
水稻矮杆多分蘖突变体CA648的光合生理特性分析
2015-11-03 04:24刘晓鹏朱允华李合松
亚热带植物科学 2015年1期
刘晓鹏,朱允华,刘 清,史 齐,李合松
(1.湖南农业大学 植物激素与生长发育湖南省重点实验室,湖南 长沙 410128;2.南华大学 药学与生物科学学院,湖南衡阳 421001)
水稻是我国最重要的粮食作物之一[1],杂交水稻的推广应用大幅度提高了水稻单位面积产量[2],超级稻育种计划所确立的理想株型塑造与杂种优势利用相结合的技术路线已取得良好效果[3],其中培育理想株型品种被认为是提高水稻产量途径中最具潜力的方法之一,相关科研工作者希望通过水稻品种设计培育出“新株型”品种,使其最大限度地提高群体光能利用率,从而实现增产目标[4]。
水稻光合能力既受环境影响,更受品种制约,在相同环境中不同品种的光合特性存在较大差异[5]。光合作用的相关产物既是植物自身生长发育的物质基础,也是作物生产力高低的决定性因素。蔗糖除满足植物体内各种生长组织和再生组织的生长需求外,还参于纤维素、储藏物质的生物合成,如淀粉、果聚糖、脂类等[6]。可溶性糖是植物体内一种重要的化合物,其含量的高低反映了植物体内可利用态物质和能量的供应基础[7]。比较分析不同株型水稻材料生长发育过程中光合生理特性的差异,可为新品种选育过程中,考察与光合利用效率相关的某些生理指标提供参考依据。本研究前期获得一个水稻突变体材料CA648,该突变体表型明显不同于对照中花11,株高可受不同生长调节剂处理而显著改变,但其多分蘖特性非常稳定[8]。本文对该突变体及对照的叶绿素含量、蔗糖含量、光合作用以及相关酶活性等生理指标进行比较分析,以期为后续阐明该突变体相关基因的生物学功能提供基础。
1 材料与方法
1.1 材料
粳稻品种中花11(ZH11),突变体丛矮648(CA648)。其中CA648是由T-DNA插入ZH11基因组引起突变的矮杆多分蘖突变体。
1.2 方法
1.2.1 实验设计 试验在湖南农业大学植物激素与生长发育湖南省重点实验室的网室进行。供试种子在常温下浸种72 h后,取发芽种子播种在铺满花园土的陶土盆中育苗。育苗30 d后移栽至盆钵中,单本移植,4次重复。
1.2.2 分蘖数测定 分蘖数从水稻分蘖期(以2014年7月7日为水稻分蘖期)开始,分别在分蘖期开始后10 d(分蘖前期)、20 d(分蘖盛期)、30 d(分蘖盛期)、40 d(分蘖后期)测定,共测4次。每品种测量9~12个单株,取平均值。
1.2.3 叶绿素相对含量测定 采用Konica Minolta SPAD-502进行测量,每盆测量10个水稻叶片,取平均值。
1.2.4 光合作用测定 按王惠群等[9]的方法稍作修改,在水稻分蘖后期晴朗天气的上午 10:00~12:00,采用Li-6400便携式光合作用系统测定净光合速率、胞间CO2浓度、气孔导度和蒸腾速率等光合作用参数,测定环境条件为开放式气路、红蓝光源、光通量密度1000 μmol·m-2·s-1、相对湿度70%和温度20 ℃。选取生长一致的4盆中各1株的顶下第2完全展开叶进行测定。
1.2.5 还原糖、蔗糖含量测定 还原糖的测定根据DNS比色法稍作修改。在分蘖盛期(2014年7月29日)分别取两种材料的新鲜茎和叶置于研钵中,剪碎,加液氮研磨,磨匀后称重并倒入容量瓶,加 20 mL 50%乙醇,45 ℃水浴2 h(重复2次),离心并过滤收集2次上清,蒸馏水定容至100 mL。其余步骤按郝再彬等[10]的方法进行。
