
日本囊对虾G1群体生长性状遗传参数估计
2015-11-10 07:23:06钟声平苏永全刘洪涛
厦门大学学报(自然科学版) 2015年4期
钟声平,苏永全,王 军,刘洪涛,毛 勇
(厦门大学海洋与地球学院,福建 厦门361102)
日本囊对虾(Marsupenaeus japonicus)俗称:花虾、竹节虾,是具有重要经济价值的养殖虾类[1],其广泛分布于印度、西太平洋地区,在我国主要分布在长江口以南沿海,其肉质鲜嫩可口、适应性强、可活体运输、销售价格高[2],是我国主要的养殖虾类之一.相对于其他养殖对虾种类,日本囊对虾生长较慢、成活率较低,严重影响日本囊对虾养殖产业的发展.
选择育种是传统、基本的育种方法,是水产养殖动物性状改良、提高生长速度以及成活率的最重要途径,自20世纪70年代以来,在水产动物育种领域得到广泛应用,已在大西洋鲑鱼(Salmo salar)、罗非鱼(Oreochromis niloticus)和凡纳滨对虾(Litopenaeus vannamei)等种类中成功培育出养殖新品种[3].有关日本囊对虾遗传育种理论较少,也还没有日本囊对虾新品种的报道.
早期的选择育种通常以个体表型值的大小进行选种,但个体的表型值容易受到环境的影响,难以将各种环境效应、母系效应等非加性遗传效应剔除,导致其生长选育进展较慢[4].本文通过规模构建日本囊对虾全同胞家系,并在高位池的养殖环境中进行生长性状对比测试实验,采用动物模型分析其遗传方差组分,估算日本囊对虾G1群体(第1世代选育群体)生长性状遗传力和遗传相关,获得了我国东南沿海日本囊对虾基础选育群体的生长遗传参数,为采用家系选育结合家系内个体选育的综合育种技术开展日本囊对虾生长选育提供了理论依据.
1 材料与方法
1.1 亲本交配及催产
实验所用亲本为课题组培育的台湾海峡群体的日本囊对虾基础群体的后代,2009年6月,挑选生长较快的雌虾和雄虾(雌虾体质量32~45g,雄虾体质量24~39g),分成6个组合(每个组合雌雄各150尾)放入高位池中的6个围网(24m3/网)中进行自然交尾.期间投喂“福星”牌亲虾强化饲料促进性腺发育.7月份参考孔杰等[5]日本囊对虾家系构建方法挑选带精荚、性腺III期的雌虾,烫镊法摘除单侧眼柄后,每尾雌虾单独放入室内水泥池(5m3/池)催产,每尾雌虾生产的后代为单独一个家系,在一个月内构建了56个全同胞家系.
1.2 幼体培育及标记
参考中国明对虾(Fenneropenaeus chinensis)幼体培育及标记的方法[6]略作修改.每个家系以灯诱法各选留1万尾活力较好的无节幼体,以虾片、轮虫、丰年虫等为饵料,培育到仔虾第10天(P10)时,进行第1次标准化分苗,每个家系选留2 000尾仔虾(P10),体长0.9~1.2cm;到仔虾第25天(P25)进行第2次标准化分苗,每个家系选留600尾仔虾(P25),体长2.4~3.2 cm,到仔虾第40天(P40)时,每个家系随机挑选300尾,体长4.2~5.1cm,在其腹节处注射不同组合的荧光物(美国西北海洋技术公司,红、褐、绿、黄、紫、蓝等6种颜色)作为家系标记.分别培育一周后,从56个家系中挑选存活率高且生长快的30个家系,放入室外180m3高位池混合养殖(50尾/m3),进行生长性状测试对比试验.
1.3 生长性状数据采集
混养在高位池的30个家系,按高位池养殖管理操作投喂“福星”牌日本囊对虾专用饲料,视情况泼洒“南水利生”牌益生菌、电解多维以改善水质保持良好的养殖环境.养殖至103d时,每个家系随机取30尾,按照《日本对虾亲虾标准(SC/T 2040—2011)》[7],以刻度尺和电子天平(精度0.001g)测量其体长和体质量等数据.
