
阿拉善地区人为干扰下啮齿动物群落的β多样性
2015-11-01 03:41:50武晓东张晓东付和平高全荣
天津师范大学学报(自然科学版) 2015年3期
武晓东 ,张晓东 ,袁 帅 ,付和平 ,高全荣
(1.内蒙古农业大学生态环境学院,呼和浩特010019;2.草业与草地资源教育部重点实验室,呼和浩特010019;3.内蒙古自治区林业监测规划院,呼和浩特010020;4.内蒙古医科大学,呼和浩特010059)
早期的生态学家将一定面积上的物种多样性分为2个组成部分[1-3]:①相对于在单个立地或生境上发现的物种数量,即α多样性;②关注在立地中间、栖息地或者梯度中物种的变化,即β多样性.γ多样性是所有立地或栖息地总的多样性,且γ=α×β.这种简单关系已在许多研究中得到应用,特别是处理栖息地中、小尺度生物多样性的差异[4-5].环境异质性和栖息地多样性是群落组成的重要决定因素,许多研究都强调了环境异质性对群落组成的影响[6-9],因此物种分布的空间变化是理解群落动态的关键[10].人类主导的景观使得许多栖息地破碎化,研究此变化对于生物区系的保护管理日益重要[11].Hubbel[12]于2001年提出了生物多样性中性理论,此后学界对大尺度上β多样性的研究较多[13-15],在其测度方法上也提出了加性分配法[16]、相似性法[17]等.国内β多样性的研究较少[18].Dahl等[19]指出由于环境条件的空间变化、物种扩散的限制以及种内相互作用等原因,目前难以确定影响β多样性的具体因素,特别是动物的β多样性.
近年来,由于移民和农业开发,内蒙古阿拉善荒漠原生荒漠植被人为干扰严重,景观破碎化现象极为普遍.作者于2002至2010年在该区对人为干扰下景观不同破碎化中啮齿动物群落的β多样性进行了研究.
1 研究区域自然概况与研究方法
1.1 研究区域自然概况
研究区域位于阿拉善左旗南部的嘉尔嘎勒赛汉镇,东经 104°10′~105°30′,北纬 37°24′~38°25′,地处腾格里沙漠东缘.该地区气候为典型的高原大陆性气候,冬季严寒、干燥,夏季酷热,昼夜温差大,极端最低气温–36℃,最高气温42℃,年平均气温9.0℃,无霜期156 d.年降水量45~215 mm,降水极不均匀,主要集中在7—8月份.年蒸发量3 000~4 700 mm.土壤为棕漠土,淋溶作用微弱,土质松散、瘠薄,表土有机质含量(质量分数)1%~1.5%,含有较多的可溶性盐.草地类型是典型的温性荒漠,植被稀疏,结构单调,覆盖度低(1%~20%).植物种类贫乏,主要以旱生、超旱生和盐生的灌木、半灌木、小灌木和小半灌木为主.建群植物以藜科(Chenopodiaceae)、菊科(Compositae)和蒺藜科(Zygop-hyllaceae)为主,其次为蔷薇科(Rosaceae)、柽柳科(Tamaricaceae).地形起伏不平,丘陵、沙丘与平滩相间.
在原生生境植被条件一致的情况下,依据该地区对草地利用方式的不同,选择4种不同干扰类型的生境作为取样样区,样区的植被特征如下:
禁牧区:在原生植被基础上,自1997年开始围封禁牧,到本研究开始的2002年已禁牧5年以上,植被以红砂(Reaumuria soongorica)建群,其次为霸王(Sarcozygium xanthoxyln)、驼绒藜(Ceratoides latens)、油蒿(Artemisia ordosica)等小灌木,草本以白草(Pennisetum flaecidum)为优势种,其次为糙隐子草(Cleistogenes squarosa), 伴 生 有 雾 冰 藜(Bassia dasyphylla)等植物,植被盖度29.5%.土壤为灰漠土,水分含量(质量分数)4.40%~8.22%.
