
茄科主要经济作物青枯病抗性育种研究进展及综合防治
2015-11-26 20:26赵家昱尹延旭李宁王飞姚明华焦春海
湖北农业科学 2015年21期
赵家昱++尹延旭++李宁++王飞++姚明华++焦春海

摘要:综述了中国茄科(Solanaceae)主要栽培植物番茄、辣椒、茄子、马铃薯和烟草等青枯病病原菌(Ralstonia solanacearum)菌系的分类研究概况。总结了青枯病病原菌侵染过程、青枯病主要抗源材料的结构特征及茄科青枯病抗性遗传规律的研究进展。结合以上研究结果,提出了青枯病防治的主要途径。
关键词:茄科(Solanaceae);青枯病;抗病育种;综合防治
中图分类号:S641.3 文献标识码:A 文章编号:0439-8114(2015)21-5201-06
DOI:10.14088/j.cnki.issn0439-8114.2015.21.001
Research Advances in Bacterial Wilt Resistance Breeding and Integrated Control in Major Economic Crops of the Family Solanaceae
ZHAO Jia-yu1,YIN Yan-xu2,LI Ning2,WANG Fei2,YAO Ming-hua2,JIAO Chun-hai3
(1.Huanggang Normal College, Huanggang 438000,Hubei, China; 2. Cash Crops Institute, Hubei Academy of Agricultural Sciences, Wuhan 430064, China; 3. Hubei Academy of Agricultural Sciences, Wuhan 430064, China)
Abstract: The paper presented genetic diversity of Ralstonia solanacearum strains in major economic crops of the family Solanaceae like tomato, pepper, eggplant, potato and tobacco. It also summarized the research advances in the infection process of Ralstonia solanacearum, the structural differences between resistant and susceptible materials and the inheritance of resistance mechanism to bacterial wilt. According to the results, strategies of integrated control need to be used in the future.
Key words: Solanaceae;bacterial wilt;disease resistance breeding;integrated control
茄科(Solanaceae)是双子叶植物中的一大家族,在中国乃至世界上广泛栽培。青枯病是危害茄科植物的一种毁灭性的土传细菌性病害,病原为茄科劳尔氏菌(Ralstonia solanacearum)[1]。该病害发生时,植株茎叶萎蔫,最后整株材料枯死,青枯病在世界范围内分布[2]。该病危害大,造成损失重,在中国北至甘肃,南到海南都有发生[3]。该病害的抗性育种和防治已经成为茄科生产的研究热点。
