
TLR5在不同非小细胞肺癌细胞株的表达及其活化机制的初步探讨
2015-08-26 12:40周辉罗媚文亦戈马安迪罗永忠易青陈建华肖玲
中国肺癌杂志 2015年1期
周辉 罗媚 文亦戈 马安迪 罗永忠 易青 陈建华 肖玲
肺癌是最常见的恶性肿瘤,发病率高,生存率低,预后差,居恶性肿瘤死亡率首位[1]。非小细胞肺癌(non-small cell lung cancer, NSCLC)约占肺癌的80%-85%,研究[2]发现慢性炎症与肿瘤发生相关。Toll样受体5(toll-like receptor 5, TLR5)是toll样受体家族中的一员,是一类最具有特征性的模式受体分子,能募集多种配体蛋白,激发信号转导,导致一些特异性转录因子的活化。细菌的鞭毛蛋白(Flagellin)作为TLR5特异性的外源性配体,TLR5与其结合后具有保护性的抗炎作用,其可能在肿瘤异常表达。我们前期研究[3]发现,与正常肺组织比较,TLR5在NSCLC组织中高表达,且与吸烟分化程度呈正相关,但有关TLR5在NSCLC高表达后的信号通路活化情况的研究并不多见。本研究旨在探讨不同NSCLC细胞株中TLR5表达情况,寻找较好的实验模型,并探讨其信号通路活化的机制。
1 材料与方法
1.1 材料
1.1.1 细胞株 人肺腺癌细胞株SPC-A-1购于中科院上海细胞研究所;人肺腺癌细胞株A549(cat# CCL-185),人肺鳞癌细胞株SK-MES-1(cat# HTB-58)均购自美国菌种保藏中心(American Type Culture Collection, ATCC)。
1.1.2 主要试剂和耗材 DMEM培养基、FBS、胰酶均购自美国ATCC公司;Triton X-100购自美国Genview公司;反转录试剂盒、PCR Mix试剂盒均购自加拿大Fermentas(MBI Fermentas)分子生物技术公司;Tris-Base、10%SDS均购自美国Sigma公司;TEMED购自上海生工生物工程股份有限公司;天然膜蛋白抽提试剂盒ProteoExtractTM(M-PEK)购自德国MERCK公司(中国);Total protein Extraction试剂盒购自美国ProMab公司;NucBusterTMProtein Exaction Kit购自德国MERCK公司(中国);Mouse GAPDH antibody、Goat Anti Rabbit IgG/HRP购自美国Santa Cruz生物公司;Rabbit Anti Goat IgG/HRP购自中杉金桥技术有限公司(北京);Goat Anti Mouse IgG/HRP、Goat Anti Mouse IgG+A+M(H+L)/HRP均购自美国Zymed Laboratories公司;Goat anti-rat IgG-HRP、Rabbit Anti-Sheep IgG/HRP均购自美国Santa Cruz生物技术公司;NF-κB荧光素酶报告基因质粒购自江苏碧云天技术研究所。shRNA空载体pLKO.3G购自Addgene公司;LipofectamineTM2000试剂、Opti-MEM®低血清培养基均购自美国Invitrogen公司。
1.1.3 引物 见表1。
1.2 方法
1.2.1 TLR5在不同NSCLC细胞系的表达 免疫荧光观察不同NSCLC细胞株TLR5蛋白表达,Trizol法提取细胞的RNA,RT-PCR检测不同肺癌细胞株TLR5 mRNA的水平,并用琼脂糖凝胶电泳检测。通过gel-pro 6.0灰度扫描软件分析各细胞株TLR5的表达状况。Western blot检测TLR5蛋白表达水平。