1.4 数据处理及统计分析
通过SAS软件对体长、体质量数据进行正态分布检验[8].采用约束最大似然法(REML)来估计方差组分,其遗传参数评估的动物模型为:
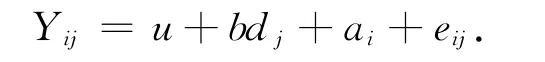
式中Yij为个体性状测量值,u为总体均值,b为混养前体长、体质量对收获体长、体质量的回归系数,dj为第j个家系混养前体长、体质量(协变量);ai为第i尾对虾的加性遗传效应(随机效应),eij为随机残差效应(随机效应).统计分析采用WOMBAT软件[9],生长性状测量数据及系谱数据按照软件要求的格式整理,迭代运行使得统计模型似然值及统计参数收敛,计算生长性状的遗传力及表型相关、遗传相关.遗传力(h2)的计算公式[10]为,表型相关及遗传相关(rp/g(XY))的计算公式[10]为其中和e分别为加性方差和随机误差,σp/g(XY)为性状X与性状Y的协方差,σp/g(X)和σp/g(Y)分别为性状 X 和性状Y 的加性方差.
2 结果与分析
本研究中选育的G1世代日本囊对虾体长、体质量遗传力均属于高遗传力水平(h2>0.3),通过家系选育能获得较高的遗传进展;其体长与体质量间呈高度遗传相关(r>0.75)并且统计检验结果显著(p<0.001),体长或者体质量均可作生长选育的评估性状.
2.1 数据统计特征
将30个家系900个测量数据以SAS软件整理,以体长、体质量测量数据的平均值、变异系数及数据正态性检验显著性等描述统计指标,结果如表1所示,并按家系分组绘制体长、体质量数据的箱型图(图1和2)和体长、体质量散点图(图3).结果显示:日本囊对虾不同家系、不同个体生长速度存在差异,体质量的变异系数大于体长的变异系数.体长与体质量的测量数据正态性检验显著性p值大于0.05,显示其测量数据呈正态分布,可用于下一步的遗传参数的评估.体长与体质量测量数据的散点图呈高度的线性关系,表明高位池养殖环境下,日本囊对虾G1群体体长与体质量间存在很大的统计学关联.

表1 选育日本囊对虾体长、体质量数据统计特征Tab.1 Descriptive statistics traits of G1 M.japonicus
2.2 体长、体质量性状遗传参数