青枯病病原菌可以侵染54科近450多种植物[3-5]。茄科植物中重要的经济作物番茄、辣椒、茄子、马铃薯和烟草等均受青枯病的危害[6],统计表明,茄子青枯病发生时,在美国可以造成茄子减产约50%,在印度造成减产75%以上,而在中国长江流域损失在20%~60%[7]。烟草青枯病发生时,烟叶大面积减产、品质变劣,造成严重的经济损失[8]。在保护地栽培中,青枯病每年在中国的江南、华南地区严重发生并造成番茄巨大的经济损失[9]。马铃薯青枯病是马铃薯生产上仅次于晚疫病的世界第二大病害。辣椒青枯病在中国辣椒栽培中发病率极高,一般年份发病在20%以上,严重时在50%以上,有的年份甚至造成绝收[10,11]。近年来,随着生产集约化水平的提高,青枯病发生更为严重。青枯病病原菌对不良环境的抵抗力强,分布广泛,繁殖速度快,常规方法难以有效防治[12,13]。青枯病的生理生化很复杂,迄今为止,尚无有效的化学防治药剂。该病害的严重性已经得到热带、亚热带和温带地区很多国家的重视[14]。
总之,对青枯病的研究及综合防治意义重大。本文对中国茄科主要栽培植物番茄、辣椒、茄子、马铃薯和烟草等青枯病病原菌菌系分类、青枯病病原菌侵染过程、青枯病主要抗源材料的结构特征、茄科青枯病抗性遗传规律等方面的研究进展综述如下,在此基础上提出了茄科主要经济作物青枯病防治主要途径。
1 侵染5个茄科作物的青枯病病原菌菌系分类
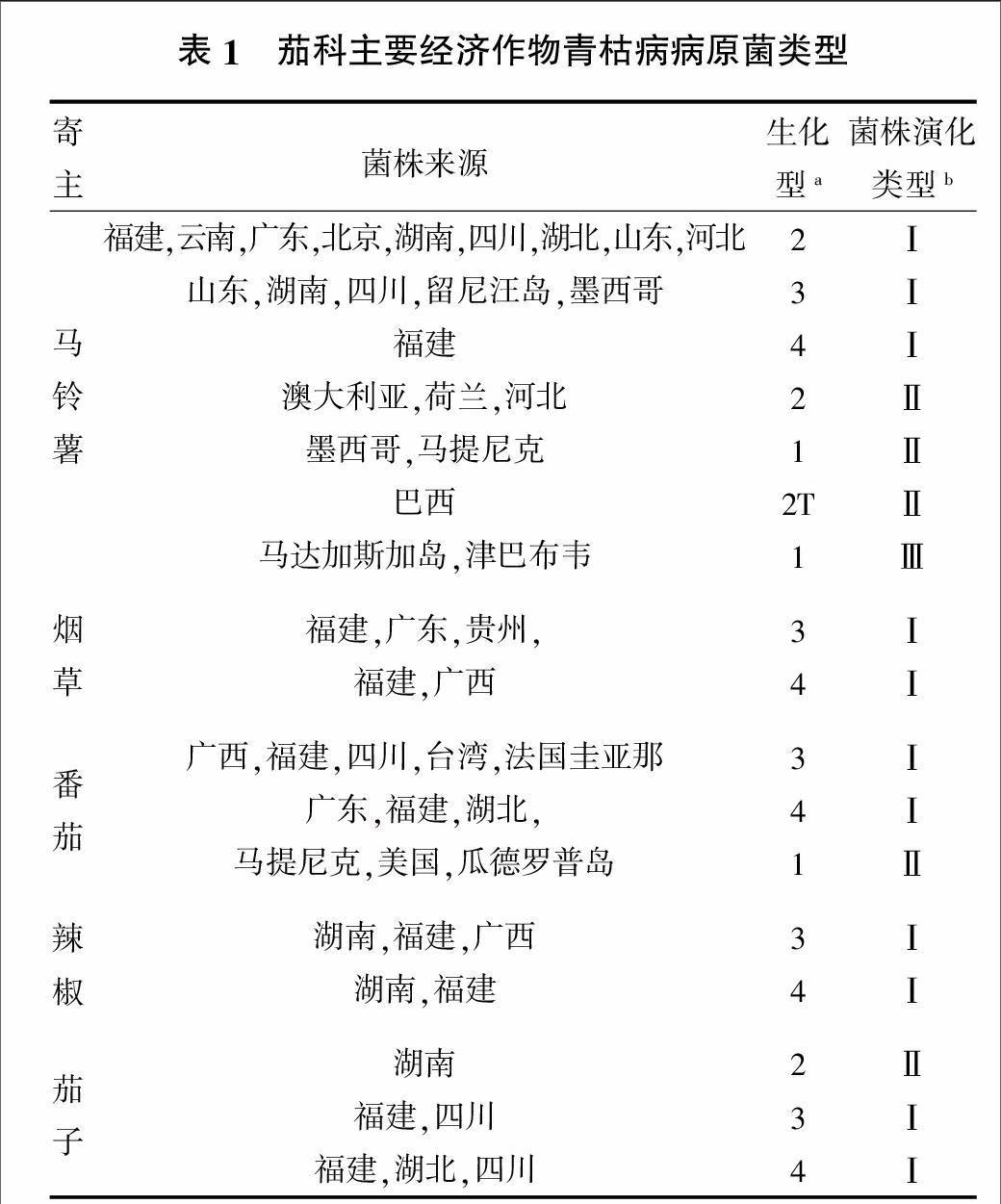
青枯病病原菌的生理生化类型比较复杂。传统的分类方法是依据寄主差异将青枯病病原菌分为5个不同的生理小种,也有依据菌株对麦芽糖、乳糖和纤维二糖及甘露醇、山梨醇和卫矛醇的氧化产酸能力差异分类,根据这一分类标准,将青枯病病原菌分为4个生化型[15]。目前,以分子生物学方法为依托的分类系统(利用限制性内切酶片段长度多态性和指纹识别系统等方法进行分析)能够更好地揭示青枯病病原菌的遗传多样性(表1),根据这一分类标准,将青枯病病原菌分为6个生化型(1、2、3、4、5和2T)[3]。通过多态性分析表明,侵染中国主要茄科植物的青枯病病原菌分类如下。
1)中国茄科的青枯病病原菌属于演化类型Ⅰ(PhylotypeⅠ)和Ⅱ(PhylotypeⅡ),以演化类型Ⅰ为优势菌群,该类群包括生化变种(Biovar)2、3和4。演化类群Ⅱ仅在河北省马铃薯栽培区和湖南省茄子栽培区中检测到。endprint
2)侵染中国马铃薯的主要青枯病病原菌以演化类型Ⅰ为主,其生化型为2、3、4,国外侵染演化类群为Ⅰ、Ⅱ、Ⅲ,生化型为1、2和2T。
3)侵染中国烟草的主要青枯病病原菌演化类型为Ⅰ,生化型为3和4。
4)侵染中国番茄的主要青枯病病原菌演化类型为Ⅰ,生化型为3和4,国外侵染演化类群为Ⅱ,生化型为1。
5)侵染中国辣椒的主要青枯病病原菌演化类型为Ⅰ,生化型为3和4。
6)侵染中国茄子的主要青枯病病原菌演化类型为Ⅰ(生化型为3和4)和Ⅱ(生化型为2)。
该分类结果表明,青枯病病原菌的群体比较复杂,区域之间的生理生化特征具有一定的规律性和可变性,这也反映了青枯病比较难以防治。在不同区域之间引种之后,对不同区域优势菌株及抗源抗性的再评价对于青枯病防治非常重要。
2 青枯病病原菌的侵染过程