1.2.2 TLR5与NF-κB信号通路的关系 分别用0 μg/mL、0.01 μg/mL、0.1 μg/mL、1 μg/mL、5 μg/mL、10 μg/mL的鞭毛蛋白刺激,用NF-κB荧光素酶报告基因质粒瞬时转染三种细胞株,检测细胞内NF-κB荧光素酶的活性。选择出TLR5表达最高的细胞株作为研究对象,选择适合浓度的鞭毛蛋白,分别用0 μg/mL、0.01 μg/mL、0.1 μg/mL、1 μg/mL、10 μg/mL的TLR5抗体抑制通路活化,检测细胞内NF-κB荧光素酶的活性,验证TLR5活化通路。
1.2.3 TLR5信号通路磷酸化水平检测 用适合浓度的鞭毛蛋白刺激SPC-A-1细胞,分别在0 min、10 min、30 min和60 min用Western blot方法检测TLR5活化后作用通路中的p-IKBα、IKBα、p-ERK1/2、ERK1/2、p-JNK等分子的变化。构建靶向TLR5基因的shRNA质粒,用脂质体法体外转染SPC-A-1细胞,48 h后用相同浓度鞭毛蛋白刺激,Western blot法在相同的四个时间点检测对TLR5信号通路分子的变化(β-actin作为内参)。
1.3 统计学方法 数据分析采用SPSS 17.0软件(美国)进行,计量数据表示为均数±标准差(Mean±SD)。计量资料使用t检验,计数资料使用χ2检验,P<0.05认为差异有统计学意义。
2 结果
2.1 TLR5在不同NSCLC细胞系的表达 免疫荧光显示TLR5蛋白在肺腺癌细胞株SPC-A-1中呈高表达,且主要表现在细胞膜上,在细胞质中也表达(图1A)。在两种肺腺癌细胞株均可检测到TLR5 mRNA表达(SPC-A-1和A549细胞中TLR5 mRNA相应对表达量分别为0.386±0.022、0.293±0.018),而肺鳞癌细胞株SK-MES-1未检出(P<0.05) (图1B)。Western blot检测结果显示肺腺癌细胞株SPC-A-1和A549中均有TLR5蛋白表达(SPC-A-1和A549细胞中TLR蛋白相对表达量分别为0.446±0.023、0.195±0.011),而且SPC-A-1细胞株中TLR5蛋白的水平明显高于A549细胞株(P<0.05),肺鳞癌细胞株中未检出TLR5蛋白(图1C,图1D)。
2.2 TLR5与NF-κB信号通路的关系(不同浓度Flagellin刺激) 用NF-κB荧光素酶报告基因质粒瞬时转染后,检测细胞内荧光素酶的活性。随着鞭毛蛋白浓度的增加,SPC-A-1细胞内NF-κB荧光素酶的活性增强,且呈浓度依赖性,0.1 μg/mL鞭毛蛋白即可明显增强NF-κB荧光素酶的活性(P<0.05);而在A549和SK-MES-1细胞中,NF-κB荧光素酶的活性变化不如SPC-A-1细胞明显(表2,图2A)。SPC-A-1细胞内NF-κB荧光素酶的活性可被TLR5抗体抑制,随着TLR5抗体浓度的增加,NF-κB荧光素酶的活性明显降低(P<0.05),与TLR5抗体浓度负相关(表3,图2B)。
