
5个凡纳滨对虾抗WSSV家系遗传性状的比较
2015-08-30 08:55:35王成桂杨世平蔡志方黄海立梁华芳陈兆明孙成波
浙江海洋大学学报(自然科学版) 2015年3期
王成桂,杨世平,蔡志方,黄海立,梁华芳,陈兆明,孙成波
(广东海洋大学水产学院,广东湛江 524088)
5个凡纳滨对虾抗WSSV家系遗传性状的比较
王成桂,杨世平,蔡志方,黄海立,梁华芳,陈兆明,孙成波
(广东海洋大学水产学院,广东湛江524088)
通过WSSV感染后存活的凡纳滨对虾作为亲本,培育9个抗WSSV家系,比较其中5个家系生长、抗盐度突变、抗氨氮和亚硝酸盐浓度突变、抗WSSV的性状,研究凡纳滨对虾生长、抗逆和抗WSSV的遗传关系。结果表明:各抗WSSV家系的生长有显著差异(P<0.05),K8生长最快,平均体长和体质量为12.44 cm、25.78 g,K3最慢,平均体长和体质量为9.06 cm、9.45 g;抗盐度突变能力存在显著差异(P<0.05),K8的存活率最高,为90%,显著高于对照组Gjk-F1·1、Klw-F1和Gjk-F1·2(P<0.05),K4存活率最低,为56.67%;各家系抗氨氮浓度突变能力差异显著(P<0.05),K6存活率最高,为96.67%,显著高于对照组Sis-F1(P<0.05),K8存活率最低,为76.67%;各家系抗亚硝酸盐浓度突变能力差异显著(P<0.05),K2存活率最高,为96.67%,显著高于所有对照组(P<0.05),K4存活率最低,为36.67%;各家系抗WSSV能力差异显著(P<0.05),K6、K3和K4差异不显著(P>0.05),但显著高于K2和K8(P<0.05),存活率分别为33.33%、30.00%和30.00%,K2和K8差异不显著(P>0.05),存活率为16.67%和10.00%。因此,从单一性状考虑,K8可以作为高生长或者高抗盐度突变的选育材料,K6为高抗氨氮浓度突变或者抗WSSV选育材料,K2为高抗亚硝酸盐浓度突变的选育材料;综合多性状因素考虑,仅有K6可以选为优良抗WSSV家系的选育材料。研究为凡纳滨对虾的抗WSSV家系的选育提供一定的数据支持。
凡纳滨对虾;家系;抗WSSV;遗传性状
凡纳滨对虾Litopenaeus vannamei又称南美白对虾,自然分布于秘鲁至墨西哥的太平洋沿岸[1],自20世纪80年代末由中科院海洋研究所引进我国养殖[2],其生长快、抗病抗逆力强、出肉率高、适合低盐度养殖等优点[3],已在全国沿海及部分内陆地区大面积养殖,亦是目前全球主要养殖的对虾品种之一[4]。经过多年的累代养殖、近亲繁殖等因素对虾种质资源退化,对虾养殖业日益受病害的困扰[5-9],养殖产量急剧下降,2013年全球养殖对虾产量预计较2012年减少23%[10]。尤其WSSV感染和死亡率极高,1995年10月,西半球报道了第一例对虾WSS[11],至今WSS已是全球对虾养殖业面临的主要的病害,近年来,国内外学者已进行了许多关于WSS的研究[12-15],但由于WSSV感染途径广,防治仍然困难,效仿IO选育抗TSV方法[16]进行抗WSSV家系选育不妨为一种有效的针对措施。
