
绿针假单胞菌HT66中luxR_19对吩嗪-1-甲酰胺合成的影响
2015-10-25 02:58陈亚汶彭华松王威张雪洪
生物技术通报 2015年8期
陈亚汶 彭华松 王威 张雪洪
(上海交通大学生命科学技术学院 微生物代谢国家重点实验室,上海 200240)
绿针假单胞菌HT66中luxR_19对吩嗪-1-甲酰胺合成的影响
陈亚汶 彭华松 王威 张雪洪
(上海交通大学生命科学技术学院微生物代谢国家重点实验室,上海200240)
旨在研究调控基因luxR_19对绿针假单胞菌HT66中的抑菌物质吩嗪-1-甲酰胺(phenazine-1-carboxamide,PCN)合成及菌体生长的影响,通过分子操作构建了luxR_19基因敲除株、回补菌株、过表达菌株以及luxR_19(G85A)点突变菌株,检测了目标菌株生长曲线、PCN发酵结果、细菌泳动性与从动性等。结果显示,将luxR_19敲除后PCN产量明显降低,对luxR_19基因进行过表达,发现能够将PCN产量提高2倍;luxR_19对于吩嗪合成的多个调控基因以及合成基因表达均产生影响;luxR_19的敲除对HT66的生长及泳动性不产生影响,而对细菌的从动性有一定的促进作用;luxR_19(G85A)点突变菌株吩嗪产量较野生型相差很小。结果表明,luxR_19对PCN的合成为正调控基因,一定程度上影响HT66的从动性,第254位碱基突变对PCN合成产量不产生影响。
绿针假单胞菌;HT66;吩嗪-1-甲酰胺;LuxR家族转录调控因子
天然吩嗪类衍生物是一类对环境安全、对植物病原真菌具有广谱抑菌活性的微生物源抗生物质[1]。吩嗪类衍生物种类繁多,主要包括吩嗪-1-羧酸(phenazine-1-carboxylic acid,PCA)、1-羟基-吩嗪(phenazine-1-hydroxy)、2-羟基-吩嗪(phenazine-2-hydroxy,2-OH-PHZ)、吩嗪-1-甲酰胺(phenazine-1-carboxamide,PCN)等[2]。合成吩嗪类衍生物的微生物主要是假单胞菌和链霉菌,绿针假单胞菌被公认为是一种安全、有效的植物根际促生菌,除了良好的抑菌作用,还能促进作物生长,目前仍然没有关于其致病性的相关报道,可以认为其是一种环境友好的促生菌[3-5]。据报道,许多绿针假单胞菌能够分泌多种具有抑菌活性的次级代谢产物,例如:吩嗪类衍生物、藤黄绿脓菌素、硝吡咯菌素、氢氰酸以及杀虫蛋白P. fluroscent insect toxin D(FitD)等[6,7]。其中,吩嗪类衍生物的分泌是其最为重要的抑菌机制之一。在前期的研究报道中,已阐明了绿针假单胞菌中吩嗪生物合成机理,发现了多个参与吩嗪合成的调控因子,初步构建了吩嗪的合成调控网络,这为利用微生物高效合成吩嗪类物质打下了良好的基础。但吩嗪类物质的产量仍然较低,影响了其作为生物农药的开发前景,因此,研发吩嗪类物质的高产菌株十分迫切。
Pseudomonas chlororaphis HT66是由本实验室从上海郊区水稻根际分离所得。其除了保留绿针假单胞菌的普遍性质外,更具有两大优点:一是其分泌的吩嗪类衍生物几乎全为PCN。据报道,在中性条件下,PCN的抑菌性远远高于PCA,为PCA的10倍[4],其他公开报道的绿针假单胞菌,如GP72、30-84以及O6等菌株,都产生多个吩嗪类衍生物,包括PCA、2-OH-PHZ[5,8-10],这使得吩嗪类化合物的分离更为简便。二是在前期的实验中发现野生型HT66菌株,其PCN的产量高达400 mg/L,高于目前所报道的各假单胞菌野生型菌株的吩嗪产量,这提供了将其作为构建PCN高产菌株的研发优势。