2.3 TLR5信号通路磷酸化水平检测 选取0.1 μg/mL鞭毛蛋白处理SPC-A-1细胞,分别在作用0 min、10 min、30 min、60 min后,收集细胞进行Western blot检测。结果显示,与0 min相比较,p-IKBα、p-ERK1/2、p-JNK水平在鞭毛蛋白刺激10 min即明显增高,30 min达到高峰,60 min开始下降(P<0.05),且与10 min和60 min相比,p-IKBα、p-ERK1/2、p-JNK水平在30 min明显增高(P<0.05);而IKBα、ERK1/2的水平在0 min、10 min、30 min、60 min均无明显变化(P>0.05)(表4,图3)。
2.4 TLR5-shRNA转染效率及转染后TLR5的表达情况 在转染48 h后,在荧光显微镜下可见空质粒对照组和TLR5-shRNA组被转染SPC-A-1细胞数多,转染效果高。TLR5-shRNA转染SPC-A-1细胞48 h后,与空白对照组(TLR5 蛋白相对表达量为0.382±0.017)相比,TLR5-shRNA组细胞TLR5表达(TLR5 蛋白相对表达量为0.118±0.008)明显受到抑制(图4)。
2.5 Flagellin对TLR5-shRNA SPC-A-1细胞TLR5信号通路的影响 TLR5-shRNA转染SPC-A-1细胞48 h后,加入Flagellin(0.1 μg/mL),用Western blot法分别在0 min、10 min、30 min、60 min检测信号通路相关蛋白的表达。结果显示,在四个不同的作用时间点,IKBα、ERK1/2蛋白的水平无明显变化(P>0.05),p-ERK1/2蛋白水平随着时间延长明显增高(P<0.05),而p-IKBα、p-JNK蛋白均未检出(表5,图5 )。
3 讨论

表1 引物Tab1 Primer
TLRs在机体粘膜免疫、肿瘤起始和发展中发挥重要作用,不仅在免疫细胞上表达,也广泛表达于正常上皮细胞和胃癌等多种肿瘤细胞[4,5]。我们前期实验发现TLR5蛋白在NSCLC组织中表达明显增高,在不同组织学类型的表达阳性率为鳞癌65%、腺癌82.1%、鳞腺癌50%、其他类型20%,其中TLR5在腺癌中表达阳性率最高。为了进一步研究其高表达的意义和作用机制,挑选合适的细胞株作为研究对象,我们分别检测TLR5在两种人肺腺癌细胞株SPC-A-1、A549细胞和人肺鳞癌细胞株SK-MES-1中的表达情况。免疫荧光、RT-PCR和Western blot结果表明在人肺鳞癌细胞株SK-MES-1中未检测到TLR5表达,两种不同NSCLC腺癌细胞株中均有TLR5的表达,而且SPC-A-1中TLR5表达明显增高,这与我们在组织中观察到的结果大致一致。Sławińska A等[6-8]发现不同品种的鸡相同器官、组织上TLR5表达不一致,其表达的强度与由生物体的生理状态(尤其与炎症)密切相关,我们发现三种人肺癌细胞株TLR5的表达存在差异,因此推测可能与细胞株的细胞来源不同相关。免疫荧光结果还显示,TLR5在SPC-A-1细胞膜和细胞质均有表达,而且主要表达在细胞膜上,这与既往文献报道[9,10]相符。基于TLR5在NSCLC组织和不同NSCLC细胞株中的表达情况,因此我们选择SPC-A-1细胞株作为实验对象,研究TLR5与特异性配体结合后,其信号通路的活化情况。
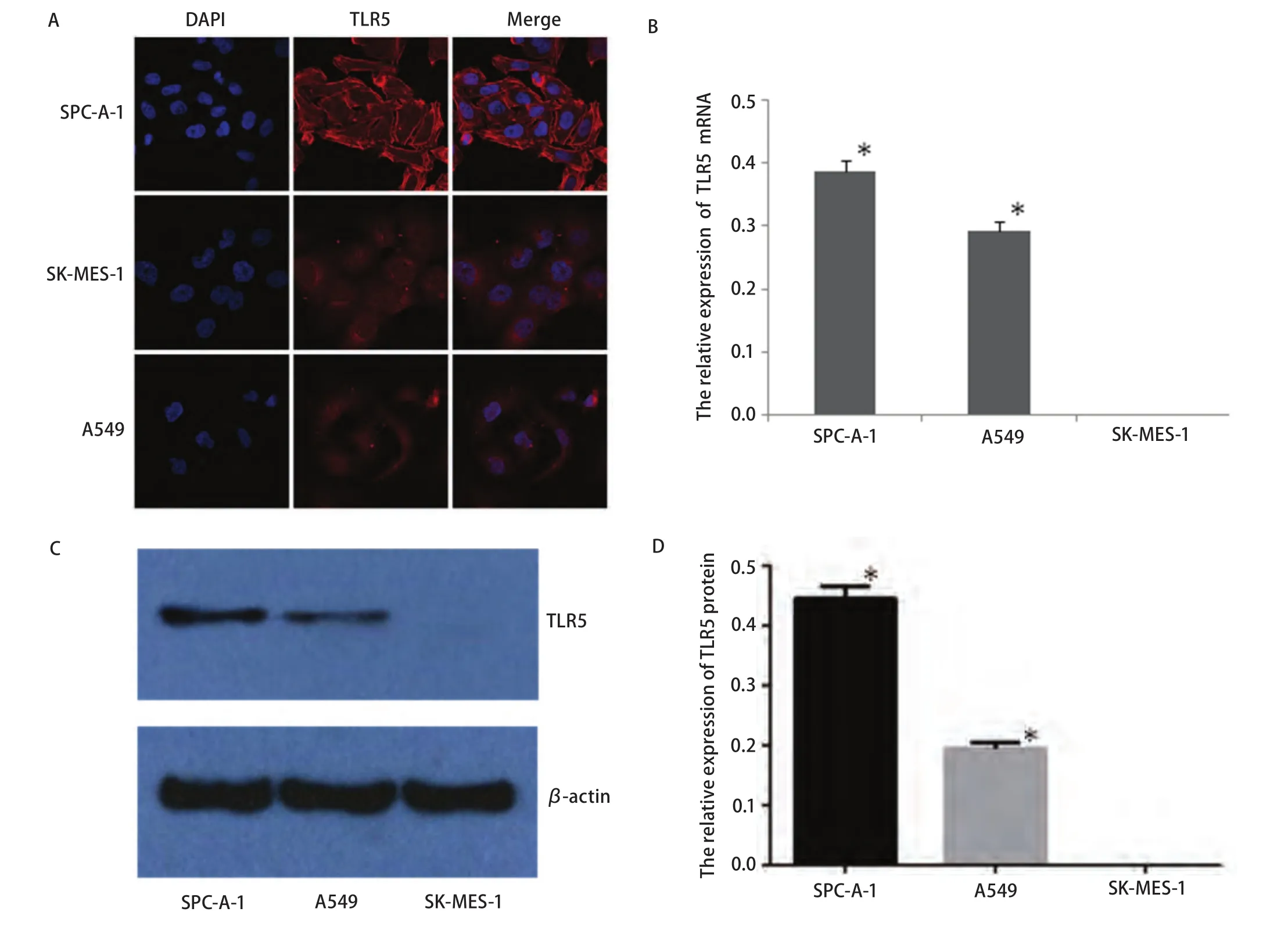