蔗糖含量的测定按照张迟[11]的方法。按上述还原糖测定的方法测定样品的还原糖含量(R1),取测完的样品处理液50 mL,加入5 mL 6 mol·L-1盐酸,在68~70 ℃水解15 min,冷却后加2滴甲基红指示剂,用5 mol·L-1氢氧化钠溶液中和,定容至100 mL,摇匀后按上述方法测定还原糖含量(R2)。蔗糖含量的计算公式为:蔗糖含量X= (2×R2 - R1) × 0.95
1.2.6 过氧化物酶活力测定 按照张志良[12]的方法进行测定。酶活计算公式如下:
酶的比活力(0.01ΔA·g-1·min-1)=(ΔA×D) / (0.01×W×t)
式中,ΔA为反应时间内吸光度的变化,t为反应时间(min),D为稀释倍数,W为冷冻干燥样品重(g)。
2 结果与分析
2.1 CA648和ZH11分蘖数比较
从水稻分蘖期(四叶期)开始,分别在分蘖开始后10 d(分蘖前期)、20 d(分蘖盛期)、30 d(分蘖盛期)、40 d(分蘖后期)测定供试材料的分蘖数。由表1可知,CA648的分蘖数在分蘖期间均显著多于ZH11,CA648的最终分蘖数可达20多个,而对照ZH11的分蘖数一般为10个左右。

表1 CA648与ZH11不同时期分蘖数比较Table1 Comparisons of tiller number in different periods between CA648 and ZH11
2.2 CA648和ZH11叶绿素相对含量比较
从水稻分蘖期(四叶期)开始,分别测定分蘖开始后10 d(分蘖前期)、20 d(分蘖盛期)、30 d(分蘖盛期)、40 d(分蘖后期)相同叶序上中上部位的叶片SPAD值。结果表明,随着生长时间延长,CA648与ZH11的叶片SPAD值逐步增大,表明叶绿素相对含量均有所增加,对照ZH11的SPAD值在分蘖开始后 10 d(分蘖前期)、40 d(分蘖后期)均显著高于 CA648(表 2)。

表2 CA648和ZH11叶绿素相对含量比较Table2 Comparisons of relative chlorophyll content in different periods between CA648 and ZH11
2.3 CA648和ZH11光合特性分析
表3表明,CA648在分蘖前期(10 d),叶片的净光合速率、气孔导度和蒸腾速率均显著低于同期的ZH11,而胞间CO2浓度却显著高于ZH11;分蘖盛期(20d),CA648和ZH11植株光合生理指标差异不显著;分蘖盛期(30d),CA648和ZH11叶片的净光合速率出现了显著性差异,其中分蘖后期(40d),ZH11的净光合速率比CA648高4.1%,ZH11的气孔导度、胞间CO2浓度和蒸腾速率都高于CA648。值得注意的是,分蘖中后期CA648分蘖数增加(表 1)。

表3 CA648与ZH11叶片光合作用及蒸腾作用比较Table3 Comparisons of photosynthesis and transpiration between CA648 and ZH11
2.4 CA648和ZH11叶片的还原糖、蔗糖含量比较
在分蘖盛期后,CA648的分蘖与ZH11存在最大差异(表1), CA648叶片中的还原糖含量明显比对照高;而茎杆中的还原糖含量两者间差异不明显(图 1)。CA648叶片中的蔗糖含量低于对照,约为ZH11的77.5%;而茎杆中的蔗糖含量却高于对照,比ZH11高 30.7%(图1)。