在实验室前期对HT66进行化学诱变的实验中,通过多轮诱变最终得到P3株,其PCN产量达到了1 597 mg/L,较同一批次发酵的野生型而言产量提高了3倍[11]。经过全基因组测序,发现共有244个碱基发生了突变,其中有3个LuxR家族转录调控因子发生了突变。LuxR家族转录调控因子多与次级代谢产物调控相关[12]。在HT66野生株中,共有24个luxR家族调控因子,按照3个突变的luxR基因在基因组中出现的先后顺序排列,分别是luxR_13(M217_04406)、luxR_14(M217_04413)和luxR_19(M217_05833),其中luxR_19的第254个碱基由G突变为C,导致第85位氨基酸由Gly突变为Ala。通过氨基酸序列分析发现,在第85位氨基酸前后10-20个氨基酸位点均高度保守,与其活性密切相关。因此,luxR_19这个基因是否与吩嗪的合成相关,其第85位氨基酸的突变是否会对吩嗪合成调控产生影响,是我们首先关心的问题。同时,菌体的生长状况在一定程度上影响到次级代谢产物的合成,luxR_19是否会影响到菌株的生长状况,同样值得关注。
本研究通过分子操作手段,对luxR_19基因进行敲除,回补以及过表达,以研究luxR_19基因对HT66合成吩嗪能力的影响,进一步通过luxR_19(G85A)点突变菌株的构建,研究其对吩嗪合成的影响,旨在为进一步了解PCN高产的调控机制,构建高产PCN菌株打下良好的基础。
1 材料与方法
1.1 材料
1.1.1 菌株与质粒 本研究中所用到的菌株,质粒以及引物见表1和表2。野生型绿针假单胞菌HT66作为构建突变菌株的出发菌株。pK18mobsacB质粒用于得到不包含目的片段的基因片段。质粒pK18CL用于敲除luxR_19基因。质粒pK18CP用于在野生型HT66上获得点突变菌株。pME6032质粒用于构建回补以及过表达质粒。pME-luxR分别用于在野生型HT66中过表达luxR_19基因以及在HT66△luxR菌株中回补该基因。
1.1.2 培养基 Luria-Bertani(LB)培养基:10.0 g/L胰蛋白胨,5.0 g/L酵母提取物,10.0 g/L NaCl,固体培养基加入12.0 g/L琼脂。E. coli的培养温度为37℃,P. chlororaphis的培养温度为28℃。
King’s Broth(KB)培养基:20.0 g /L胰蛋白胨,15.0 mL/L甘油,0.514 g/L K2HPO4,0.732 g/L MgSO4,固体培养基加入12.0 g/L琼脂,用于培养P. chlororaphis,培养温度为28℃。
抗生素浓度:对于E. coli:50 μg/mL kanamycin,15 μg/mL tetracycline;对于P. chlororaphis:50 μg/mL kanamycin,125 μg/mL tetracycline以及100 μg/mL Am-picillin。

表1 本研究所用的菌株和质粒
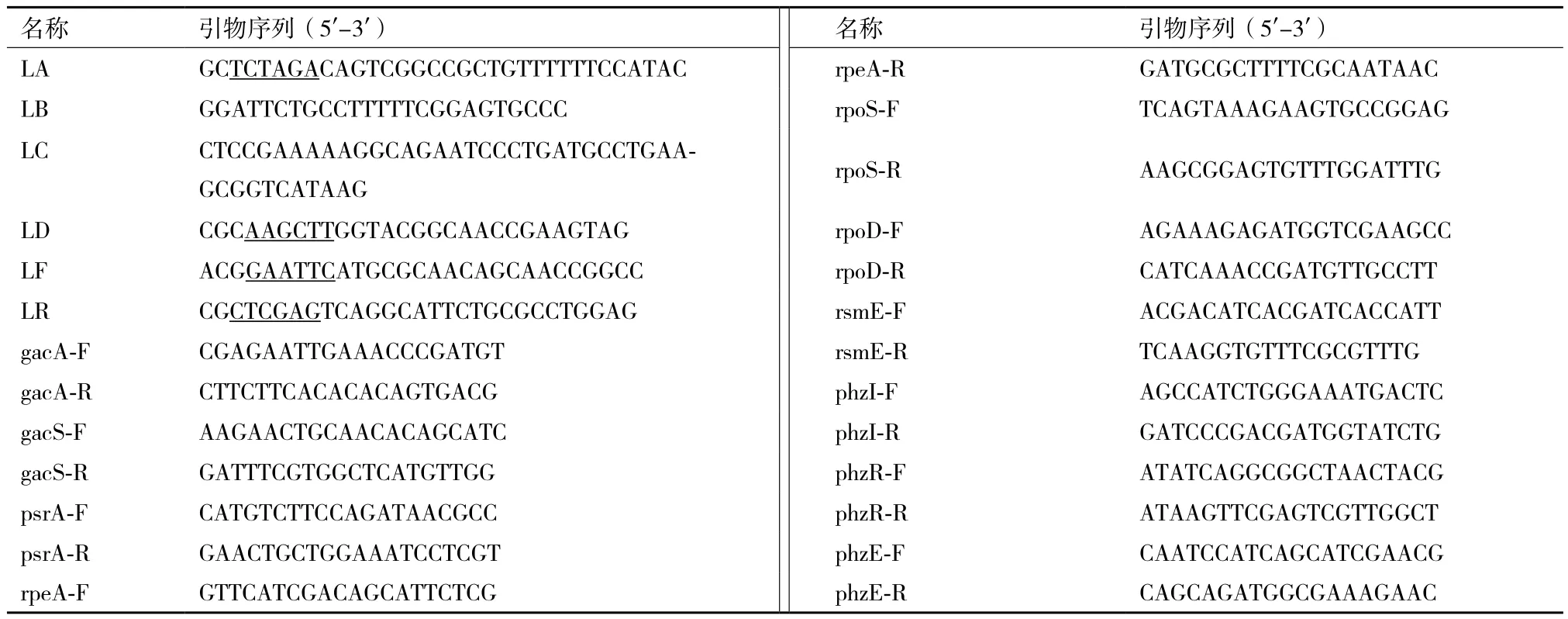
表2 本研究所用引物序列
泳动性(Swimming)实验培养基:10.0 g/L 胰蛋白胨,5.0 g/L NaCl以及3.0 g/L琼脂糖。