图1 TLR5在不同NSCLC细胞株中的表达。A:免疫荧光显示TLR5在三种NSCLC细胞株中的表达,其主要表达在细胞膜上;B:三种NSCLC细胞株中的表达TLR5 mRNA的相对表达量,与SK-MES-1细胞株相比,*P<0.05;C、D:Western blot检测三种NSCLC细胞株中TLR5蛋白的表达及其相对表达量,与SK-MES-1细胞株相比,*P<0.05。Fig1 Expression of TLR5 in three different NSCLC cell lines. A: TLR5 expression in three kinds of NSCLC cell lines by immunofluorescence, which is mainly expressed in the cell membrane; B: The relative expression of TLR5 mRNA in three kinds of NSCLC cell lines; compared with that of SKMES-1 cell lines, *P<0.05; C, D: the expression of TLR5 protein in three kinds of NSCLC cell lines and their relative expressions detected by Western blot; compared with that of SK-MES-1 cell lines, *P<0.05. NSCLC: non-small cell lung cancer.
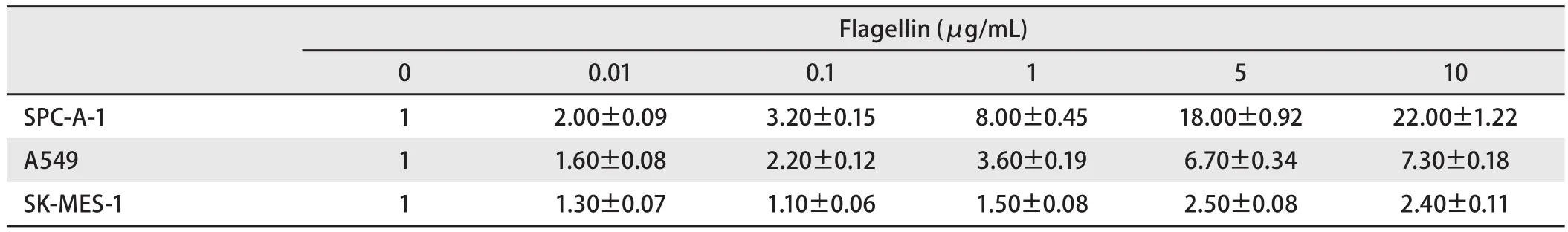
表2 Flagellin刺激后,NSCLC细胞内NF-κB荧光素酶相对活性Tab2 The relative activity of NF-κB luciferase of NSCLC cells after stimulated by flagellin
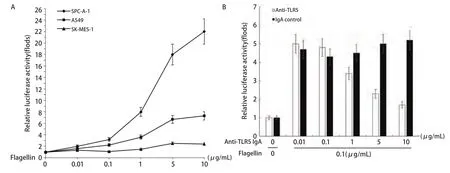
图2 Flagellin刺激后,细胞内NF-κB荧光素酶活性。A:三种NSCLC细胞株的NF-κB荧光素酶活性;B:加入TLT5抗体拮抗后,SPC-A-1细胞内NF-κB荧光素酶活性。Fig2 The relative activity of NF- kappa B luciferase of the cells when stimulated by flagellin. A: the relative activity of NF-κB luciferase of three kinds of NSCLC cell lines; B: After added with TLR5 antibody, the relative activity of NF-κB luciferase of SPC-A-1 cells.
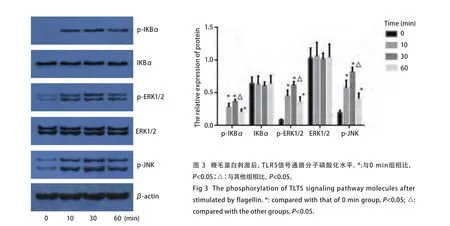
图3 鞭毛蛋白刺激后,TLR5信号通路分子磷酸化水平。*:与0 min组相比,P<0.05;△:与其他组相比,P<0.05。Fig3 The phosphorylation of TLT5 signaling pathway molecules after stimulated by flagellin. *: compared with that of 0 min group, P<0.05; △:compared with the other groups, P<0.05.