目前,我国养殖的凡纳滨对虾亲虾主要来源为SIS、科拿湾及正大等国外育种公司,通过正规途径进口到国内的亲虾超过20万对,并且每年都在增加[17],进口亲虾花费大量外汇,但近年质量并不稳定[18],同时种质资源受制于人,优良亲虾急需国产化。快速生长、高抗逆抗病能力等特性是对虾遗传选育的主要选择指数[19],国内外已开展较多的研究。1995-1998年,美国国家农业部(USDA)和海洋研究所(OI)开展基于凡纳滨对虾生长性能和桃拉综合病毒(Taura Syndrome Virus,TSV)抗性等权重的综合选择指数进行选择育种[18];王专伟等[19]开展15个斑节对虾家系生长及WSSV分析;纪荣兴等[21]研究日本对虾抗WSSV子三代的抗病性能;黄永春等[22-23]研究凡纳滨选育家系抗病、生长及生长特性。本实验采用WSSV攻毒后存活的对虾作为亲虾,培育的抗WSSV家系,比较其生长、抗盐度突变、抗氨氮和亚硝酸盐浓度突变、抗WSSV等遗传性状,以期为凡纳滨对虾遗传选育提供依据及参考。
1 材料与方法
1.1材料
2011年8月随机挑选广东海洋大学东海岛海洋生物研究基地选育的凡纳滨对虾1个家系42 850尾幼虾,平均体长4~7 cm,投喂感染WSSV的病死虾,11 d后,剩余288尾,停止攻毒,换池后,以剩余对虾作为抗WSSV基础群体养殖。2012年7月15日至10月31日,经催熟、交配、产卵、孵化、育苗等程序培育9 个F1代不同胞抗WSSV家系,分别标记为K1~K9,每个家系随机取6 000尾虾苗移至室外50 m2水泥池养殖,养殖池盐度为25~30。2013年6月10日至2013年12月21日选择生长性状较好的5个抗WSSV家系,分别为K2、K3、K4、K6、K8作为实验材料,并以进口亲虾子一代作为对照组,分别记为Gjk、Sis、Klw。
1.2方法
1.2.1生长比较
抗WSSV家系养殖45 d后,密度调整为1 000尾/池,100 d后调整为600尾/池。每个家系用抛网随机取虾30尾,按照吴立峰等[24]方法,吸干水份后用电子天平(JCS-300 g)测量体质量,数据精确到0.01 g,用游标卡尺测量体长,数据精确到0.01 cm。
1.2.2抗盐度突变能力比较
每个家系和对照组设3个平行组,每组取30尾,实验虾预先在盐度为30海水中暂养2 d,在室内150 L塑料白桶中进行,水体30 L,自然光照,温度32~34℃,pH为8.0~8.3,接照杨海朋等[25]方法将实验虾直接从盐度为30的暂养桶移到盐度为5的白桶中,每隔4 h统计存活情况。
1.2.3抗氨氮浓度突变能力比较
按照胡贤德等方法[26]以相同规格SIS亲虾F1代做预备实验计算半致死浓度,96h半致死浓度为31.67mg/L,采用氯化铵(分析纯)调节实验海水氨氮浓度至此值,实验桶为150 L塑料桶,放置实验海水30 L,将实验虾直接从暂养桶移到实验桶,每隔4 h统计存活率,并采用简易比色盒检测和调整水体中的氨氮浓度,试验时间为96 h。每个家系和对照组进行3次实验,每次取虾30尾。