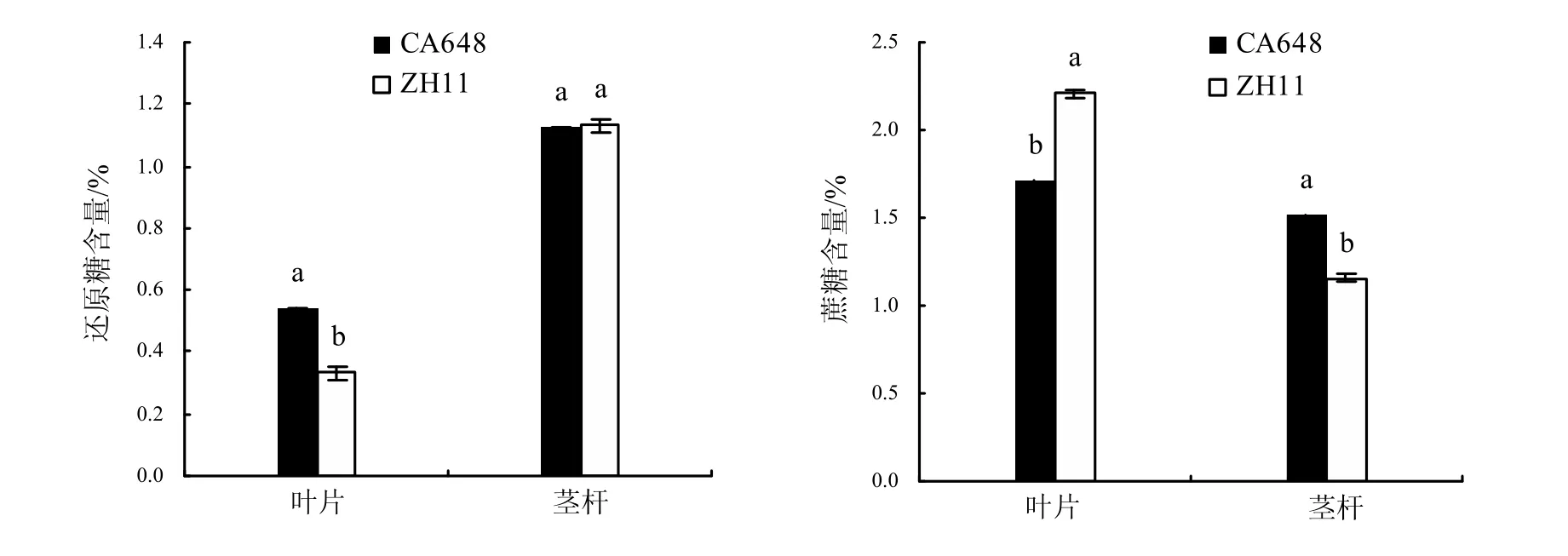
图1 CA648与ZH11的还原糖、蔗糖含量比较Fig.1 Comparisons of the content of reducing sugar and sucrose between CA648 and ZH11
2.5 CA648和ZH11过氧化物酶活力比较
图2表明,进入分蘖期后CA648与ZH11叶片中过氧化物酶活性发生变化,CA648呈现出先升后降的趋势。在7~14 d期间,CA648叶片中的过氧化物酶活力明显高于ZH11,在分蘖期后14 d时,CA648过氧化物酶活力比ZH11高73.0%;14 d后活性开始下降,在21 d时其活性与ZH11无差异。ZH11叶片的酶活性变化在不同时期变化较小,变化趋势是先下降、然后增加再缓慢下降,在28 d时活力最高(157.9 0.01ΔA·g-1·min-1),在 28 d、35 d时叶片的过氧化物酶活力略高于CA648。
图2可知CA648与ZH11茎杆中的过氧化物酶活性变化趋势都是先上升后下降,且CA648茎杆中的过氧化物酶活力均高于ZH11,在14 d时,两者差异达到了16.8%。

图2 CA648和ZH11过氧化物酶的比活力比较Fig.2 Comparisons of peroxidase activity between CA648 and ZH11
3 讨论
水稻株型对产量具有重要影响。水稻育种专家希望通过改良穗、叶、根等农艺性状来获得“理想株型”,以尽可能提高群体的光能利用率和干物质生产能力[13]。本研究的突变体CA648具有矮化、多分蘖特性,其表型在苗期与对照ZH11无差异,进入分蘖期后株高明显低于野生型植株,而分蘖数目明显多于野生型植株[8]。至成熟期时,CA648的不同单株均可形成20多个有效分蘖,而ZH11的有效分蘖数目一般为10个(表1)。
研究表明,引起植物叶片光合效率降低的主要因素包括气孔部分关闭和叶肉细胞光合活性下降,前者使细胞间隙CO2浓度降低,后者则使其增高[14]。光合特性比较结果显示,突变体CA648在分蘖初期气孔导度显著小于ZH11,且净光合速率也低于ZH11,而胞间CO2浓度却显著高于ZH11。其原因除了与CA648叶绿素含量相对较低有关外,还可能与CA648的多分蘖特性有关。CA648比ZH11形成更多的分蘖,较多的分蘖可导致单位面积内分蘖及叶片相互遮挡而影响光合特性。
过氧化物酶在植物生长发育中起着十分重要的作用,可以降解植物体内IAA,而IAA可以促进细胞伸长,从而影响茎的生长,因此茎中过氧化物酶活性增高可能引起植株矮化[15]。CA648叶片中的过氧化物酶活力在分蘖前期与分蘖盛期均高于ZH11,而茎中的过氧化物酶活力在整个分蘖期均高于ZH11。过氧化物酶可降解IAA,而IAA/CTK比值是植物维持顶端优势的关键因素[16]。因此,CA648叶片及茎杆中较高的过氧化物酶活性可导致IAA含量发生改变,进而影响IAA/CTK比值,该结果与已报道的CA648、ZH11两者间植物激素存在显著差异的结果一致[8],推测较高的过氧化物酶活力可能与CA648矮化多分蘖特性之间存在一定的相关性。
CA648和ZH11相关光合产物比较分析表明,在分蘖盛期CA648的叶片、茎杆中的还原糖、蔗糖含量均与ZH11存在差异。其原因可能是CA648需要分配更多的光合产物用于分蘖,也可能是光合效率降低导致光合产物减少而造成的结果,有待进一步研究。