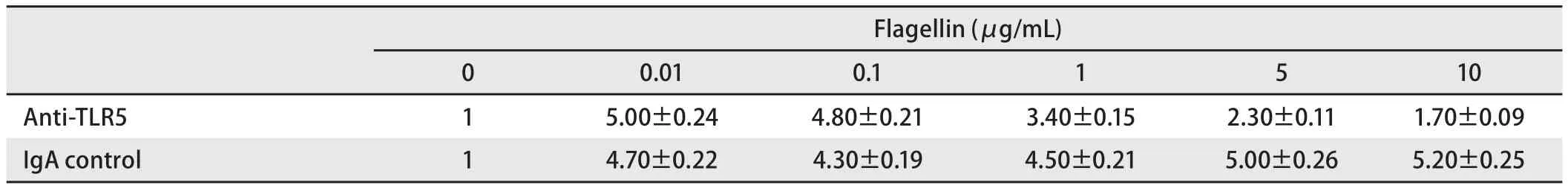
表3 加入TLR5抗体,Flagellin刺激SPC-A-1细胞,其细胞内NF-κB荧光素酶相对活性Tab3 The relative activity of NF-κB luciferase of SPC-A-1 cells stimulated by flagellin when added with TLR5 antibody
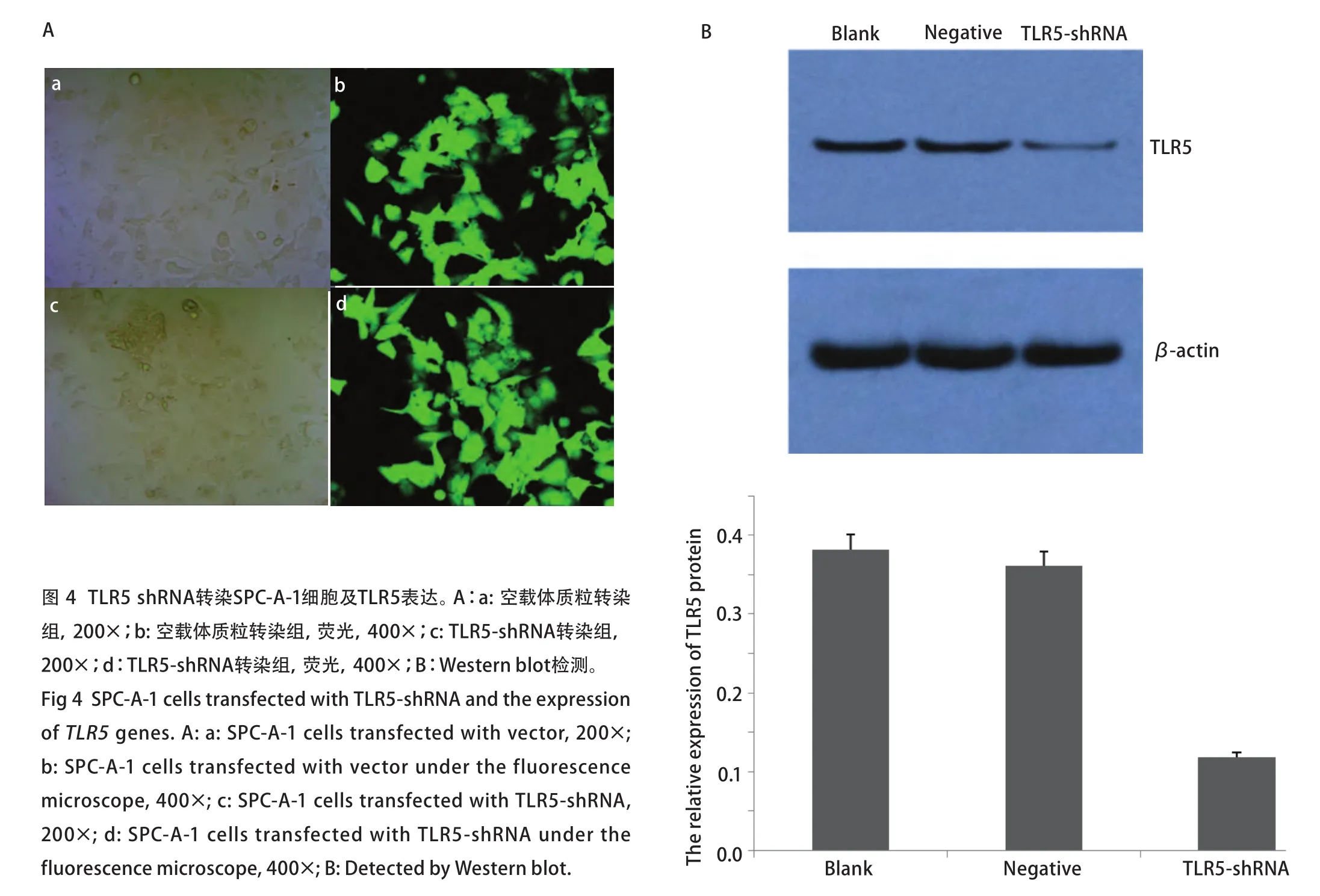
图4 TLR5 shRNA转染SPC-A-1细胞及TLR5表达。A:a: 空载体质粒转染组,200×;b: 空载体质粒转染组,荧光,400×;c: TLR5-shRNA转染组,200×;d:TLR5-shRNA转染组,荧光,400×;B:Western blot检测。Fig4 SPC-A-1 cells transfected with TLR5-shRNA and the expression of TLR5 genes. A: a: SPC-A-1 cells transfected with vector, 200×;b: SPC-A-1 cells