1.2.4抗亚硝酸盐浓度突变能力比较
按照胡贤德等方法[27]以相同规格SIS亲虾F1代做预备实验计算半致死浓度,96 h半致死浓度为50.51 mg/L,采用亚硝酸钠(分析纯)调节实验海水亚硝酸盐浓度至此值,实验桶为150 L塑料桶,放置实验海水30 L,将实验虾直接从暂养桶移到实验桶,每隔4 h统计存活率,并采用简易比色盒检测和调整水体中的亚硝酸盐浓度,试验时间为96 h。每个家系和对照组进行3次实验,每次取虾30尾。
1.2.5抗WSSV能力比较
每个家系设3个平行组,每组取虾30尾,实验桶为150 L,水体30 L,投喂感染WSSV的病死虾肌肉,实验10 d,每天观察并及时剔除死亡个体,统计存活率。
1.3数据处理
对于测量和计算的数据采用SPSS11.5软件进行单因素方差分析,有显著差异的再作Duncan多重比较,以P<0.05作为差异显著水平。
2 结果
2.1生长比较
不同家系的平均体长和平均体质量存在显著性差异(P<0.05)。平均体长最长的为K8,为12.44 cm,其次为K2,为11.57 cm,K3的平均体长最短,为9.06 cm,比K8短27.17%;平均体质量最大的为K8,为25.78g,其次为K2,为18.06g,K3的平均体质量最小,为9.45 g,比K8轻63.34%(表1)。

表1 抗WSSV家系生长比较Tab.1 Comparison on growth of anti WSSV families
2.2抗盐度突变能力比较
不同家系抗盐度突变能力存在显著差异(P<0.05)。K4的存活率最低,为56.67%,显著低于其它实验组及对照组Gjk-F1·1、Sis-F1、Gjk-F1·2(P<0.05),K4和对照组Klw-F1差异不显著(P>0.05);其次为K6,存活率为73.33%,显著低于对照组Sis-F1(P<0.05);实验组K8的存活率显者高于其它实验组(P<0.05),和对照组Sis-F1差异不显著(P>0.05),但显著高于其它对照组(P<0.05)(表2)。

表2 抗盐度突变能力比较Tab.2 Comparison on ability of enduring salinity change
2.3抗氨氮浓度突变能力比较
不同家系抗氨氮浓度突变能力存在显著差异(P<0.05)。K8、K4和K2存活率差异不显著(P>0.05),分别为76.67%、83.33%和86.67%,显著低于其它实验组和对照组Gjk-F1·1、Klw-F1、Gjk-F1·2(P<0.05),K4和K2存活率显著高于对照组Sis-F1(P<0.05);K3和K6存活率差异不显著(P>0.05),分别为93.33%和96.67%,和对照组Gjk-F1·1、Klw-F1、Gjk-F1·2差异不显著(P>0.05)(表3)。
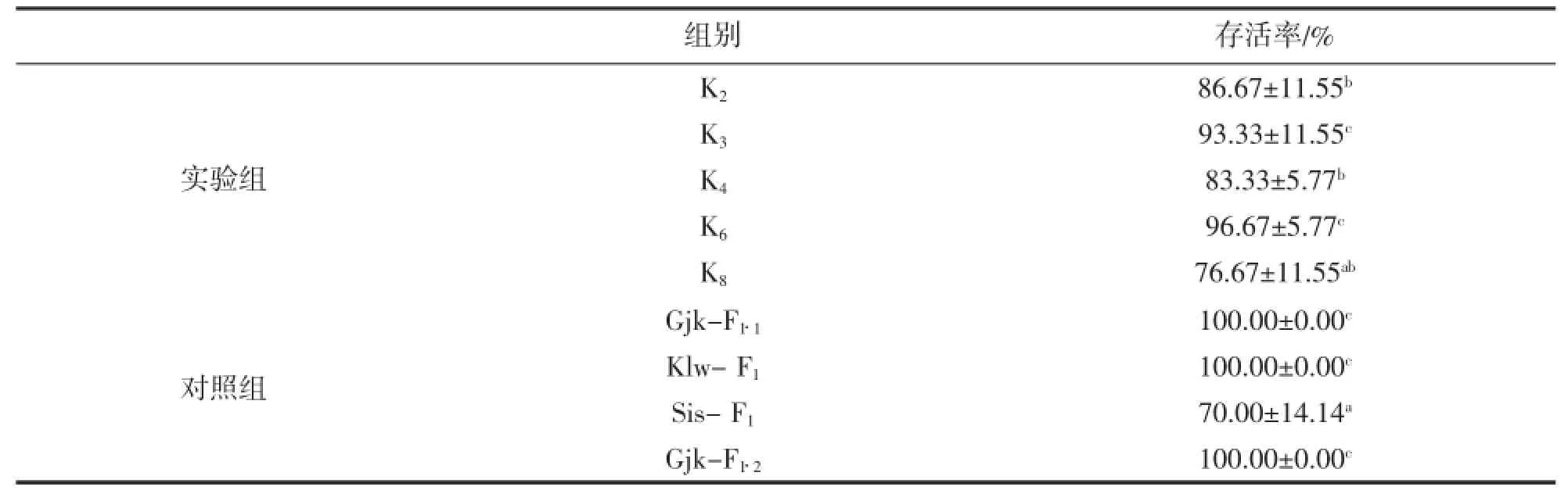
表3 抗氨氮浓度突变能力比较Tab.3 Comparison on ability of enduring ammonia nitrogen density change