从动性(Swarming)实验培养基:5.0 g/L葡萄糖,10.0 g/L胰蛋白胨,5.0 g/L酵母提取物和5.0 g/L琼脂。
1.2 方法
1.2.1 luxR_19无痕敲除菌株HT66CL的构建 设计两对引物LA(Xba I)-LB,LC-LD(Hin d III)用于对luxR_19基因的敲除。利用LA-LB从HT66基因组上扩增M217_05833基因上游673 bp,利用LC-LD从HT66基因组上扩增luxR_19基因下游791 bp。利用LB与LC之间20 bp的同源片段,通过融合PCR将两段基因片段连接在一起。利用限制性内切酶Xba I和Hin d III对获得的1.5 kb的融合片段进行酶切,将所获得的片段克隆至pK18mobsacB质粒中。所获得的质粒pK18CL首先被转入E. coli S17-1(λpir),之后通过接合转移至P. chlororaphis HT66中,获得HT66C1菌株。将HT66C1菌株涂布于含有100 μg/mL Ampicillin(Amp)和50 μg/mL kanamycin(Kan)的LB平板上以筛选出单交换菌株,之后挑选单菌落涂布于含有15%蔗糖的LB平板上以获得双交换菌株。获得的菌株分别点种于含有50 μg/mL Kan的LB平板和无抗LB平板上,挑选在Kan 50 平板上不生长,在无抗LB平板上生长的菌株,利用PCR以及测序进行验证,最终得到无痕敲除luxR_19的菌株HT66CL。
1.2.2 luxR_19基因回补与过表达 利用引物LF(Eco R I)-LR(Xho I)从野生型HT66基因组中通过PCR扩增luxR_19基因,总长度为861 bp。利用限制性内切酶Eco R I和Xho I同时对扩增片段以及质粒pME6032进行酶切,连接得到pME-luxR质粒。将pME-luxR质粒分别电击转化至△luxR以及HT66野生株中,得到回补以及过表达菌株。
1.2.3 点突变菌株HT66PM的构建 利用引物LF(Eco R I)-LR(Xho I)从诱变高产株P3基因组中利用PCR扩增出长达861 bp的基因片段,通过酶切连接获得pK18CP质粒。剩余操作同1.2.2,最后通过测序筛选得到luxR_19(G85A)点突变菌株HT66PM。
1.2.4 生长曲线的测定 挑取单菌落接种到5 mL LB液体培养基中,28℃,180 r/min过夜培养。利用分光光度计进行检测,检测波长为600 nm。将细菌悬浮液接种于60 mL KB培养基中,确保初始OD600=0.01,28℃,180 r/min培养。每株菌3个平行。
1.2.5 PCN产量的检测 用6 mol/L的HCl将发酵液调至pH2.0,加入9倍体积的乙酸乙酯,充分振荡混匀,6 000 r/min离心5 min分层,PCN转移至有机相中。取上层萃取液于通风橱中,使有机溶剂挥发干净。取适量乙腈溶解萃取物,HPLC检测发酵液中PCN产量。检测条件:流动相为乙腈-25 mmol/L乙酸铵,色谱柱为WondaSil C18-WR 反相柱(5 μm;4.6×250 mm,Shimadzu,Japan),检测波长254 nm,检测条件:0-2 min,8%乙腈-92% 25 mmol/L乙酸铵,2-20 min,乙腈浓度从8%升至60%,乙酸铵浓度从92%下降至40%,20-21 min,8%乙腈-92% 25 mmol/L乙酸铵。柱温30℃,流速1.0 mL/min。
