shengrong1, Zhang Weirui1, Ke Binrong2, Liu Xinrui2, Wu Xiaoping2
(1.Department of Biology, Ningde Normal University, Ningde Fujian 35200; 2.College of life science, Mycological Research Center, Fujian Agriculture and Forestry University, Fuzhou Fujian 35002)
Classification of 81 strains collected from different areas in China was carried out based on antagonism tests. The results showed that 81 tested strains could be divided into 11 different groups. Of these groups, the largest group consists of 25 strains. However, there are three groups each containing only one strain. The results showed that a high diversityoccurs instrains in China. Apparently synonymy may be a serious problems in: many of strains with different names may fall within the same group.
; antagonism test; classification; genetic diversity
S646
A
2095-0934(2015)01-33-04
吴小平,博士,福建农林大学生命科学学院教授,主要从事食用菌研究工作
猜你喜欢
煤炭与化工(2024年2期)2024-03-30 08:09:52
现代装饰(2022年2期)2022-05-23 13:15:40
今日农业(2020年23期)2020-12-31 09:00:42
今日农业(2020年22期)2020-12-25 02:30:40
现代装饰(2020年11期)2020-11-27 01:48:32
军事文摘(2020年20期)2020-11-16 00:32:12
今日农业(2020年24期)2020-03-17 08:58:04
中成药(2018年12期)2018-12-29 12:26:02
Asian Journal of Urology(2015年3期)2015-12-16 14:42:15
世界科学(2014年8期)2014-02-28 14:58:24