2.4抗亚硝酸盐浓度突变能力比较
不同家系抗亚硝酸盐浓度突变能力存在显著差异(P<0.05)。K4存活率最低,为36.67%,显著低于其它家系(P<0.05),和对照组Sis-F1差异不显著(P>0.05),但显著低于其它对照组(P<0.05);K6和K3存活率差异不显著(P>0.05),分别为80.00%和83.33%,和对照组Gjk-F1·1差异不显著(P>0.05),显著高于对照组Gjk-F1·2和Klw-F1(P<0.05),显著低于K8和K2(P<0.05);K2存活率显著高于其它实验组和所有对照组(P<0.05),为96.67%,其次为K8,存活率为90.00%,显著高于所有对照组(P<0.05)(表4)。
2.5抗WSSV能力比较
不同家系抗WSSV能力存在显著性差异(P<0.05)。K8和K2存活率差异不显著(P>0.05),分别为10.00%和16.67%,显著低于K6、K3、K4(P<0.05),K6、K3、K4存活率差异不显著(P>0.05),分别为33.33%、30.00%和30.00%(表5)。
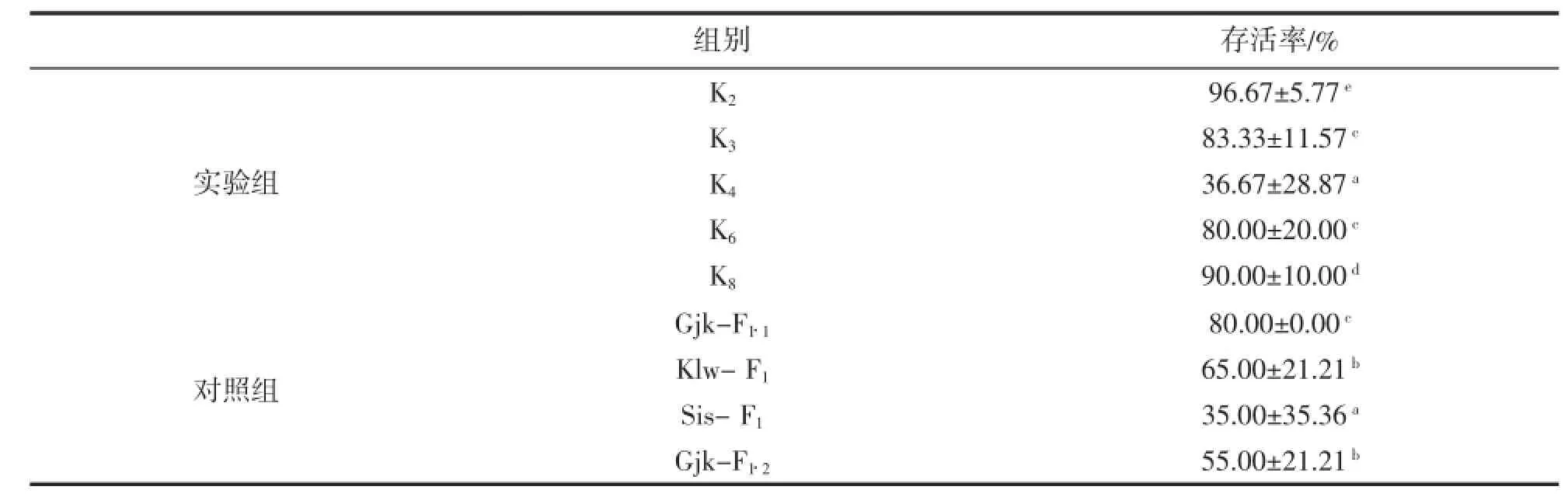
表4 抗亚硝酸盐浓度突变能力比较Tab.4 Comparison on ability of enduring nitrite density change
3 讨论
养殖对虾生长速度受环境、管理水平、遗传因素等影响,有学者已证实生长性状是可以遗传的[28-31]。本实验各家系的生长速度差异显著(P<0.05),K8的体质量是K3的2.72倍,各家系在相同的环境和一样的管理水平下养殖,生长速度主要受遗传因素影响,由于各家系父代为全同胞家系,生长速度至F2代出现分化,表明和生长相关的优良的遗传性状可以进一步通过杂交、自交等方式稳定下来。
盐度是对虾养殖中重要的环境因子之一,影响对虾存活、生长、渗透压和酶活性等功能[32],抗盐度突变能力是对虾抗逆能力主要参考指标,甚至有养殖者试苗时通过计算虾苗在不同盐度海水的存活率来检验对虾的抗逆性状[33]。本实验各家系抗盐度突变能力差异显著(P<0.05),和林红军等研究结果类似。K8的存活率最高,接近目前亲虾主要来源SIS品种,可以作为对虾淡化养殖品种的选育材料。
养殖环境恶化后,氨氮和亚硝酸盐浓度会升高,在高密度养殖凡纳滨对虾的池塘中后期投饵增加及清底不及时容易发生。向赟等[34]研究表明对于携带WSSV的对虾,氨氮变化越大,WSSV从潜伏感染转为急性感染的可能越大。本实验,各家系抗氨氮和亚硝酸盐浓度突变能力比对照组SIS强,K6、K2可以分别造作抗氨氮和亚硝酸浓度突变的选育家系。由于本实验设计针对抗WSSV家系的选育,养殖家系父本时,经常换水和清底,氨氮和亚硝酸盐浓度控制在安全范围以内,父本没有经受氨氮和亚硝酸盐浓度突变的自然选择,各家系抗氨氮能力低于对照组Gjk-F1·1、Klw-F1、Gjk-F1·2,并没有表现较强的抗性,这与没有经过抗氨氮选育有关。K8抗氨氮能力差,抗WSSV力亦较差,与向赟研究结果相类似。K2、K3、K8的抗亚硝酸盐浓度突变能力高于所有对照组,表明具有高抗亚硝酸盐性能,至于是否与遗传因素相关,受本实验条件限制,亦不能作出结论,须进一步实验证实。