transfected with vector under the fluorescence microscope, 400×; c: SPC-A-1 cells transfected with TLR5-shRNA,200×; d: SPC-A-1 cells transfected with TLR5-shRNA under the fluorescence microscope, 400×; B: Detected by Western blot.
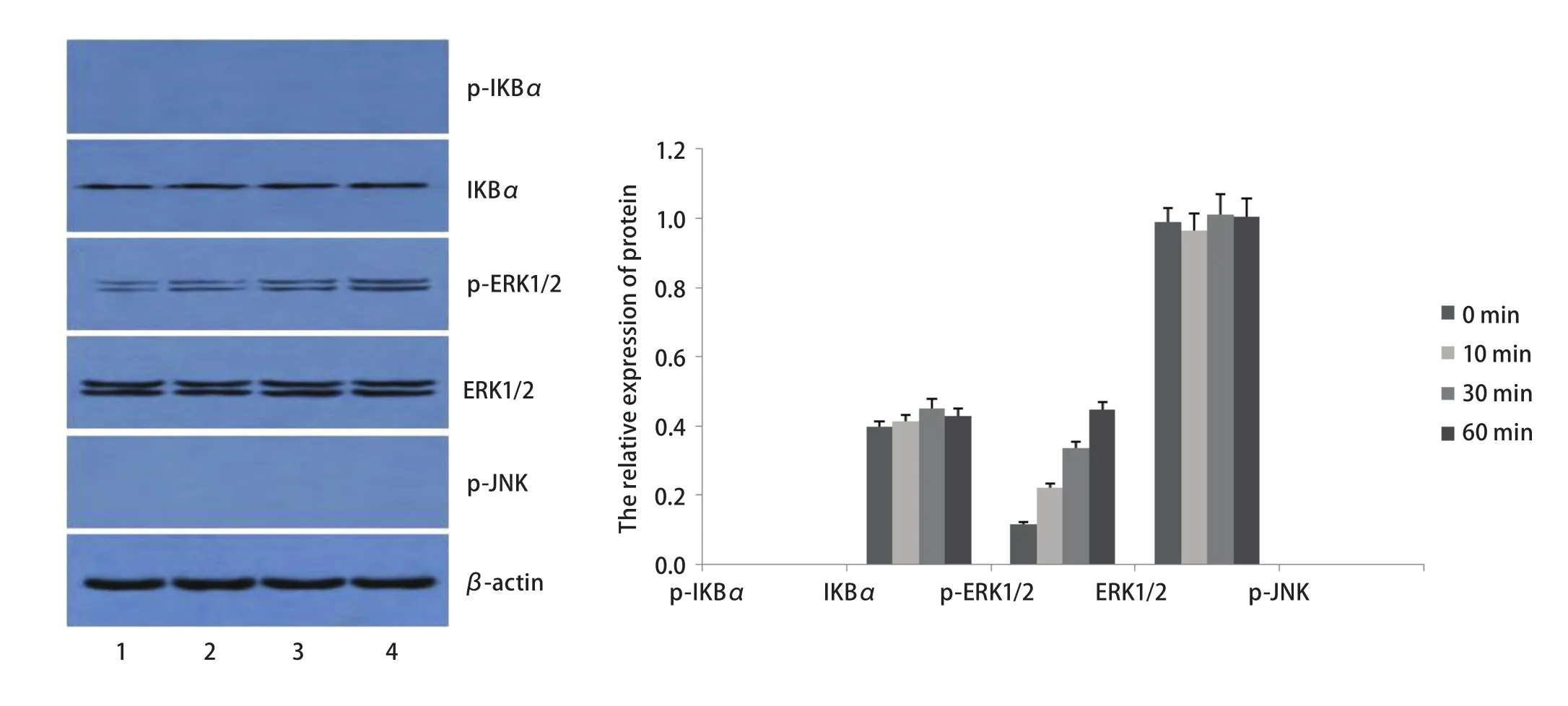
图5 Flagellin刺激TLR5 shRNA SPC-A-1细胞不同时间点TLR5通路分子的表达。1:0 min;2:10 min;3:30 min;4:60 min。Fig5 The relative expression of TLT5 signaling pathway molecules at different time after TLR5-shRNA SPC-A-1 cells were stimulated by flagellin. 1:0 min; 2: 10 min; 3: 30 min; 4: 60 min.