1.2.6 泳动性(Swimming)和从动性(Swarming)的检测 挑取单菌落接种于LB液体培养基中28℃,180 r/min过夜培养。将细菌悬浮液稀释至OD600=0.1,取2 μL添加于置于平板中央的无菌滤纸片。28℃培养24 h。
1.2.7 qRT-PCR 实验 由上海桑尼生物科技有限公司完成,所用引物如表格3所示。内参基因为16S rRNA,以ΔΔCt值的方法分析基因的相对表达量。分别检测了HT66以及HT66CL两株菌的rpeA、phzR、phzI、phzE、rpoS、rpoD、psrA、gacA、gacS以及rsmE基因的相对表达量变化情况。
2 结果
2.1 HT66菌株中luxR_19基因的比对
将luxR_19基因与NCBI数据库中的序列比对,结果(表3和图1)显示,luxR_19基因为转录调控因子,属于LuxR家族。比对还表明luxR_19基因与Pseudomonas protegens CHA0、Pseudomonas sp. UW 4、Pseudomonas fluorescens Pf0-1三株菌的luxR序列相似度较高,在核苷酸水平达到了80%以上,而与Pseudomonas putida KT2440和Burkholderia cenocepacia J2315相比,相似度仅为69%。进一步的氨基酸序列分析结果显示,在第85位氨基酸前后的10-20个氨基酸序列均具有很高的同源性,高度保守,属于LuxR_19的活性区域。

表3 luxR_19基因相似性比对

图1 LuxR_19蛋白序列第74-107位氨基酸多序列比对图
2.2 重组菌株HT66CL、HT66PM以及HT66/pME-luxR和HT66CL/pME-luxR的构建
成功构建了pK18CL与pK18PM质粒,利用热激转化导入S17菌株中,之后再通过接合转移导入HT66野生型中,经过两轮同源重组,最终提取基因组,PCR扩增后送上海华大基因公司测序,结果表明成功构建了重组菌株HT66CL和HT66PM。
构建pME-luxR质粒,利用电击转化分别导入HT66以及HT66CL菌株中,从含有四环素抗性的平板上挑取单克隆,提取质粒进行PCR,PCR扩增结果经测序表明HT66/pME-luxR和HT66CL/pME-luxR构建成功。
2.3 luxR_19基因敲除株HT66CL的生长影响
以HT66野生株为对照,同时测定luxR_19基因敲除株HT66CL与HT66WT生长曲线,在28℃,180 r/min的条件下检测48 h。结果(图2)显示,HT66CL与HT66的生长情况十分相似,6 h以前处于适应期,约6 h进入对数期,到24 h以后,HT66CL与HT66均进入稳定期。由此可以推断,luxR_19基因的敲除对于HT66菌体生长情况而言,无较大影响。
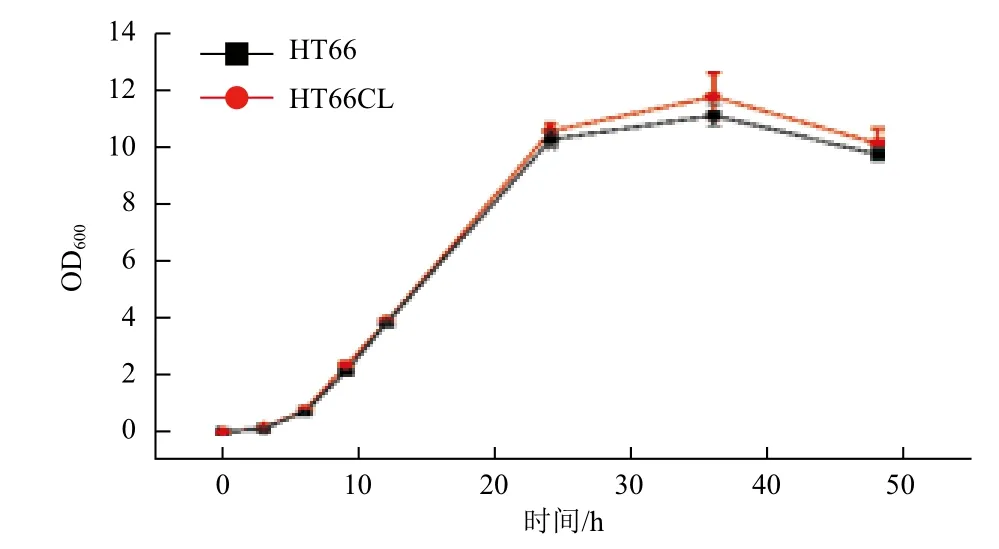
图2 HT66与HT66CL生长曲线测定
同时,考察了HT66CL菌株的泳动性(Swimming)以及从动性(Swarming)变化。28℃条件下培养24 h,拍照记录菌落大小与形态。Swimming实验结果(图3-A和图3-B)显示,HT66与HT66CL菌株边缘均呈现锯齿状,菌落大小相近,无明显差异。Swarming实验结果(图3-C和3-D)表明,HT66与HT66CL边缘不规则且界限模糊,HT66CL形成的生长圈大小明显大于HT66。luxR_19基因的敲除未对HT66的泳动能力产生较大影响,可能是由于其基因本身与控制泳动能力相关的基因无太多作用;luxR_19基因的敲除导致HT66从动能力的增加,说明其在一定程度上能够影响或者控制HT66的从动能力。