WSSV是对对虾危害非常严重的病原[35],对虾发病后3~10 d就会100%死亡[9]。本实验材料父本经WSSV感染后,存活率仅为0.67%,剩余父本具有高抗WSSV的性状,杂交后遗传至实验家系。实验结果显示,杂交后的各家系存活率都高过上一代,抗WSSV遗传性状得到提高。各家系存活率差异显著(P<0.05),与王专伟等[20]研究结果类似。存活率最高的K6,仅为33.33%,不具备生产意义,需要作一步抗WSSV选育。但本实验家系建立的方式是以经感染WSSV的虾作为亲本,WSSV经繁殖程序传至子代,在对虾育苗上不符合生产SPF对虾要求,在生产上亦不能应用,但此选育方式效果直接,相对于采用SPF对虾作亲本再对子代进行攻毒实验,随机性少,选择强度大。
不同家系生长性状与抗逆、抗WSSV能力不同,选育必须按照不同指标权重进行选择,寻找满足多性状选育的最佳组合,但许多研究表明各选育指标并不呈最理想线型呈现。张吕平等[36]发现生长速度和体长变异系数差异不明显;陈锚等[37]发现生长和畸形率无显著性差异结果表明,生长较快的家系与生长较慢的家系畸形率统计上并无明显差异;MOSS等[38]1999年报道,514个全同胞家系的生产性能与TSV感染的家系存活率呈负相关(r=-0.45)。本实验结果和上述实验相类似,发现生长性能与抗逆、抗WSSV能力无明显的相关关系,本实验中,K8生长最高,抗盐度突变能力亦最强,但抗WSSV能力最差,本实验主要以抗WSSV能力作为选育指标,因此K8不能作为下一步选育的备材料。K3生长较差,K2抗WSSV能力较差,K4抗亚硝酸盐浓度能力最差,这3个家系亦不能作为选育家系。从多性状选育分析,考虑选育综合指数要求[19],K6具有高抗WSSV性状,生长、抗逆等指标亦较强,可以作为抗WSSV家系选育材料作进一步选育。
[1]张伟权.世界重要养殖品种--南美白对虾生物学简介[J].海洋科学,1993(3):69-73.
[2]张乃禹.我国对虾产业化养殖核心技术-“中国对虾工厂化育苗技术的研究”攻关回眸[J].海洋科学,2012,36(11):114-117.
[3]吴琴瑟.对虾健康养殖大全[M].北京:中国农业出版社,2007:125-129.
[4]崔和,肖乐.2011~2012我国对虾产业发展现状及展望[J].中国水产,2012(7):25-26.
[5]王云祥,李秀梅,孙金生,等.天津地区对虾爆发性流行病的病因、病理及传播途径的初步研究[J].海洋科学,1994(6):1-5.
[6]苏永全,蔡心一,王军.1992-1993年福建南部地区虾病的调研[J].海洋科学,1995(1):1-4.
[7]蔡生力,黄倢,王崇明,等.1993-1994对虾暴发病的流行病学研究[J].水产学报,1995,19(2):112-119.
[8]黄倢,蔡生力,宋晓玲,等.对虾暴发性流行病病原的人工感染研究[J].海洋水产研究,1995,16(1):51-58.