轮牧区:在原生植被基础上,1995年开始采取围栏轮牧的利用方式,轮牧小区的放牧间隔期为一个月.植被以霸王建群,其次为红砂、刺叶柄棘豆(Oxytropis aciphylla)等多年生小灌木,伴生有骆驼蓬(Peganum harmala)、雾冰藜等一年生植物,植被盖度24.5%.土壤为灰漠土,水分含量2.46%~9.32%.
过牧区:在原生植被基础上,自由放牧利用,而且是全年连续放牧.植被以白刺(Nitraria sphaercarpa)、霸王建群,伴生有红砂、刺叶柄棘豆、糙隐子草等多年生植物和条叶车前(Plantago lessingii)、牛心朴子(Cynanchum komarovii)等一年生植物,植被盖度8.5%.土壤为灰漠土,表面严重沙化,水分含量(质量分数)4.66%~12.46%.
开垦区:在原生植被基础上,1994年开垦.植被主要以人工种植的梭梭(Haloxylon ammodendron)、沙拐枣(Calligonummongolicum)等建群,伴生有雾冰藜、条叶车前等一年生杂类草,植被盖度可达65%.土壤为灰漠土,水分含量(质量分数)2.77%~10.58%.
1.2 研究方法
1.2.1 啮齿动物调查方法
2002—2010 年在上述4种不同干扰类型的生境中,每种生境均设计2个动物标志流放样地,面积分别为1 hm2,采用标志重捕法.每年4—10月份的月初进行野外调查取样,每个样地布放56个鼠笼,笼距和行距为15 m×15 m,每月连捕4 d.对捕获过的跳鼠采用带编号的金属环进行标记,其余鼠种用剪趾法标记.记录所捕个体的种名、性别、体重、繁殖状况及捕获笼子的行列号,标志后原地释放.
1.2.2 统计方法
统计研究期内捕获的啮齿动物物种数,运用不同干扰生境中啮齿动物物种数,计算每年啮齿动物群落的β多样性指数.依据当地人为干扰下景观破碎的情况将研究区域内生境梯度的变化按照:禁牧区→轮牧区,轮牧区→过牧区,过牧区→开垦区,禁牧区→过牧区,禁牧区→开垦区,轮牧区→开垦区进行划分.分别运用Jaccard指数(CJ)、Sorenson指数(Cs)和Cody(βC)指数来计算啮齿动物群落的β多样性.
CJ指数和Cs指数反映了地区间物种组成的相似程度;βC通过对2个地区新增加物种数和失去物种数进行比较,反映不同地区间物种组成的变化.计算公式如下:

式中:j为2个群落或样地共有种数;a和b分别为样地A和样地B的物种数;gH为沿生境梯度H增加的物种数目;lH为沿生境梯度H失去的物种数目,即在上一个梯度中存在而在下一个梯度中没有的物种数目.
CJ1、CJ2、CJ3、CJ4、CJ5、CJ6分别代表禁牧到轮牧、轮牧到过牧、过牧到开垦、禁牧到过牧、禁牧到开垦、轮牧到开垦的 Jaccard 指数;CS1、CS2、CS3、CS4、CS5、CS6分别代表 6 个生境梯度的 Sorenson 指数;βC1、βC2、βC3、βC4、βC5、βC6分别代表6个生境梯度的Cody指数.
各鼠种在不同干扰类型中的捕获量比例以每次捕到的动物个体计算,分别统计各鼠种在2个样地中每年的个体总数(捕多次仅算1次),计算其均值作为该年的个体数.其计算公式为:

式中:Pi为第i种个体的捕获量比例;ni为第i种个体总捕获数量.
利用SAS9.2统计软件的单因素方差分析和主成分分析检验每年啮齿动物群落3种β多样性指数不同梯度间以及不同年度间的差异性.