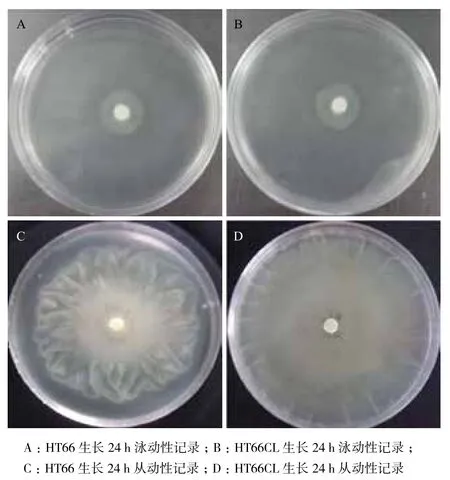
图3 HT66与HT66CL的泳动性及从动性
2.4 luxR_19基因对PCN产量的影响

图4 各菌株发酵24 h后的PCN产量
将HT66、HT66CL以及HT66PM菌株发酵至36 h,萃取发酵液,利用HPLC检测PCN产量。结果(图4)显示,野生型HT66菌株发酵至24 h,PCN产量为402.8 mg/L和473.5 mg/L。HT66CL发酵至24h,PCN产量为153.7 mg/L和198.9 mg/L。就PCN的产量而言,HT66CL比HT66下降了约50%。反观HT66PM,24 h的PCN产量为387.3 mg/L,整体而言虽然比HT66略有下降,但是下降幅度不明显,点突变后无明显变化。
将 HT66、HT66/pME、HT66/pME-luxR、HT66CL、HT66CL/pME以及HT66CL/pME-luxR菌株接种于大瓶KB培养基中发酵,培养至OD600=0.1,添加终浓度为1 mmol/L的IPTG,发酵至24 h,萃取发酵液,利用HPLC检测PCN产量,结果如图4所示。首先是luxR_19的回补:HT66CL/pME-luxR在24 h PCN产量为475.3 mg/L,与HT66CL的153.4 mg/L相比有明显回升;较HT66的405.2 mg/L略高,说明通过luxR_19基因的回补,其产吩嗪能力回复到野生型;同时,HT66CL/pME-luxR的吩嗪产量明显高于HT66/pME的203.5 mg/L,是HT66/pME的2倍;一般来说,由于pME6032质粒较大,可能对菌株的生长带来一定影响,HT66CL/pME的PCN产量最低,仅有103.5 mg/L。然后是luxR_19基因的过表达,HT66/pME-luxR的PCN产量达到了623.7 mg/L,较HT66提高了约50%,较HT66/pME提高了2倍。
2.5 利用qRT-PCR检测相关基因表达水平
根据之前研究和文献报道,选取了参与吩嗪合成和调控吩嗪合成的重要基因,包括双元调控基因gacS/gacA,全局性调控基因rpeA、psrA、rsmE、rpoS,群体感应调控基因phzI/phzR,吩嗪合成必需基因phzE等进行qRT-PCR实验,以研究luxR_19基因的敲除对其产生的影响。以HT66野生型为参照对象,基因表达量的变化如表4所示。

表4 HT66菌株及其基因敲除株中基因表达量变化
表4显示,在HT66CL菌株中,直接参与吩嗪合成的基因phzI/phzR以及phzE基因,其表达水平都有一定的降低,其中phzI为野生型的0.822,phzR为野生型的0.739,phzE的表达量较phzI与phzR而言下降的较少,为野生型的0.901倍。这一结果与发酵结果,即PCN产量有一定下降所一致,从分子层面给上验证了luxR_19基因的敲除会导致吩嗪合成的降低。