青枯病病原菌侵染根系或者茎部可以分为有伤侵染和无伤侵染两种类型。研究表明,青枯病病原菌一般从植物根部或者颈部伤口处侵入。以番茄萌发3 d后的幼根为对象研究青枯病病原菌无伤侵染模式,利用扫描电镜分析发现病原菌侵染路线:根冠或根毛区、相邻细胞间隙→幼根细胞壁。该侵染路线中,病原菌优先通过幼根的根冠或者根毛区吸附并繁殖,也可以通过幼根表面相邻细胞间的病态间隙侵入,进而分解幼根细胞壁,从而入侵[17]。通过大田烟草青枯病病原菌的侵染动态分析,移栽烟草第5周根部样品首先检测出青枯病病原菌,此时个别烟草根部出现黑色坏死症状;移栽第7周根部样品检出阳性率达到50%左右,根部有黑色坏死,茎部还没有明显症状,发病率开始快速上升;移栽第9周,个别烟草茎部出现黑色坏死症状;移栽第12周,茎部检出率、根茎发病率均在80%以上[18]。
3 青枯病主要抗源材料特征及抗性遗传规律
3.1 青枯病病原菌寄主抗源分析
研究发现,茄科植物没有对青枯病免疫的材料[19]。对不同茄科植物中的主要抗源分述如下。
3.1.1 马铃薯青枯病抗源 已有的研究表明,马铃薯野生种和栽培种有200多个,用于生产的栽培种仅有8个[20]。马铃薯二倍体野生种、原始栽培种如Solanum acaule、S.chacoense、四倍体野生种S.stnotomum、四倍体栽培种资源对青枯病具有不同程度的抗性或耐性[21]。French等[22]研究表明,S. acaule、 S.boliviense、 S.bukasovii、S. candolleanum、S.coelestipetalum、 S.leptophytes、 S.peloquinianum、S.phureja、S.raphanifolium、S.sparsipilum、S.sogarandinum和S.tapojense,这些马铃薯野生种内含有高抗青枯病的抗源。
3.1.2 烟草青枯病抗源 匡传富等[23]通过室内接种青枯病病原菌鉴定发现,抗病品种有TI448A、贵烟、CV 91、大晒烟、香烟、K326等6个,中抗品种有大幅、D101、G28、安流晒烟、子州羊角大烟、G28× MD609、小样无烟、清间羊角大烟、N326等9个品种。诸多研究表明,TI448A是一个抗青枯病材料[24-26]。也有研究表明,Pinkney Arthur、DSPA、Davis Special、400、南美引进TIS系列品种、Oxford207、Xanthi、kokubu、D101,G80、 Enshu、 Hatanodaruma和79-X等材料能够抗青枯病侵染[24-31]。
3.1.3 番茄青枯病抗源 培育抗青枯病材料的抗源,国内主要从国外引进。其中亚洲蔬菜研究与发展中心(Asian Vegetable Research and Development Center,AVRDC)引进的抗源有R-3034-10-N-UG、Cravel、CLN1463-160-40-60、CLN1464-111-30-45 和Redlander,这些材料发病少、产量、品质较好[32],可以在育种上进一步利用。在中国南方科研院所选育的抗青枯病品种有0718-3、0769-1、杂交1号、杂交3号、丰顺、大丰顺、好时年、福安、多宝、夏红、抗青1号、抗青19号、粤红玉、粤宝、粤星、新星、东方红1号、年丰、穗丰、益丰和益丰2号[32,33]。辽宁省农科院T2-08-555材料对青枯病表现为抗性[34]。国际上番茄青枯病高度抗病自交系有LS89、Lycopersicon pimpinellifolium PI127805A、Lycopersicon esculentum var.cerasiforme CRA66、Lycopersicon pimpinellifolium PI129080、Beltsville#3814、Lycopersicon esculentum AS52、Hawaii7996[35-38]。
