
在无杀虫剂选择下家蝇拟除虫菊酯抗性等位基因频率的变化*
2015-11-12 00:32:38周云辉李保同邱星辉
寄生虫与医学昆虫学报 2015年3期
潘 婧 周云辉 李 梅 李保同 邱星辉**
(1.中国科学院动物研究所农业虫害鼠害综合治理研究国家重点实验室,北京100101;2.中国科学院大学, 北京100049;3.江西农业大学,南昌 330045)
家蝇Muscadomestica是一种重要的卫生害虫,可以传播100多种人和动物疾病(Greenber, 1965)。 家蝇的骚扰还可以降低禽类的产蛋量和家畜的饲料转化率 (Scottetal., 2009)。控制家蝇主要依赖化学杀虫剂,各种类型的杀虫剂如传统的有机磷、氨基甲酸酯、拟除虫菊酯以及相对新的吡虫啉等都被用于家蝇的化学防治。拟除虫菊酯类杀虫剂因其具有低毒和高效等特点,被广泛使用,由此带来了家蝇对其抗性的产生和扩散,家蝇抗药性的事例有很多的报道 (Wangetal., 2012; Gaoetal., 2012; Scottetal., 2013)。
家蝇对拟除虫菊酯的抗性已知主要有两类机制,即靶标(钠离子通道)不敏感性和细胞色素P450介导的代谢解毒作用增强的代谢抗性(Scottetal., 2004)。至今已鉴定出3种钠离子通道抗性等位基因,包括kdr(1014F, F) (Williamsonetal., 1996),kdr-his(1014H, H) (Liuetal., 2002) andsuper-kdr(918T + 1014F) (Williamsonetal., 1996, Qiuetal., 2007; Rinkevichetal., 2007)。也发现一个或多个细胞色素P450的过量表达与家蝇对拟除虫菊酯类杀虫剂抗性相关联 (Scottetal., 2004; Gaoetal., 2012),比如在美国的LPR和中国及美国的田间抗性家蝇种群存在CYP6D1v1 抗性基因 (Kasaietal., 2000; Seifertetal., 2002;Rinkevichetal., 2006; Gaoetal., 2012)。
通常认为,抗药性是适应杀虫剂选择的一种进化现象,在无该杀虫剂的环境下抗药性昆虫种群通常具有一定的适合度代价(Kliotetal., 2012)。尽管这一假设得到了不少事例的支持,但也有研究表明抗性昆虫并不表现适合度不利 ( McCartetal., 2005; Crowderetal., 2009; Basitetal., 2012)。这些前期的研究表明,与抗性相关联的适合度可能因抗性机制或抗性种群的不同而不同。有关家蝇钠离子通道突变以及CYP6D1v1 相关的抗药性是否存在适合度代价还不清楚。本研究以从北京采集的家蝇为研究系统,通过在实验室不接触杀虫剂的条件下,连续饲养和监测其抗性等位基因频率的变化,依此推测抗性等位基因是否存在适合度代价。
1 材料与方法
1.1 家蝇种群