从相关调控基因而言,HT66CL菌株的gacA的表达量显著降低,仅为野生型的0.593倍;而gacS的表达量基本维持在稳定的水平,为野生型的1.170倍。rpeA的表达量显著升高,为野生型的1.894倍,根据目前报道,rpeA能够在一定程度上抑制吩嗪类产物的合成[13],与本结果一致。对于两个sigma因子而言,rpoS的表达量保持稳定,rpoD有一定的下降,均与本研究的发酵结果一致。luxR_19基因的敲除导致rsmE与psrA基因表达量下降,分别为野生型的0.367与0.759,但根据相关文献,这两个基因均属于负调控基因。结果表明luxR_19能够影响多个基因的表达,应属于全局性调控基因。
3 讨论
根际促生菌株P. chlororaphis HT66是一株高产PCN的菌株,以HT66作为出发菌株,通过前期的菌种诱变,我们得到了PCN高产菌株P3,基于对突变株的全基因组比较分析,发现luxR_19发生了点突变。通过基因序列的比对,我们发现luxR_19在假单胞菌中具有较高同源性,进一步分析氨基酸序列,发现第85位氨基酸序列附近的氨基酸序列高度保守,处于luxR_19的活性区域。于是我们通过一系列的分子操作研究了luxR_19基因对其生长及吩嗪产量方面的影响。同时,利用点突变株HT66PM验证第85位氨基酸的改变对吩嗪产量的可能影响。
由于次级代谢产物的合成在很大程度上会受到菌株生长情况的影响,LuxR家族调控因子对于菌体生长的调控是多方面的,如通过控制群体感应,调节生物的从动能力以增加生物的环境适应性,如ppoR,通过调控相关代谢产物的合成以增加其环境竞争力等[14]。因此在将luxR_19基因敲除后,我们首先对其进行了生理生化能力的检测,包括测定生长曲线,菌体泳动性以及群体从动性3个指标。结果表明luxR_19基因的敲除不影响菌体的生长状况,但是会在一定程度上影响其群体从动性。在后期的工作中,可以尝试通过对该基因进行突变,从而达到增加群体从动性,确保菌株能够在自然环境中更有竞争力地定植于植物根部,从而促进合成PCN,更好地进行根际促生作用。
luxR_19基因敲除后,PCN产量有一定下降,在对其进行回补后,产量回到了正常水平,将该基因进行过表达后,产量较野生菌株有很大的提升。说明该基因确实对PCN的产量有一定影响,且对于PCN的合成起正调控作用。通过qRT-PCR实验,我们检测了与PCN合成相关基因的表达水平,发现luxR_19基因的缺失直接导致gacA基因表达量明显降低,rpeA负调控因子表达量上升,并在一定程度上导致phz基因簇的表达情况。据此推测luxR_19对于PCN合成的调控作用可能是通过Gac系统来进行的,但具体的调控机制还有待进一步的研究。
通过点突变菌株的构建发现,该位点的点突变与P3菌株的高产无直接联系,也说明化学诱变株高产原因的复杂性。但无论是通过本实验或是相关报道,我们都能发现LuxR家族转录调控因子确实都参与到次级代谢产物的调控中。后期工作将继续分析鉴定P3菌株高产的影响位点,阐明P3菌株高产PCN的机制。
虽然HT66/pME-luxR较HT66而言,吩嗪产量仅仅提高了50%,但我们认为这是由于实验所选用的载体pME6032质粒较大,其本身会对菌体的生长带来一定影响。对比HT66/pME-luxR与HT66/pME两株菌的吩嗪产量,可以发现前者是后者的两倍,这足以展现luxR_19这一基因在提升吩嗪产量上的潜在能力。
在后续的实验中,我们可以通过以下两个方面来提升luxR_19在促进吩嗪生成方面的作用:选用拷贝数更加高的载体,减小对细菌生长的影响;对该基因作逐步的片段分析,发现其作用位点,从而直接在基因组上进行修饰,解除对载体的依赖,构建吩嗪高产菌株。同时,我们将对诱变株的高产原因作继续研究,揭示其高产机理。