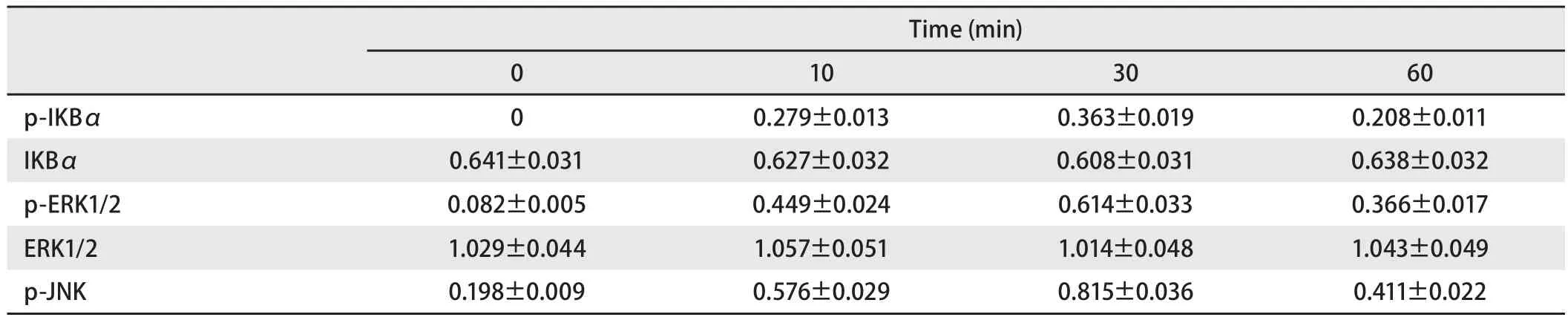
表4 鞭毛蛋白刺激后,TLR5信号通路分子相对表达量Tab4 The relative expression of TLT5 signaling pathway molecules after stimulated by flagellin
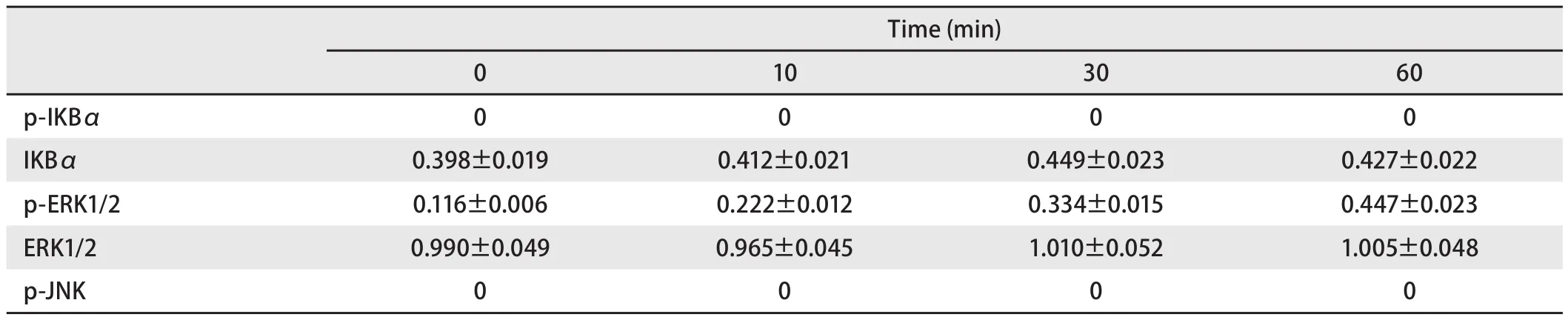
表5 Flagellin刺激TLR5-shRNA SPC-A-1细胞不同时间点TLR5信号通路分子相对表达量Tab5 The relative expression of TLT5 signaling pathway molecules at different time after TLR5-shRNA SPC-A-1 cells were stimulated by flagellin
TLR在癌细胞的表达增加,可上调NF-κB,产生抗凋亡蛋白,从而促进肿瘤细胞增殖;亦可介导肿瘤细胞释放趋化因子和细胞因子,诱导机体的免疫细胞参与肿瘤细胞微环境中的免疫反应。免疫细胞进一步释放炎性细胞因子、生长因子和血管形成因子,促进肿瘤细胞的生长、侵袭、新的血管淋巴管形成、转移。既往研究[11]还发现TLR4、TLR9、TLR7、TLR8等Toll样受体表达在人肺癌细胞株上,与肿瘤细胞的存活与化疗耐药性等相关。Cherfils-Vicini等[11]发现TLR7、TLR8可以表达在离体培养的肺癌组织和细胞株上,两者被激活后,NF-κB蛋白、抗凋亡蛋白Bcl-2的表达上调,增加肿瘤细胞的存活及其耐药性。进一步分析发现用配体刺激TLR7或TLR8后,在人肺原发肿瘤细胞和人肺肿瘤细胞株原位检测显示基因表达上调,提示肿瘤细胞受到慢性刺激,说明TLR信号可以直接有促进肿瘤的发生发展。我们选取SPC-A-1细胞株,发现随着鞭毛蛋白浓度的增加,TLR5被激活,其通路下游分子NF-κB荧光素酶的活性增强,且呈浓度依赖性;其活性能被TLR5抗体抑制,随着TLR5抗体浓度的增加,NF-κB荧光素酶的活性逐渐降低,与TLR5抗体浓度呈负相关。说明鞭毛蛋白活化了TLR5通路,并引起下游分子的活化,与既往文献相符。