3.1.4 辣椒青枯病抗源 目前已知国内辣椒材料的抗源或品种有宁优6-2-4、31-2-a、79-5×18-1、SE、79-5×18-1、52-5-2纯、39-2-a-1、136-3、65-5-B、76a、宁优6-1×18-1、64-5纯牛、83B-2×18-1、55-2-2-2[39];红塔单身朝天椒[10];湘研1号、早杂2号、通椒1号、粤椒1号、新椒1号[40]。国际上辣椒青枯病抗源有KA2、PM687、Anugraha,商品种有:Smatra、Hot Beauty、Delicacy、Avon Eg、Ever Flavor、Akashi、Fushimi-Amanga、Mie-Midori Commercial hybrids Kyo-Nami、Shin-Sakigake 2、Baramashi、LS1439、Tachi-Yatsubusa、Weonkyo 306;Capsicum. frutescens栽培种:Heiser 6240和LS 2390;C. baccatum栽培种:BGH 1761、LS1716和Casali;PBC385、PBC066、PBC204、PBC1347、CNPH143(MCendprint
4)、CNPH144(MC5)和CNPH145(HC10)[41]。
3.1.5 茄子青枯病抗源 目前已报道的茄子抗性资源主要有:来自马来西亚的TS3、S47A;印度尼西亚的S56B(Terong Hi jan)、TS69(Gelatik)和TS90;印度的EG192(Arkashirish)、EG193(ArkaNidhi)和 EG195(BB49)[42];S69[43];抗性材料WCGR112-8、抗性材料LS1934[44];MM960[6]等。
不同茄科经济作物中的主要抗源及其文献来源如表2所示。
3.2 青枯病抗源材料的结构特征
通过对番茄材料的显微观察表明,青枯病病原菌在番茄抗病试验材料与感病试验材料植株根内的扩展差异显著;青枯病病原菌可存在于感病试验材料皮层薄壁细胞间隙,瓦解细胞壁,形成溶生腔。在抗病试验材料维管组织中青枯病病原菌则被细胞壁所吸附;抗病试验材料的导管分子比感病试验材料短[51]。
3.3 抗青枯病育种现状
3.3.1 马铃薯青枯病抗性遗传规律 马铃薯青枯病抗性的遗传机制较为复杂,目前研究分析尚未得到一致的结论。
3.3.2 烟草青枯病抗性遗传规律 孙学永等[52]对15份烟草青枯病抗性程度不同的烟草种质研究表明烟草青枯病遗传模型为加性-显性-上位模型;遗传效应以加性效应为主,加性×加性上位性作用效应明显,加性及其上位性与环境有互作效应,显性效应较小或不显著,抗性狭义遗传力为30.40%~57.14%。范江等[30]以抗青枯病的烟草品种“Oxford207”和感病品种“红花大金元”为亲本,构建了 F1、F2和 BC1群体,研究结果表明“Oxford207”的青枯病抗性不符合典型的显性或隐性基因控制模型,属于加性基因控制。杨友才等[25]选用高感青枯病品种红花大金元与高抗青枯病品种Ti448A配制杂交组合,人工注射法接种鉴定两亲本及其杂交后代 F1、F2、BC1P1代,结果表明抗病亲本材料Ti448A的抗病性由一对显性基因控制。高加明等[24]以香料烟抗青枯病品种Xanthi、感病品种Samsun、F1以及F2群体为研究材料,采用青枯病圃进行抗性鉴定,香料烟青枯病抗性基因是受2对加性-显性-上位主基因+加性-显性多基因(E-1模型)控制;主基因的遗传率为49.63%。