4 结论
在Pseudomonas chlororaphis HT66菌株中:luxR_ 19对PCN的合成是正调控因子;luxR_19对细菌从动性有一定影响,而对菌体生长及泳动性无较大影响;诱变高产株P3的PCN产量提高不是由于luxR_19的碱基点突变造成。
[1]张锋华, 许煜泉, 张雪洪.农用杀菌剂吩嗪-1-羧酸的生物合成与基因调控[J].农药研究与应用, 2006(5):4-7.
[2]张云, 胡洪波, 彭华松, 等.产PCA基因工程菌M18G反复补料分批培养研究[J].工业微生物, 2007:16-20.
[3]Han SH, Lee SJ, Moon JH, et al. GacS-dependent production of 2R, 3R-butanediol by Pseudomonas chlororaphis O6 is a major determ inant for eliciting systemic resistance against Erwinia carotovora but not against Pseudomonas syringae pv. tabaci in tobacco[J]. Mol Plant-Microbe Interact, 2006, 19(8):924-930.
[4]Chin-A-Woeng TF, Bloemberg GV, van der Bij AJ, et al. Biocontrol by phenazine-1-carboxamide-producing Pseudomonas chlororaphis PCL1391 of tomato root rot caused by Fusarium oxysporum f. sp. radicis-lycopersici[J]. Mol Plant-Microbe Interact, 1998, 11(11):1069-1077.
[5]Liu H, He Y, Jiang H, et al. Characterization of a phenazineproducing strain Pseudomonas chlororaphis GP72 with broadspectrum antifungal activity from green pepper rhizosphere[J]. Curr Microbiol, 2007, 54(4):302-306.
[6]Shen X, Hu H, Peng H, et al. Comparative genomic analysis of four representative plant growth-promoting rhizobacteria in Pseudomonas[J]. BMC Genomics, 2013, 14(1):271.
[7]Loper JE, Hassan KA, Mavrodi DV, et al. Comparative genomics of plant-associated Pseudomonas spp.:insights into diversity and inheritance of traits involved in multitrophic interactions[J]. PLoS Genet, 2012, 8(7):e1002784.