2 结果
2.1 啮齿动物群落的物种组成
2002—2010年4 种干扰生境共捕获9个鼠种,分属3科,分别为:跳鼠科(Dipodidae)3种,包括五趾跳鼠(Allactaga sibirica),三趾跳鼠(Dipus sagitta),蒙古羽尾跳鼠(Stylodipus andrewsi);仓鼠科(Cricetidae)5种,包括:子午沙鼠(Meriones meridianus),长爪沙鼠(Meriones unguiculatus),小毛足鼠(Phodopus roborovskii),黑线仓鼠(Cricetulus barabensis),短耳仓鼠(Cricetulus eversmanni);松鼠科(Sciuridae)1 种,为阿拉善黄鼠(Spermophilus alaschanicus).群落中各组成鼠种的捕获量比例见表1,4种干扰生境9年的累计捕获鼠种相差不大,轮牧区、过牧区和开垦区均为8种,禁牧区为7种.
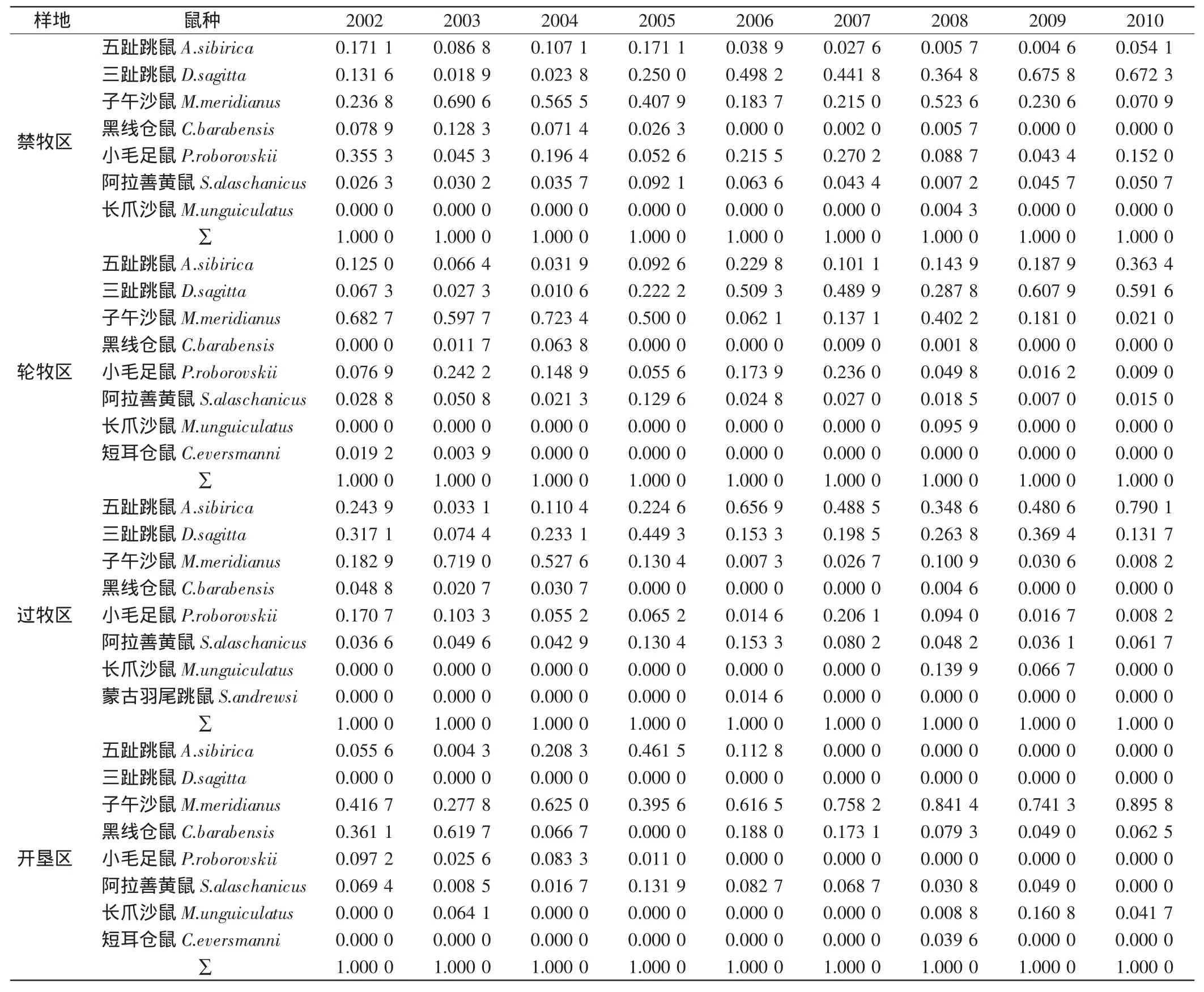
表1 啮齿动物群落中各物种的捕获量比例Tab.1 Proportion of captured rodentsof each species in rodent community
2.2 啮齿动物群落β多样性