3.3.3 番茄青枯病抗性遗传规律 李海涛等[47]以感病材料98TL-1(P1)、L-402母本自交系(P3)、高抗青枯病自交系LS89(P2)及抗、感材料配制杂交组合和回交世代为试材,发现抗病基因对感病基因表现为部分显性。番茄抗青枯病自交系的遗传是一个基因起主导作用,该基因受核基因控制,质基因对其影响不大。利用不同的番茄青枯病抗性资源分析,其抗性遗传有隐性、不完全显性和显性控制。控制抗性基因的数目除上述主基因外,还有寡基因或多基因控制,基因间的作用有显性、上位和加性效应[35]。
3.3.4 辣椒青枯病抗性遗传规律 Lafortune等[53] 对热带地区的抗性材料PM687和感病材料YoLo wonder的杂交后代双单倍体进行分析发现寄主对该病抗性为寡基因控制,抗性稳定遗传。Hwang等[49]通过对抗感青枯病材料的辣椒“KC305”和“Chilbok”接种青枯病病原菌后1、3、6 d的转录组数据库分析,筛选到115个抗性基因(R)和109个感病基因(S),其中R基因表现上调,S基因表现下调,R基因主要参与木葡聚糖合成和细胞壁的构建,而S基因主要响应胁迫反应和细胞死亡。
3.3.5 茄子青枯病抗性遗传规律 茄子青枯病抗性遗传规律研究开展以来,发现茄子对青枯病的抗性遗传符合“加性-显性”效应模型,遗传效应中同时有加性-显性-反交效应,但以加性效应为主[54]。Lebeau等[6]对抗病茄子材料AG91-25(MM960)的遗传分析表明,其对青枯病病原菌生化型I的抗性为数量遗传,受至少1个显性主效基因控制。应用MM738×AG91-25的F6 RIL群体对抗性相关基因进行定位,得到一个主效QTL ERs1,位于LG2,与标记COX067a紧密连锁,遗传距离小于1 cM;QTL间存在互作效应,LG13上的QTL的缺失与否影响其对青枯病病原菌菌株PSS4的抗性。李猛等[43]以茄子高抗S69材料和感病品种三月茄为试材,结果显示茄子对青枯病病原菌的抗病性属于不完全隐性遗传,感病属于不完全显性遗传,抗性由1~2对基因控制,抗性基因连锁的AFLP标记39A980(引物组合为E-ACC/M-CTC),该标记与抗性基因间的遗传距离为4.9 cM。
4 青枯病防治的主要途径
4.1 选用抗病品种
青枯病严重危害茄科植物,病害发生快,造成大量植株死亡,产量降低。近年来提倡减肥减药的健康栽培模式,因此,选育抗病品种是解决该病害的关键。因该病害在茄科中抗病遗传多受到隐性和多基因或寡基因控制,育种的难度增加、育种速度缓慢。通过引入一部分抗源和筛选一部分野生抗源,逐步育成较强的青枯病抗性材料,进而解决青枯病抗性育种瓶颈。茄科抗病资源对于选育抗病品种意义重大,马铃薯抗源中虽然种间利用受到倍性水平和胚乳平衡数的差异限制,但是现代生物技术育种在不断解决常规育种中出现的生物学障碍,为常规育种工作快速发展奠定了基础。
4.2 生物防治
青枯病的生物防治措施,在国内外已经引起高度的重视。生物防治主要是利用无致病力的青枯病病原菌株产生细菌素,通过该物质对同种细菌的其他菌株或者近缘菌株产生杀伤等作用。目前在防治效果上,正在由防效偏低、持效性短及不稳定向成熟大田试验方向转化。
4.3 嫁接防治
在茄科育种中,受到生殖隔离和远缘杂交亲和性的影响,限制不同种属野生资源的利用,通过茄科不同种属间的嫁接措施,充分利用自然抗源材料来提高番茄等茄科经济作物的抗青枯病能力。endprint
4.4 加强田间管理,合理轮作
青枯病综合防治主要包括合理选地、培育壮苗、合理轮作、搞好田间卫生等,发现田间病株,应该立即拔除病株,并撒少量石灰消毒。减少大水漫灌,早育苗、早栽培、早收获。条件合适的地方,尽可能开展与十字花科轮作4~6年,减少病害的流行。目前,部分地区结合功能有机肥与功能基质的综合防控措施,在青枯病防治上有一定的效果。