在2010年10月,于北京奥林匹克公园的垃圾筒用网捕的方法采集100多头家蝇(亲代),带回实验室建立种群(取名为BJ2010),并以500~1 000头的种群规模连续传代。将每代产卵后的成虫置于100% 乙醇中,保存在 -20℃ 用于个体基因型分析。对BJ2010品系家蝇的毒力测定结果显示,相对于实验室敏感品系,BJ2010对氯菊酯和溴氰菊酯的抗性倍数分别为25和167倍。
1.2 钠离子通道基因和CYP6D1基因的基因分型
在保存的亲代(P)、子1代(F1)、子6代(F6)、子10代(F10)、子15代(F15)、子20代(F20)、子25代(F25)成虫样品中,分别随机抽取30头以上个体的头胸部,单头提取其基因组DNA(Rinkevichetal.,2006)。 CYP6D1的基因型采用PCR-RFLP 的方法(Rinkevichetal.,2006)确定。 钠离子通道基因型亲代样品采用测序的方法(Rinkevichetal.,2006),其他代样品采用PCR-RFLP 方法确定(Qiuetal., 2012 )。
2 结果
2.1 BJ2010亲代样品拟除虫菊酯杀虫剂抗性等位基因种类与频率
对田间采集的家蝇样品中,检测到了细胞色素 P450 CYP6D1基因的抗性和敏感等位基因。 抗性等位基因(CYP6D1v1, R)的5’侧区包含15 bp的插入片段,其频率为0.06 (图1), 不包含该特征15 bp的插入片段的CYP6D1基因为敏感等位基因(S),频率高达0.94。 三种可能的基因型(SS, SR, RR)在样品中均存在,敏感纯合、杂合子和抗性纯合子的频率分别是0.91、 0.07 和0.02(图2)。

图1 家蝇种群CYP6D1等位基因频率变化Fig.1 Changes in the frequencies of CYP6D1 alleles over 25 generations under laboratory conditions

图2 家蝇种群CYP6D1基因型频率变化Fig.2 Changes in the frequencies of CYP6D1 genotypes over 25 generations under laboratory conditions
从样品中鉴定了3个钠离子通道等位基因,包括2个抗性等位基因kdr和kdr-his和1个敏感等位基因(1014L,L),未检测到文献报道的super-kdr,其中,敏感等位基因频率最高(0.81),kdr抗性等位基因频率最低(0.07) (图3)。 同样,所有的6种基因型个体(LL, FF, HH, FL, HL, HF)在种群中都存在(图4),其中最常见的基因型是敏感纯合子(0.69), 其次是HL 杂合子(0.17) 和FL 杂合子(0.06), 另外三种基因型(HH, FF, HF) 的家蝇个体很罕见(频率在0.02~0.03之间)。
2.2 无杀虫剂选择下的家蝇种群对拟除虫菊酯杀虫剂抗性等位基因频率的变化