2002—2010 年按照干扰梯度测得的β多样性的3种指数结果及变化特征见表2—表4.

表2 啮齿动物Jaccard指数Tab.2 Jaccard diversity indicesof rodent communities
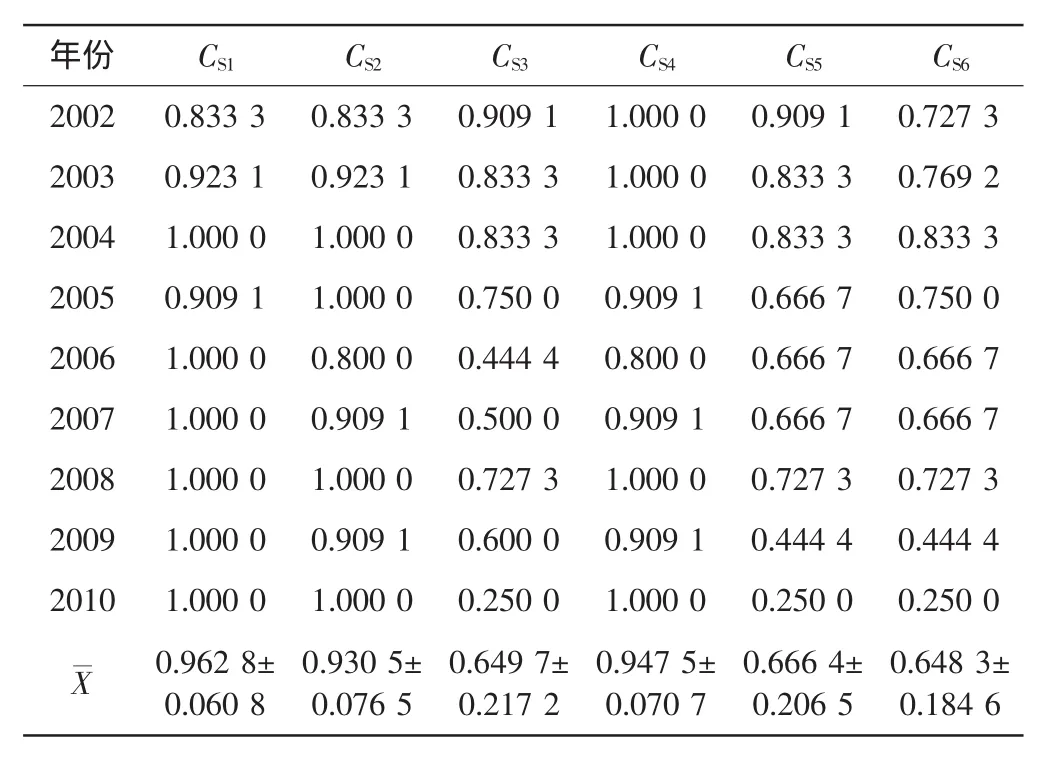
表3 啮齿动物群落Sorenson多样性指数Tab.3 Sorenson diversity indicesof rodent communities

表4 啮齿动物群落Cody多样性指数Tab.4 Cody diversity indicesof rodent communities
由表2和表3可知,过牧→开垦、禁牧→开垦、轮牧→开垦的Jaccord指数和Sorenson指数均相对较小,相互间的差异没有统计学意义(p>0.05),而禁牧→轮牧、轮牧→过牧、禁牧→过牧之间的差异具有高度统计学意义(p<0.01).年度变化中只有2010年与其他年份之间的差异具有统计学意义(p<0.05).由表4可知,过牧→开垦、禁牧→开垦、轮牧→开垦的Cody指数均相对较大,且相互间差异较小,而禁牧→轮牧、轮牧→过牧、禁牧→过牧之间的差异具有高度统计学意义(p<0.01).年度变化中只有2010年与其他年份之间的差异具有统计学意义(p<0.05).