4.5 化学防治
目前还没有高效、低毒、无污染的化学杀菌剂应用于生产。不同化学药剂(如铜制剂和芽孢杆菌)的综合使用,能够降低青枯病的发病率。
通过解析茄科植物的抗青枯病机制,争取在茄科主要经济作物青枯病抗性改良上有所突破,结合以上措施降低青枯病发生造成的损害。
参考文献:
[1] MORI T, OZAKI K, MATSUSAKI H, et al. Behavior and mutation of Ralstonia solanacearum in Solanum toxicarium grown in aseptic culture[J]. Journal of General Plant Pathology, 2007,73(2):142-146.
[2] BARRETTIP B, DE SOUZA R M, POZZA E A, et al. Combination of endophytic bacteria and resistant cultivars improves control of Ralstonia wilt of tomato[J]. Australasian Plant Pathology, 2012,41(2):189-195.
[3] XU J, PAN Z,PRIOR P,et al.Genetic diversity of Ralstonia solanacearum strains from China[J]. European Journal of Plant Pathology,2009,125(4):641-653.
[4] 卢 同.我国作物细菌性青枯菌的研究进展[J].福建农业学报,1998,13(2):33-40.
[5] 黄真池,刘 媛,曾富华,等.青枯菌致病机理及作物抗青枯病研究进展[J].热带亚热带植物学报,2008,16(5):491-496.
[6] LEBEAU A,GOUY M, DAUNAY M,et al.Genetic mapping of a major dominant gene for resistance to Ralstonia solanacearum in eggplant[J]. Theoretical and Applied Genetics,2013,126(1):143-158.
[7] 朱育菁,刘 波,林抗美,等.ANTI-8098A生物杀菌制剂在连作重病区对茄子青枯病的防效测定[J].中国植保导刊,2005, 24(11):8-11.
[8] 兰俊荣,刘启彤,何宏仪.部分烟草种质资源的青枯病抗性鉴定[J].福建农业科技,2010(5):62-63.
[9] 周岗泉,张秀冬,刘琼光,等.抗感青枯病番茄的内生细菌数量动态分析及其对青枯病的生物防治[J].微生物学通报,2007, 588(34):885-888.
[10] 谭清群,袁 洁,杨学辉,等.贵州省辣椒新品种对疫病和青枯病的抗性鉴定研究[J].种子,2014,33(11):82-85.
[11] CHEN Y J,LIN Y S,CHUNG W H. Bacterial wilt of sweet potato caused by Ralstonia solanacearum in Taiwan [J].Journal of General Plant Pathology, 2012, 78(1):80-84.