在无杀虫剂条件下,25代内的观察,CYP6D1 抗性等位基因频率在0.02 至 0.13之间变动, 敏感等位基因频率变化于0.87 和 0.97 之间(图1),CYP6D1不同基因型个体频率的变化也不显著,敏感纯合个体、杂合个体以及抗性纯合个体的变化范围分别是0.80~0.96、 0.04~0.17 和 0~0.05 (图2)。
在实验室正常繁殖25代内,家蝇种群钠离子通道基因的敏感等位基因1014L 频率在各代中保持最高,1014H 等位基因则在0.09 和0.26之间波动, 而1014F 在波动中从起始的0.07 降到0.01(图3)。 总体上看,敏感纯合个体 (LL) 保持着高频率 (0.56~0.82), HL 杂合基因型个体频率在0.17 与 0.37之间,但抗性基因型 (FF纯合, FH 杂合和HH纯合) 个体频率一直处于低水平 (低于0.1), FF纯合、FH 杂合和HH纯合个体分别自F1、 F15 和 F20代之后检测不到(图4)。
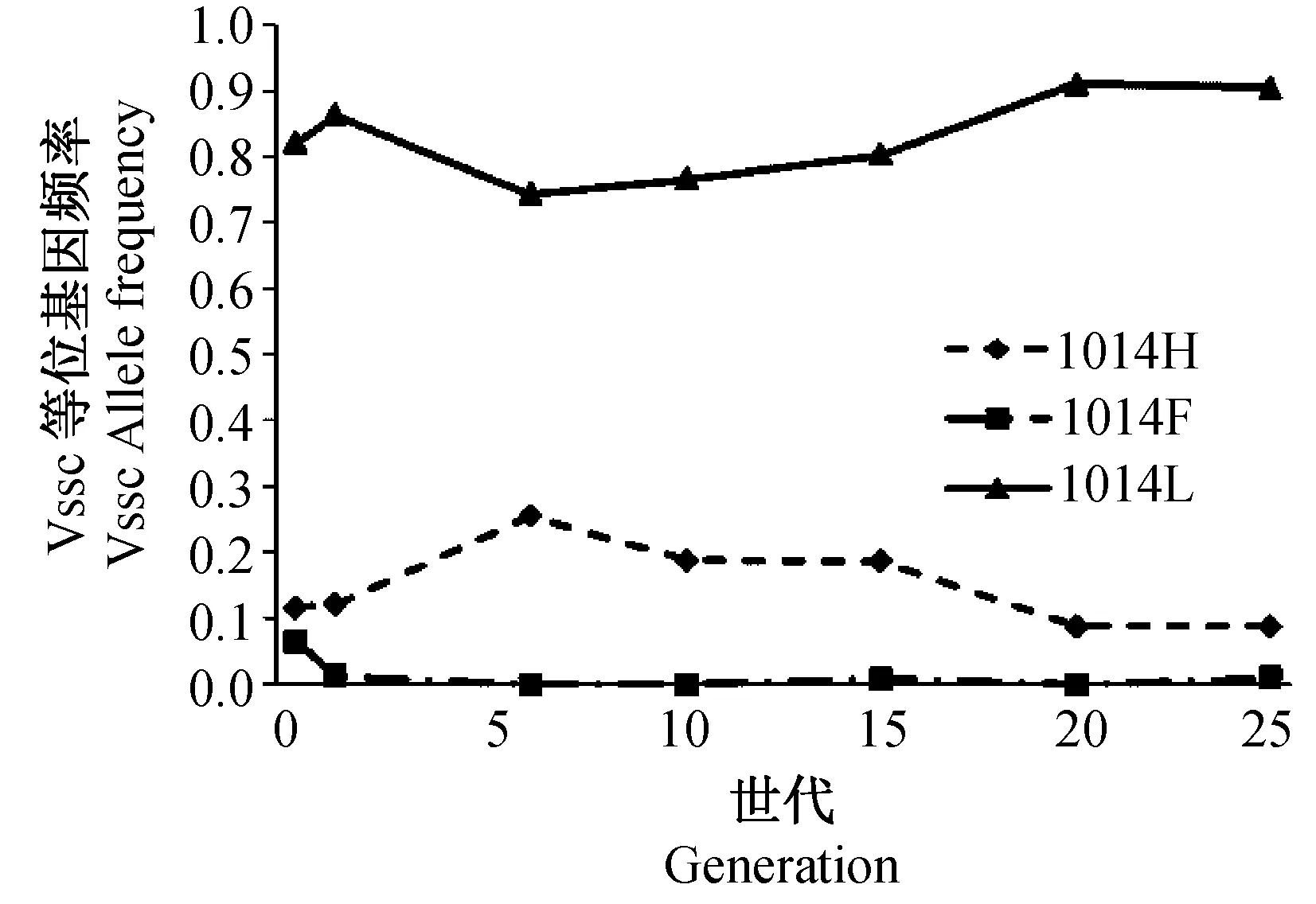
图3 家蝇种群钠离子通道等位基因频率变化Fig.3 Changes in the frequencies of Vssc alleles over 25 generations under laboratory conditions

图4 家蝇种群钠离子通道基因型频率变化Fig.4 Changes in the frequencies of six Vssc genotypes over 25 generations under laboratory conditions
3 讨论
家蝇对拟除虫菊酯的抗药性受多基因的控制,可以通过基因的位点突变(如钠离子通道基因Vssc),或更复杂的分子事情如基因的表达调控 (如CYP6D1) 来实现。 拟除虫菊酯杀虫剂用于公共卫生领域始于上世纪70年代末,至今有30多年的用药历史,家蝇产生抗药性并不令人奇怪。通过对 98 个家蝇个体的基因型检测, 发现了kdr、kdr-his和CYP6D1v1 抗性等位基因的存在。kdr-his的基因频率(0.12) 高于kdr(0.07),这与我们先前对中国采集的家蝇的研究结果一致(Wangetal., 2012)。值得注意的是,无论是钠离子通道基因,还是CYP6D1基因,敏感等位基因频率在本研究的种群中的频率很高。如敏感的钠离子通道基因(1014L)的频率在0.81, 高于从美国和丹麦采集的家蝇(Rinkevichetal., 2006; Huangetal., 2004; Scottetal., 2013), 但接近土尔其家蝇种群(0.72) (Taskinetal., 2011)。北京种群CYP6D1 敏感等位基因(S)频率高达0.94, 比报道的采自美国、土尔其等地的家蝇种群高很多(Rinkevichetal., 2006; Rinkevichetal., 2007; Taskinetal., 2011; Scottetal., 2013), 但在我国家蝇种群已报道的频率值范围内(Wangetal., 2012)。BJ2010种群具有对拟除虫菊酯的抗性,但其钠离子通道抗性等位基因以及CYP6D1v1抗性等位基因的的频率都很低,暗示该种群还存在其他抗性机制。