由以上结果可看出,过牧→开垦、禁牧→开垦、轮牧→开垦梯度的3个指数同其他梯度的差异均具有统计学意义(p<0.05).这表明开垦区β多样性的变化与原生境极为不同.
为深入了解各种梯度物种组成的变化特征,作者对禁牧区、轮牧区、过牧区和开垦区4种生境类型中的啮齿动物群落组成种进行PCA分析,结果见表5.
由表5可知,各干扰区内组成群落优势种的地位存在较大差别.禁牧区在第一、第二和第三维主成分贡献率较大者为三趾跳鼠、长爪沙鼠、小毛足鼠;过牧区在第一、第二和第三维主成分贡献率较大者为:五趾跳鼠和三趾跳鼠;轮牧区在第一、第二和第三维主成分贡献率较大者为:三趾跳鼠、小毛足鼠、长爪沙鼠;开垦区在第一、第二和第三维主成分贡献率较大者为:子午沙鼠、黑线仓鼠、长爪沙鼠.该结果进一步说明在人为干扰下,景观破碎化过程中啮齿动物群落β多样性变化明显,特别是在原生基底荒漠景观被开垦为农田后其β多样性变化最大,这主要是由于出现了新增加物种和失去了一些原生物种所致.
3 结论
本研究中,阿拉善荒漠区由于人为干扰严重,整体景观的异质性增加.Nekola等[20]推测β多样性与环境异质性的增长呈正相关关系.在阿拉善荒漠区,随着景观异质性的增加,即生境复杂程度的增加,β多样性也随之增加,特别是过牧→开垦、禁牧→开垦和轮牧→开垦的Jaccord和Sorenson多样性指数较低,Cody多样性指数较高,表明原生基底荒漠景观被开垦为农田后其β多样性变化最大,进一步说明环境变化的剧烈程度是与β多样性变化的剧烈程度相一致.而这种β多样性的增加并非是物种数量的增加所致,而是由于景观破碎化后,啮齿动物主要种群对不同干扰条件的敏感性不同[21],在原生植被破碎化后,群落中原有优势鼠种消失并且新物种增加.

表5 啮齿动物群落组成种PCA分析Tab.5 PCA analysisof rodent communities under different disturbance
[1]MELOA S,RANGELTFL V B,DINIZ-FILHO JA F.Environmental drivers of beta-diversity patterns in New-World birds and mammals[J].Ecography,2009,32(2):226-236.
[2]WHITTAKER RH.Vegetation of the Siskiyou Mountains,Oregon and California[J].Ecological Monographs,1960,30(3):279-338.
[3]WHITTAKER RH.Evolution and measurement of species diversity[J].Taxon,1972,21(2/3):213-251.