[12] 魏小伞,曹必好,雷建军,等.茄子抗病育种研究进展[J].中国蔬菜,2010(10):1-8.
[13] 俞 琼,徐根娣,刘 鹏.茄子抗病育种研究进展[J].安徽农业科学,2006,34(6):1069-1071.
[14] HORITA M,TSUCHIYA K,SUGA Y,et al.Current classification of Ralstonia solanacearum and genetic diversity of the strains in Japan[J]. Journal of General Plant Pathology, 2014, 80(6):455-465.
[15] 杨玉红.茄科植物青枯菌病害研究进展[J].江西农业学报,2008,20(5):54-55.
[16] 王胜坤,王 军,徐大平.植物青枯菌检测方法研究进展[J].南京林业大学学报(自然科学版),2007,31(2):118-122.
[17] 王 丽,汪 矛.青枯菌侵染番茄幼根的扫描电镜观察[J].农业生物技术学报,1999,7(2):157-162.
[18] 刘 勇,于海芹,秦西云,等.云南省烟草青枯病的侵染动态[J].中国农学通报,2007,23(11):207-210.
[19] PATIL V U, GOPAL J, SINGH B, Improvement for bacterial wilt resistance in potato by conventional and biotechnological approaches [J]. Agricultural Research, 2012, 1(4):299-316.endprint
[20] 田伶俐.马铃薯体细胞杂种胞质遗传组成及其与青枯病抗性的关系[D].武汉:华中农业大学,2010.
[21] KIM-LEE H, MOON J, HONG Y, et al. Bacterial wilt resistance in the progenies of the fusion hybrids between haploid of potato and solanum commersonii[J]. American Journal of Potato Research,2005,82(2):129-137.
[22] FRENCH E R, ANGUIZ R, ALEY P.The usefulness of potato resistance to Ralstonia solanacearum, for the integrated control of bacterial wilt[A]. PRIOR P, ALLEN C, ELPHINSTONE J. Bacterial Wilt Disease[M]. Germany: Springer Berlin Heidelberg,1998.381-385.
[23] 匡传富,罗 宽.烟草品种对青枯病抗病性及抗性机制的研究[J].湖南农业大学学报(自然科学版),2002,28(5):395-398.
[24] 高加明,王志德,张兴伟,等.香料烟青枯病抗性基因的遗传分析[J].中国烟草科学,2010,31(1):1-4.
[25] 杨友才,周清明,朱列书.烟草青枯病抗性基因的遗传分析及 RAPD标记[J].中国烟草学报,2006,12(2):38-42.
[26] JACK A.The CORESTA collaborative study on bacterialwilt (Ralstonia solanacearum)-2002 report[J]. CORESTA Information Bulletin, 2002,4:45-58.
[27] 钱益亮.烟草青枯病抗性遗传分析及QTL定位研究[D].合肥:安徽农业大学,2013.
[28] KIM S,OLSON T,SCHAAD N,et al. Ralstonia solanacearum race 3,biovar 2,the causal agent of brown rot of potato,identified in geraniums in Pennsylvania, Delaware,and Connecticut[J].Plant Disease,2003,87(4):450.
[29] SMITH T,CLAYTON E,Inheritance of resistance to bacterial wilt in tobacco[J]. Journal of Agricultural Research,1948,76(1):27-32.
[30] 范 江,刘 勇,童治军,等.烤烟品种‘Oxford207青枯病抗性的遗传分析与分子标记初选[J].中国农学通报,2013,29(34):50-55.
[31] 吴 超,夏岩石,吕永华,等.烟草青枯病抗性分子标记的研究进展[J].分子植物育种,2015,13(4):937-945.
[32] 汪国平,袁四清,熊正葵,等.广东省番茄青枯病相关研究概况[J].广东农业科学,2003(3):32-34.
[33] 丘漫宇,张素平,刘玉平,等.抗青枯病番茄新品种‘益丰2号[J].园艺学报,2013,40(11):2321-2322.
[34] 邹庆道,朱 华,张子君,等.番茄抗青枯病材料的筛选[J].北方园艺,2011(2):28-29.