[9]胡超群.广东沿海养殖对虾疾病流行特点及病因[J].海洋科学,1994(6):9-10.
[10]国家虾产业技术体系等编著.对虾肝胰腺坏死症和白斑综合症防控技术手册[Z].2014:1-16.
[11]孙成波,李婷,王平,等.高位池养殖对虾携带白斑综合症病毒变化[J].海洋通报,2009,28(2):116-120.
[12]王吉桥,姜玉声,张建.白斑综合征病毒(White spot syndrome virus)的形态和分子生物学[J].现代渔业信息,2008,23 (11):3-6.
[13]LIGHTNER D V.A Handbook of Shrimp Pathology and Diagnostic Procedures for Disease of Cultured Penaeid Shrimp[M]// Sec-tion 3:Viruses World Aquaculture.Lovisiana:Boton Rouqe,1996.
[14]江世贵,何建国,吕玲,等.白斑综合症对斑节对虾亲虾的感染及垂直传播的初步研究[J].中山大学学报:自然科学版,2000,39(增刊):164-171.
[15]何建国,周化民,姚泊,等.白斑综合症杆状病毒的感染途径和宿主种类[J].中山大学学报:自然科学版,1999,38(2):65-69.
[16]ARGUE B J,ARCE S M,LOTZ J M,et al.Selective breeding of Pacific white shrimp(Litopenaeus vannamei)for growth and resistance to Taura Syndrome Virus[J].Aquaculture,2002,204(38):447-460.
[17]李建宏.塞班又一亲虾新品牌进入中国市场[J].当代水产,2014(5):40-41.
[18]梁华芳,杜国平,黄海立,等.凡纳滨对虾快速生长家系选育的初步研究[J].广东海洋大学学报,2011,31(3):12-15.
[19]范兆廷.水产动物育种学[M].北京:中国农业出版社,2005:85-98.
[20]王专伟,黄建华,杨其彬.15个斑节对虾家系生长及抗白斑病毒分析[J].海洋科学进展,2011,29(4):521-528.
[21]纪荣兴,邹文政,鄢庆枇.日本对虾抗白斑病子三代的抗白斑综合征病毒感染能力及免疫特性[J].水产学报,2008,32(1):98-104.
[22]黄永春,艾华水,殷志新,等.第四代凡纳滨对虾抗WSSV选育家系的抗病及免疫特性研究 [J].水产学报,2010,34 (10):1 549-1 558.
[23]黄永春,艾华水,殷志新,等.凡纳滨对虾抗WSSV选育家系的抗病与生长特性[J].台湾海峡,2011,30(3):412-418.
[24]吴立峰,张吕平,胡超群,等.2个凡纳滨对虾全同胞家系在不同盐度下的生长比较[J].海洋生物学,2011,30(1):152-158.
[25]杨海朋,胡超群,张吕平,等.凡纳滨对虾家系淡水耐受性状与生长性状的关系[J].海洋生物学,2014,33(4):69-76.
[26]胡贤德,孙成波,蔡鹤翔,等.不同盐度条件下氨氮对斑节对虾的毒性试验[J].广西科学,2009,16(2):206-209.
[27]胡贤德,孙成波,王平,等.不同盐度条件下亚硝酸氮对斑节对虾的毒性影响[J].沈阳农业大学学报,2009,40(1):103-106.
[28]HETZEL D J S,CROCOS P J,DAVIS G P,et al.Response to selection and heritability for growth in the Kuruma prawn Penaeus japonicus[J].Aquaculture,2000,181:215-223.
[29]GOYARD E,PATROIS J,PEIGNON M,et al.Selection for better growth of Penaeus stylirostris in Tahiti and New Caledonia [J].Aquaculture,2002,204:461-468.
[30]FINLEY L M,HALEY L E.The genetic of aggression in the juvenile American lobers,homarus amerucanus[J].Aquaculture,1983,33:135-139.
[31]LESTER L J.Difference in larval growth among families of Penaeus stylirostris Stimpson and P.vannamei Boone[J].Aquaculture Research,1988,19:243-251.
[32]李华,李强,曲健凤,等.不同盐度下凡纳滨对虾血淋巴免疫生理指标比较[J].中国海洋大学学报,2007,37(6):927-930.
[33]林红军,张吕平,沈琪,等.10个凡纳滨对虾全同胞家系淡水耐受性比较[J].海洋湖沼通报,2010(4):143-148.