[4]KOLEFFP,GASTON K J,LENNON JJ.Measuring beta diversity for presence-absence data[J].Journal of Animal Ecology,2003,72(3):367-382.
[5]MAGURRANAE.Measuringbiologicaldiversity[M].Oxford:Blackwell,2004.
[6]SRIVASTARADS.Using local-regional richnessplotsto test for species saturation:pitfallsand potentials[J].Journal of Animal Ecology,1999,68(1):1-16.
[7]LOREAUM.Are communities saturated?On the relationship between α,β and γ diversity[J].Ecology Letters,2000,3(2):73-76.
[8]GERINGJC,CRISTTO.Thealpha-beta-regionalrelationship:providing new insights into local-regional patterns of species richness and scale dependence of diversity components[J].Ecology Letters,2002,5(3):433-444.
[9]WAGNERhh,WILDI O.Realistic simulation of the effects of abundance distribution and spatial heterogeneity on non-parametric estimatorsof species richness[J].Ecoscience,2002,9(2):241-250.
[10]WALLA TR.,ENGENS,DEVRIESPJ,et al.Modeling vertical betadiversity in tropical butterfly communities[J].Oikos,2004,107(3):610-618.
[11]HOLLING C S.Cross-scale morphology,geometry and dynamics of ecosystems[J].Ecological Monograph,1992,62(4):447-502.
[12]HUBBEL SP.The unified neutral theory of biodiversity and biogeography[M].Princeton:Princeton University Press,2001.
[13]RODRIGUEZ P,ARITAh T.Beta diversity and latitude in North American mammals:testing the hypothesis of covariation[J].Ecography,2004,27(5):547-556.
[14]QIANh,BADGLEY C,FOX D L.The latitudinal gradient of beta diversity in relation to climate and topography for mammals in North America[J].Global Ecologyand Biogeography,2009,18(1):111-122.
[15]JANKOWSKI JE,CIECKA A L,MEYER N Y,et al.Beta diversity along environmental gradients:implications of habitat specialization in tropical montane landscapes[J].Journal of Animal Ecology,2009,78(2):315–327.
[16]CRIST T O,VEECH J A.Additive partitioning of rarefaction curves and species-area relationships:unifying α-,β-and γ-diversity with samplesizeand habitatarea[J].Ecology Letters,2006,9(8):923-932.
[17]CHAO A,CHAZDON R L,COLWELL R K,et al.A new statistical approach for assessing similarity of species composition with incidence and abundancedata[J].Ecology Letters,2005,8(2):148-159.
[18]蒋志刚,马克平.保护生物学的现状、挑战和对策[J].生物多样性,2009,17(2):107-116.
[19]DAHLC,NOVOTNY V,MARAVEC J,et al.Beta diversity of frogs in the forests of New Guinea,Amazonia and Europe:contrasting tropical and temperate communities[J].Journal of Biogeography,2009,36(5):896-904.
[20]NEKOLA J C,WHITE P S.The distance decay of similarity in biogeography and ecology[J].Journal of Biogeography,1999,26(4):867-878.
[21]武晓东,付和平.人为干扰下荒漠啮齿动物群落格局—变动趋势与敏感性反应[J].生态学报,2006,26(3):849-861.
猜你喜欢
疯狂英语·新读写(2024年2期)2024-03-20 01:41:16
海外星云(2021年6期)2021-10-14 07:20:46
西藏艺术研究(2021年3期)2021-06-02 09:37:16
今日农业(2020年19期)2020-12-14 14:16:52
动漫星空(兴趣百科)(2020年5期)2020-06-10 03:12:02
西藏农业科技(2019年1期)2019-07-25 00:37:08
农民致富之友(2018年20期)2018-11-25 15:35:58
消费导刊(2016年7期)2017-08-12 13:23:32
中国农业科学(2017年8期)2017-05-03 09:02:42
甘肃畜牧兽医(2016年12期)2016-09-21 01:37:12