体长、体质量生长性状的表型、遗传参数值如表2所示.养殖103d(个体体质量(21.14±5.77)g)的日本囊对虾G1群体已达到市售规格,此时的体长和体质量的遗传力分别为0.43±0.11和0.44±0.12,均属于高遗传力(h2>0.3),其统计模型的似然比检验结果也是显著的(p<0.05).结果显示:日本囊对虾G1群体生长性状为高遗传力水平(0.43~0.44),可采用个体选择的育种方法来改良生长性状.同时,体长、体质量遗传相关值为0.96±0.02,表型相关值为0.95±0.03,属高度正相关(r>0.75),可将体长或者体质量纳入育种的目标性状.
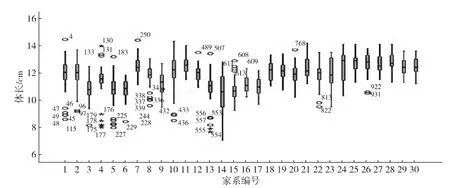
图1 日本囊对虾G1群体体长数据箱型图Fig.1Box plots of body length for the G1 M.japonicus
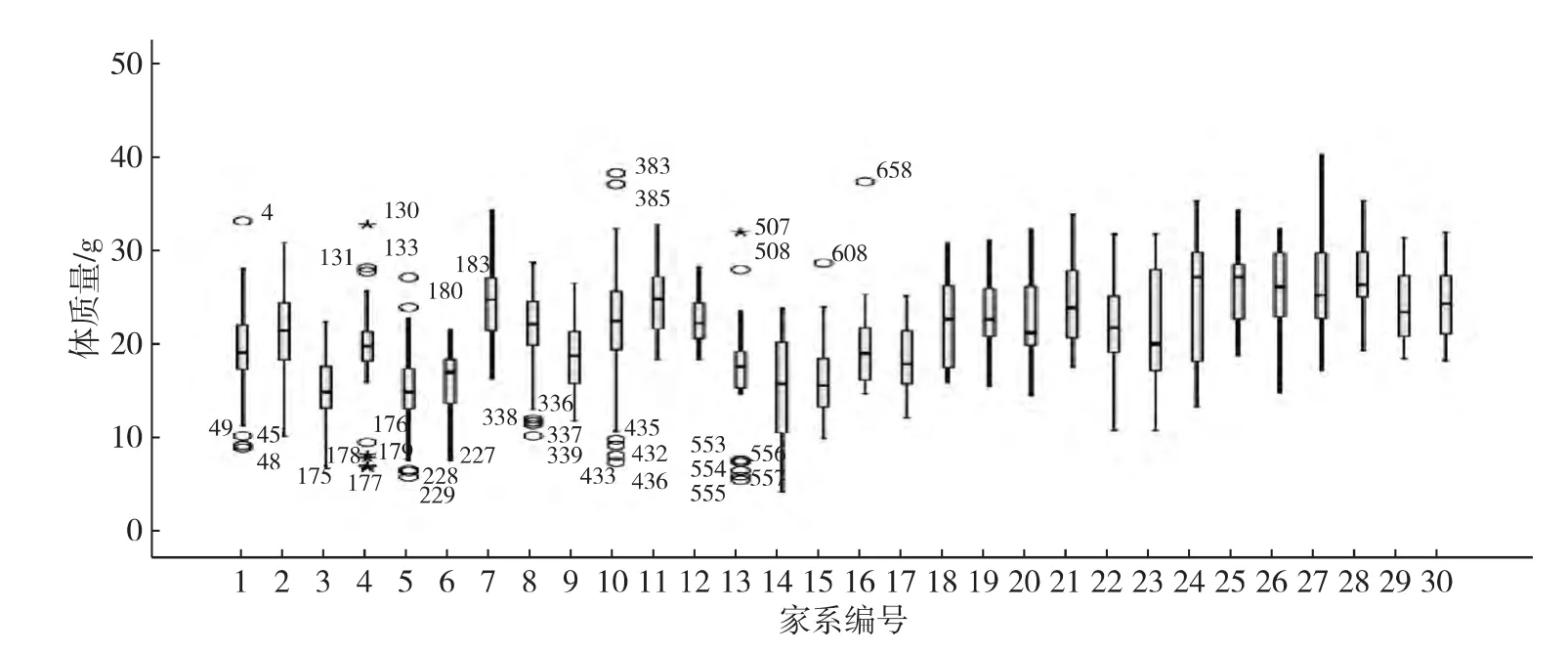
图2 日本囊对虾G1群体体质量数据箱型图Fig.2Box plots of body weight for the G1 M.japonicus
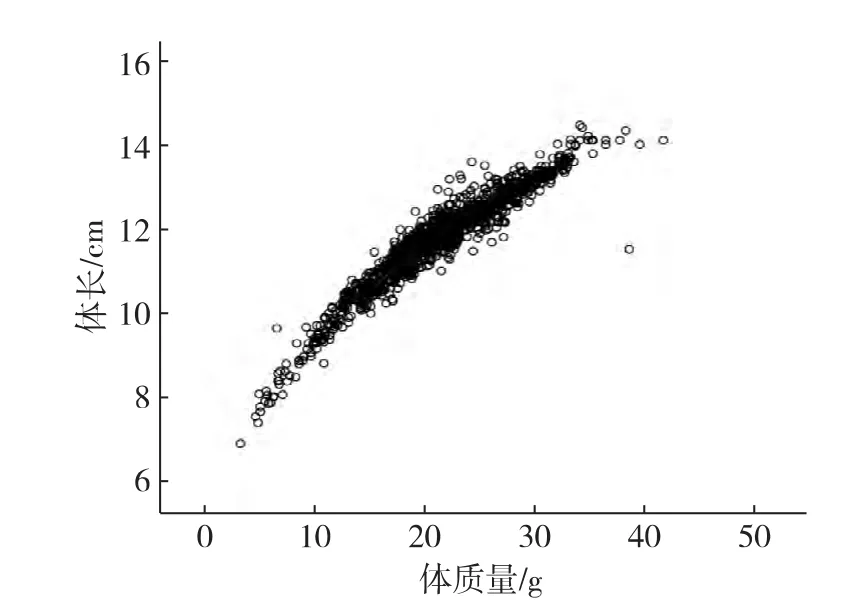
图3 日本囊对虾G1群体体长、体质量散点图Fig.3Scatter diagram between body length and body weight for the G1 M.japonicus
3 讨 论
20世纪80年代,挪威水产研究所(AKVAFORSK)首先使用线性混合统计模型来评估大西洋鲑鱼的生长性状遗传参数,显著提高了其生长遗传进展[4].至20世纪末,人工选育大西洋鲑鱼的生长速度已经高于野生的2倍.本文采用线性混合统计模型,针对基于野生群体亲本来源的基础群体所开展的G1世代选育结果表明,日本囊对虾生长性状的狭义遗传力的范围是0.43~0.44,为高遗传力.该结果与黄付友等[11]采用全同胞组内相关法评估中国明对虾1个选育世代的体质量遗传力的结果一致(0.44~0.53)均属于高遗传力.也与徐如卫等[12]使用全同胞组内相关法估计凡纳滨对虾1个选育世代的体质量遗传力(0.46~0.52)及Kenway等[13]使用线性混合统计模型估计斑节对虾(Penaeus monodon)体质量遗传力 (0.45~0.56)结果一样,同样是高遗传力.