[8]Park J, Oh S, Anderson A, et al. Production of the antifungal compounds phenazine and pyrrolnitrin from Pseudomonas chlororaphis O6 is differentially regulated by glucose[J]. Lett Appl Microbiol, 2011, 52(5):532-537.
[9]Maddula V, Pierson E, Pierson L. Altering the ratio of phenazines in Pseudomonas chlororaphis(aureofaciens)strain 30-84:effects on biofilm formation and pathogen inhibition[J]. J Bacteriol, 2008,190(8):2759-2766.
[10]Huang L, Chen MM, Wang W, et al. Enhanced production of 2-hydroxyphenazine in Pseudomonas chlororaphis GP72[J]. Appl Microbiol Biotechnol, 2011, 89(1):169-177.
[11]张平原, 彭华松, 张雪洪.高产吩嗪-1-甲酰胺的绿针假单胞菌的诱变与基因工程育种[J].上海交通大学学报:农业科学版, 2015, 33(2):90-94.
[12]He W, Lei J, Jiu Y, et al. The LuxR family members gdmri and gdm rii are positive regulators of geldanamycin biosynthesis in Streptomyces hygroscopicus 17997[J]. Arch Microbiol, 2008,189(5):501-510.
[13]Whistler CA, Pierson III LS. Repression of phenazine antibiotic p roduction in Pseudomonas aureofaciens strain 30-84 by RpeA[J]. J Bacteriol, 2003, 185(13):3718-3725.
[14]Fernández-piñar R, Cámara M, Soriano MI, et al. Ppor, an orphan luxr-family protein of Pseudomonas putida KT2440,modulates competitive fitness and surface motility independently of n-acylhomoserine lactones[J]. Environmental Microbiology Reports, 2011, 3(1):79-85.
(责任编辑 马鑫)
The Effects of luxR_19 on the Synthesis of Phenzaine-1-carboxam ide in Pseudomonas chlororaphis HT66
Chen Yawen Peng Huasong Wang Wei Zhang Xuehong
(State Key Laboratory of Microbial Metabolism,School of Life Sciences and Biotechnology,Shanghai Jiao Tong University,Shanghai200240)
In order to study the potential effects of regulator gene luxR_19 on the synthesis of antibacterial substance phenazine-1-carboxamide(PCN)and physiological-biochemical characteristics in Pseudomonas chlororaphis HT66, strains of gene luxR_19-knockout,-complementation, -overexpression and -point-mutation were constructed, and then the growth curve, fermentation production of PCN, swimming and swarming activities were tested at the target strains. The results showed that the knockout of luxR_19 lowered the production of PCN and the over-expression of gene luxR_19 increased the double production of PCN compared to the wild type;luxR_19 gene affected the expression of several genes relating to the regulation or synthesis of phenazines. Besides, the knockout of luxR_19 had no effects on the growth and swimming activity, however, promoted the swarming activity of HT66. The production of PCN in point-mutation strain of luxR_19(G85A)was similar to that in wild type. The above results indicated that luxR_19 was the positive regulator of PCN production, and could affect the swarming activity of HT66. Besides, the point-mutation at 254thsite in luxR_19 did not affect the production of PCN.
Pseudomonas chlororaphis;HT66;phenazine-1-carboxamide;transcriptional regulator of LuxR family
10.13560/j.cnki.biotech.bull.1985.2015.08.024
2014-12-08
国家自然科学基金项目(31270084),国家“863”计划项目(2012AA022107)
陈亚汶,女,硕士研究生,研究方向:微生物代谢调控;E-mail:moda_yaya@hotmail.com
张雪洪,男,博士,教授,研究方向:微生物代谢调控;E-mail:xuehzhang@sjtu.edu.cn
猜你喜欢
河北农机(2020年10期)2020-12-14
今日农业(2019年15期)2019-01-03
食品科学(2018年10期)2018-05-23
中成药(2017年9期)2017-12-19
中华老年口腔医学杂志(2016年2期)2017-01-15
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
现代检验医学杂志(2015年1期)2015-02-06
西南军医(2015年6期)2015-01-23
食品工业科技(2014年23期)2014-03-11