[1] 虞国平.水稻在我国粮食安全中的战略地位分析[J].新西部(理论版), 2009(11): 31—33.
[2] 文杜娟,陈风波.杂交水稻种植对社会经济的影响[J].作物研究, 2014(4): 402—405.
[3] 程式华.中国超级稻育种研究的创新与发展[J].沈阳农业大学学报, 2007(5): 647—651.
[4] 冯荣坤.超高产水稻育种中理想株型研究进展[J].河南农业科学, 2006(7): 19—22.
[5] 唐茜,叶善蓉,单洪丽,罗学平.引进茶树品种光合特性的比较研究[J].四川农业大学学报, 2006,24(3): 303—308.
[6] 乔亮.水稻蔗糖合成酶基因家族的初步研究[D].济南: 山东大学硕士学位论文, 2012.
[7] 黄雪方,金雅琴,李冬林.两种石蒜生长发育期鳞茎可溶性糖、蛋白质及POD活性的变化[J].亚热带植物科学, 2011(3):16—19.
[8] 刘清,童建华,史齐,彭克勤,王若仲,蔺万煌, Mohammed H K,沈革志,萧浪涛.一个矮秆多分蘖水稻突变体的植物激素动态特性分析[J].中国农业科学, 2014(13): 2519—2528.
[9] 王惠群,肖浪涛,杨艳丽,童建华,康朵兰.矮壮素对中薯 3号生理特性的影响研究[J].云南农业大学学报, 2008,23(2):200—204.
[10] 郝再彬,苍晶,徐仲.植物生理实验[M].哈尔滨: 哈尔滨工业大学出版社, 2004.
[11] 张迟.抑制马铃薯Acid Invertase基因表达对块茎淀粉—糖代谢的影响研究[D].武汉: 华中农业大学博士学位论文,2007.
[12] 张志良.植物生理实验指导手册[M].北京: 高等教育出版社, 2000.
[13] 陈庭木,王多明,方兆伟,秦德荣,樊继伟,吕孝财,徐大勇.水稻超高产潜力鉴定重要农艺性状因子分析[J].安徽农业科学, 2013(16): 7081—7085.
[14] 许大全.光合作用气孔限制分析中的一些问题[J].植物生理学通讯, 1997,33(4): 241—244.
[15] Intapruk C, Yamamoto K, Sekine M Takano M, Shinmyo A.Regulatory sequences involved in the peroxidase gene expression in Arabidopsis thaliana[J].Plant Cell Reports, 1994,13: 123—129.
[16] 李春俭.植物激素在顶端优势中的作用[J].植物生理学通讯, 1995(6): 401—406.
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
中国农业科学(2022年8期)2022-05-16
农业科技通讯(2021年1期)2021-03-06
生命科学研究(2021年1期)2021-01-25
中国食品学报(2019年12期)2019-01-13
分析化学(2018年7期)2018-09-17
吉林农业·下半月(2017年7期)2017-07-15
农民致富之友(2017年6期)2017-05-10
上海农业学报(2017年3期)2017-04-10
中国塑料(2016年3期)2016-06-15