[35] 汪国平,林明宝,吴定华.番茄青枯病抗性遗传研究进展[J].园艺学报,2004,31(3):403-407.
[36] 苗立祥.番茄抗青枯病的AFLP分子标记及其相关基因的克隆[D].杭州:浙江大学,2008.
[37] WANG J F, HO F I,TRUONG H T H,et al. Identification of major QTLs associated with stable resistance of tomato cultivar ‘Hawaii 7996 to Ralstonia solanacearum[J]. Euphytica, 2013,190(2):241-252.
[38] NAKAHO K,INOUE H,TAKAYAMA T,et al. Distribution and multiplication of Ralstonia solanacearum in tomato plants with resistance derived from different origins[J]. Journal of General Plant Pathology,2004,70(2):115-119.
[39] 党峰峰,雷玉芬,官德义,等.辣椒种质资源抗青枯病的鉴定与评价[J].植物科学学报,2013,31(4):378-384.
[40] 毛艳梅.辣椒青枯病发病规律与综合防治[J].农机服务,2010,27(11):1424-1424,1426.
[41] BABU B S,PANDRAVADA S,RAO R P,et al.,Global sources of pepper genetic resources against arthropods, nematodes and pathogens[J]. Crop Protection,2011,30(4):389-400.endprint
[42] 封林林,屈冬玉.茄子青枯病抗性材料的鉴定及性状观察[J].长江蔬菜,2000(10):35-37.
[43] 李 猛,王永清,田时炳,等.茄子青枯病抗性基因的遗传分析及其AFLP标记[J].园艺学报,2006,33(4):869-872.
[44] 李海涛,邹庆道.茄子对青枯病的抗性遗传研究Ⅱ.茄子抗病材料LS1934的遗传分析[J].辽宁农业科学,2002(3):1-3.
[45] 何礼远.抗青枯病高产优质马铃薯新品种“抗青9-1”[J].中国马铃薯,2007,21(6):381-382.
[46] GAO G,QU D,LIAN Y,et al. A RAPD marker linked to the potato resistance to bacterial wilt[J]. Acta Phytopathologica Sinica,2001,32(3):267-271.
[47] 李海涛,邹庆道.番茄对青枯病的抗性遗传研究[J].辽宁农业科学,2001(5):1-4.
[48] 郑锦荣,张衍荣.番茄抗青枯病材料的鉴定与筛选[J].中国蔬菜,1998(6):8-10.
[49] HWANG J,YOUNGMI C,JUMSOON K,et al. Microarray analysis of the transcriptome for bacterial wilt resistance in pepper (Capsicum annuum L.) [J]. Notulae Botanicae Horti Agrobotanici Cluj-Napoca,2011,39(2):49-57.
[50] 杨建国,皮向红,陈惠明,等.茄子ER300抗青枯病遗传及在育种中的应用[J].湖南农业大学学报(自然科学版),2006,32(3):277-279.
[51] 乐素菊,梁承愈,吴定华.番茄青枯病抗、感品种(系)结构性差异初探[J].华南农业大学学报,1996,17(2):50-53.
[52] 孙学永,王新胜,张丽娜,等.烟草青枯病抗性的遗传分析[J].中国农学通报,2013,29(34):56-60.
[53] LAFORTUNE D,B?魪RAMIS M,DAUB?魬ZE A M,et al. Partial resistance of pepper to bacterial wilt is oligogenic and stable under tropical conditions[J]. Plant Disease,2005,89(5):501-506.
[54] 封林林,屈冬玉,金黎平,等.茄子青枯病抗性的遗传分析[J].园艺学报,2003,30(2):163-166.endprint
猜你喜欢
广东农业科学(2017年5期)2017-08-29
少年科学(2016年6期)2017-02-08
吉林农业(2016年12期)2017-01-06
山东工业技术(2016年23期)2016-12-23
安徽理工大学学报·自然科学版(2016年2期)2016-12-22
现代农业(2015年1期)2015-02-28
植物营养与肥料学报(2012年5期)2012-10-26
中国烟草学报(2012年5期)2012-04-12
中国烟草学报(2012年6期)2012-04-09