表2 日本囊对虾体长、体质量性状表型和遗传参数Tab.2 Phenotypic and genetic parameters for body length and body weight of M.japonicus
实际上,在对虾科中凡纳滨对虾、斑节对虾和中国明对虾的遗传力研究中发现,不同物种或同一物种在不同的养殖环境中,其体质量的遗传力并不完全相同,遗传力的评估结果不仅与育种工作者采用的统计模型和评估方法密切相关,而且也与评估物种生物学特性、评估群体的遗传背景、养殖环境紧密关联.在日本囊对虾的生长选育的研究中,澳大利亚联邦科学与工业研究组织(CSIRO)对虾育种团队 Hetzel等[14]估计日本囊对虾经过1代群体选育后其G1群体的体质量的现实遗传力范围是0.17~0.32,为中、高度遗传力.相比陆生动物而言,水产动物生长性状存在较高的遗传变异[15],在甲壳类中可高达(20%~35%)[4],远超陆生动物生长性状的遗传变异(7%~10%).与对虾科其他物种相似,本研究中我国东南沿海的日本囊对虾生长性状的遗传力水平也较高,反映了其选育的野生群体均具有较高的生长遗传变异,其高的遗传力水平有利于生长性状的选育,维持稳定的生长选育进展.孔杰等[5]研究结果显示家系选育利用了个体的亲属信息,采用该选育技术来评估遗传参数比群体选育的方法更加准确、可靠,采用家系选育技术进行个体选择的可信度较高.本研究通过构建家系评估了日本囊对虾G1群体的体长、体质量遗传参数,具有较高的可靠性,利用家系选育技术的个体选择可信度较高.
动物的许多性状之间因为内在的遗传机制往往存在着一定的相关性,被称之为表型相关[16].在数量遗传学中,表型相关从相关的机理角度同样剖分为遗传相关和环境相关两部分[4].遗传相关是间接选种和早期选种乃至多性状复合育种的重要基础依据.本研究中,日本囊对虾G1群体的体长与体质量性状间为高度正相关(0.96±0.02),与田燚等[6]研究中国明对虾生长性状间的遗传相关结果一致(0.82~1.00).凡纳滨对虾生长性状间也属于高度遗传正相关(0.75~0.97)[12],其中体长与体质量间遗传相关为0.94,与本研究的结果相近.本研究中日本囊对虾体长与体质量高度遗传正相关的结果提示:控制日本囊对虾体长、体质量性状的基因可能相同或紧密连锁,生长性状间存在类似的调控机制.在生长选育的育种项目中可根据数据测量的难易程度,在保证数据准确性、精确性的前提下,选择体质量或者体长作为目标性状均可达到生长改良的目的.相比较而言,对虾中体质量性状比体长性状更容易受到环境因素的影响,其选种的稳定性较差.所以在日本囊对虾多性状复合育种项目中,可将体质量作为生长改良的重点性状并对体长性状进行加权评估,同时考察存活率与耐温性等性状,提高其生产性能的遗传进展.