通常认为,抗药性是昆虫适应杀虫剂选择下的结果,由此推测抗性种群相对于敏感种群在没有杀虫剂的环境下具有更低的适合度 (Roushetal., 1990)。 如果该推测正确,从抗性遗传的角度则抗性等位基因频率在无杀虫剂的条件下会随时间而下降。本研究的结果显示CYP6D1 的抗性等位基因频率在25代传代范围内变化非常之小,提示CYP6D1v1 并不表现出适合度代价,这一结果与美国的一个研究结果相一致(Rinkevichetal., 2013),也部分解释了CYP6D1v1 在全球广泛存在的原因 (Rinkevichetal., 2006; Taskinetal., 2011; Wangetal., 2012)。与此类似,钠离子通道1014H(kdr-his)抗性等位基因频率变化在25代期间的变化也很小,表明在无杀虫剂的实验室条件下,相对种群中存在的其他等位基因,钠离子通道1014H等位基因也不存在适合度代价。
早期的一个研究表明携带1014F 突变的抗性家蝇与敏感家蝇相比表现出温度偏好性的差异的(Fosteretal., 2003),这种差异是否代表具有适合度代价现在还不清楚。 通过对种群中1014F 频率随时间的动态变化发现,1014F 频率有所下降(从0.07到0.01),但由于1014F 的起始频率非常低,其频率的下降很有可能是遗传漂变的结果。而不是其存在适合度劣势,这一推论还需要进一步的研究数据。
综上,通过对家蝇已知的抗性等位基因频率在种群中的频率的长期监测,发现在无杀虫剂的条件下拟除虫菊酯抗性等位基因频率并不下降,说明携带抗性等位基因kdr-his或CYP6D1v1的家蝇不表现适合度不利。抗性等位基因频率的动态变化结果表明,抗性基因一旦在种群被选择出来,在无杀虫剂的选择下达到平衡,很难再从群体中消失。这一研究结果与王学军等(2008)根据毒力测定数据而得出的结论相一致。这也提示,换用杀虫剂对抗性家蝇的防治可能有效,而轮用以往常用的杀虫剂则存在风险。
猜你喜欢
江西农业学报(2022年8期)2022-11-04 07:37:02
乡村科技(2022年2期)2022-03-25 14:56:16
仪器仪表用户(2021年10期)2021-11-27 08:26:14
奇闻怪事(2020年7期)2020-09-10 01:41:40
山东第一医科大学(山东省医学科学院)学报(2019年11期)2019-11-07 02:01:12
科学种养(2016年4期)2016-04-19 03:47:43
中华老年多器官疾病杂志(2016年2期)2016-01-16 03:15:46
电源技术(2015年2期)2015-08-22 11:28:30
物理化学学报(2015年5期)2015-02-28 17:34:57
湖北科技学院学报(医学版)(2014年3期)2014-02-28 19:43:05