[34]向赟,王刚,龚永,等.氨氮质量浓度对感染白斑综合症病毒的凡纳滨对虾的影响 [J].热带生物学报,2014,5(3):220-227.
[35]YANG F,HE J,LIN X H,et al.Complete genome sequence of the shrimp white spot bacilliform virus[J].J Virology,2001,75:11 811-11 820.
[36]张吕平,吴立峰,沈琪,等.凡纳滨对虾全同胞家系的建立及生长比较[J].水产学报,2009,33(6):932-938.
[37]陈锚,吴长功,相建海,等.凡纳滨对虾的选育与家系的建立[J].海洋科学,2008,32(11):5-8.
[38]MOSS M S,ARGUE B,ARCE S.Genetic Improvement of the Pacific white shrimp,Litpenaeus vannamei,at the Oceanic Institute[J].The Advocate,1999,12:41-44.
The Compare of Genetic Characters of Five Litopenaeus vannamei Anit WSSV Families
WANG Cheng-gui,YANG Shi-ping,CAI Zhi-fang,et al
(College of Fisheries,Guangdong Ocean University,Zhanjiang524088,China)
Through the WSSV infection survival Litopenaeus vannamei were bred as parents and nine anit WSSV L.vannamei families were bred.Five families were selected to test the growth,the ablity of enduring the change of salinity,ammonia nitrogen and nitrite density for comparison the genetic characters of L.vannamei anti WSSV families.The results indicated there was significant difference on the growth each family(P<0.05),K8grew fastest,the average body length and body weight of K8was 12.44 cm and 25.78 g,K3was the slowest,the average body length and body weight of K3was 9.06 cm and 9.45 g.There was significant difference in the ability of enduring salinity change of each family(P<0.05),the survival rate of K8 was the highest and 90%,which was significant higher than Gjk-F1·1,Klw-F1and Gjk-F1·2as control groups,the survival rate of K4was the lowest and 56.67%.There was significant difference in the ability of enduring ammonia nitrogen density change of each family(P<0.05),the survival rate of K6was the highest and 96.67%,which was significant higher than Sis-F1as the control group(P<0.05),the survival rate of K8was the lowest and 76.67%.There was significant difference in the ability of enduring nitrite density change of each family(P<0.05),the survival rate of K2was the highest and 96.67%,which was significant higher than all control groups(P<0.05),the survival rate of K4was the lowest and 36.67%.There was significant difference in the ability of anti WSSV of each family (P<0.05),the survival rate of K6,K3and K4was not significantly different(P>0.05),but was significang higher than K2and K8(P<0.05),which was 33.33%,30.00%and 30.00%,respectivly.The ability of anti WSSV of K2and K8was not significantly different(P>0.05),was 16.67%and 10.00%.Therefore,considering the single target only,K8can be used as breeding materials for high growth or salinity mutation,K6can be used as breeding materials for ammonia nitrogen density change or anti WSSV,K2can be used as breeding materials for nitrite density change.But consideration of multiple traits,K6can be uesd breeding materials for better anti WSSV families only.The findings provide evidence for the selective breeding of new anti WSSV families of L.vannamei.
Litopenaeus vannamei;families;anti WSSV;genetic characters
S968.22
A
1008-830X(2015)03-0227-06
2014-12-20
广东省海洋渔业科技推广专项(A201301D02);湛江市自主创新企业科技创新项目(2011[79])
王成桂(1978-),男,助理研究员,研究方向:海产动物增养殖.E-mail:longshore@163.com
孙成波(1970-),男,博士,教授,研究方向:对虾养殖和病害防治.E-mail:suncb@gdou.edu.cn
猜你喜欢
当代水产(2022年8期)2022-09-20 06:46:22
当代水产(2022年5期)2022-06-05 07:55:46
当代水产(2021年8期)2021-11-04 08:49:04
当代水产(2021年4期)2021-07-20 08:10:54
当代水产(2019年1期)2019-05-16 02:42:00
广东海洋大学学报(2015年4期)2016-01-13 08:39:32
广东海洋大学学报(2015年4期)2016-01-13 08:39:30
听力学及言语疾病杂志(2015年5期)2015-12-24 01:47:04
首都医科大学学报(2015年4期)2015-12-16 13:00:08
郑州大学学报(医学版)(2015年2期)2015-02-27 14:50:44