相比其他经济种类如凡纳滨对虾中广泛开展的品种改良的研究而言[17],日本囊对虾在我国育种进展较慢,其苗种基本是未经选育的海捕亲虾的后代,生长较慢和成活率低等难题长期困扰着日本囊对虾养殖产业的健康发展.鉴于凡纳滨对虾中优良品种对产业推动的巨大示范效应[18],基于高遗传力的家系选育技术,开发日本囊对虾快速生长的新品种,将有效解决长期困扰日本囊对虾的生长率慢、成活率低等产业技术难题.本文的研究结果为日本囊对虾的生长改良提供了重要的遗传参数数据,为开展多代生长选育,富集生长相关的优良基因并稳定遗传,最终推出新品种奠定了重要的理论基础.
[1]庄志猛,孔杰,石拓,等.日本对虾野生和养殖群体遗传多样性的 RAPD 分析[J].自然科学进展,2001,11(3):250-255.
[2]李义军,李婷,王平,等.日本囊对虾(Marsupenaeus japonicus)3个野生种群和1个养殖种群的形态差异与判别分析[J].海洋与湖沼,2010,41(4):500-504.
[3]Gjedrem T.Genetic improvement for the development of efficient global aquaculture:apersonal opinion review[J].Aquaculture,2012,344:12-22.
[4]王清印,孔杰,江世贵,等.水产生物育种理论与实践[M].北京:科学出版社,2013:21-23.
[5]孔杰,栾生,张天时,等.中国对虾3种育种模式的BLUP遗传评定分析[J].海洋水产研究,2008,29(3):27-34.
[6]田燚,孔杰,栾生,等.中国对虾生长性状遗传参数的估计[J].海洋水产研究,2008,29(3):1-6.
[7]中国水产科学研究院黄海水产研究所.SC/T 2040—2011日本对虾 亲虾[S].北京:中国标准出版社,2011.
[8]冯国双.医学案例统计分析与SAS应用[M].北京:北京大学医学出版社,2011:28-30.
[9]Meyer K.WOMBAT:a tool for mixed model analyses in quantitative genetics by restricted maximum likelihood(REML)[J].Journal of Zhejiang University Science B,2007,8(11):815-821.
[10]Gjedrem T,Baranski M.Selective breeding in aquaculture:an introduction[M].Berlin:Springer,2009:36-39.
[11]黄付友,何玉英,李健,等.“黄海1号”中国对虾体长遗传力的估计[J].中国海洋大学学报:自然科学版,2008,38(2):269-274.
[12]徐如卫,钱昭英,刘小林,等.凡纳滨对虾生长性状遗传参数的估计[J].水产学报,2013,37(5):672-678.
[13]Kenway M,Macbeth M,Salmon M,et al.Heritability and genetic correlations of growth and survival in black tiger prawn Penaeus monodonreared in tanks[J].Aquaculture,2006,259(1):138-145.
[14]Hetzel D,Crocos P J,Davis G P,et al.Response to selection and heritability for growth in the kuruma prawn,Penaeus japonicus[J].Aquaculture,2000,181(3):215-223.
[15]Gjedrem T,Robinson N,Rye M.The importance of se-lective breeding in aquaculture to meet future demands for animal protein:a review[J].Aquaculture,2012,350:117-129.
[16]陈国宏,张勤.动物遗传原理与育种方法[M].北京:中国农业出版社,2009:115-116.
[17]Andriantahina F,Liu X,Feng T,et al.Current status of genetics and genomics of reared penaeid shrimp:information relevant to access and benefit sharing[J].Marine Biotechnology,2013,15(4):399-412.
[18]Coman G J,Arnold S J,Wood A T,et al.Age:age genetic correlations for weight of Penaeus monodon reared in broodstock tank systems[J].Aquaculture,2010,307(1):1-5.
猜你喜欢
西北农业学报(2024年7期)2024-07-12 20:26:59
当代水产(2022年8期)2022-09-20 06:46:22
天天爱科学(2022年9期)2022-09-15 01:12:28
养猪(2022年4期)2022-08-17 07:07:02
当代水产(2022年5期)2022-06-05 07:55:46
科普童话·百科探秘(2022年8期)2022-05-30 10:48:04
昆明医科大学学报(2022年1期)2022-02-28 07:46:32
当代水产(2021年8期)2021-11-04 08:49:04
当代水产(2021年4期)2021-07-20 08:10:54
湖北畜牧兽医(2015年11期)2016-01-11 10:08:24