树突状细胞(dendritic cells, DCs)位于抵御外来入侵第一道防线,接收到警报后可启动后天免疫系统。TLRs位于DCs上,是先天免疫模式识别的主要受体之一,它能识别许多病原微生物上的PAMPs,这些保守序列与TLRs结合后,进而活化机体先天免疫系统,导致一些亲炎症细胞因子的产生,因此TLRs被认为控制着由先天免疫向后天免疫的转变。我们前期研究发现TLR5蛋白不仅在NSCLC组织表达增加,还高表达于NSCLC细胞株中。由于肺癌组织或细胞中其内源性的配体目前并不明确,而鞭毛蛋白是TLR5已知特异的外源性配体,因此我们用鞭毛蛋白刺激TLR5,进一步研究其在NSCLC细胞中激活后,信号通路活化的情况。
TLR与特异PAMPs结合后,自身的异构形态发生改变,衔接分子被募集,下游分子陆续发生磷酸化和/或泛素化,大量蛋白质被募集,或结合,信号通路激活,引起炎性转录因子、炎性基因表达,机体发生免疫防御反应,产生炎症。在此通路中,NF-κB位于TLR下游信号通路的枢纽位置。NF-κB具有多向性调节作用的细胞核转录因子,也是能触发炎症、并广泛存在于炎症反应中的一种关键性因子。NF-κB参与诱导基因表达、调控免疫细胞的激活、肿瘤形成、细胞凋亡、细胞信号转导、炎症反应及多种自身免疫性疾病发生等过程[12,13],也是炎症和肿瘤的重要枢纽分子。我们检测到用鞭毛蛋白刺激SPC-A-1细胞后,p-IKBα、p-ERK1/2水平均在10 min开始升高,30 min达到高峰,60 min水平明显降低,而相应的的时间点IKBα、ERK1/2蛋白水平并无明显变化,说明SPC-A-1细胞受到鞭毛蛋白刺激后,引起胞浆内关键分子IKBα发生磷酸化,继而降解,引起NF-κB活化移位入胞核,启动下游ERK1/2蛋白磷酸化,及一系列的级联反应。而用鞭毛蛋白刺激TLR5 shRNA细胞后,在0 min、10 min、30 min、60 min四个不同的作用时间点,IKBα、ERK1/2蛋白的水平无明显变化,p-ERK1/2蛋白水平随着时间延长明显增高,而p-IKBα、p-JNK蛋白均未检出。说明TLR5基因并非完全被沉默,表达出的极少量TLR5蛋白可以引起ERK1/2的磷酸化。
NF-κB在肺癌组织中的的表达明显高于正常肺组织,提示在NSCLC的发生发展中可能起重要作用。而且烟雾中的化学物质可引起支气管损伤、支气管炎症,炎症中的一些细胞因子、趋化因子活化NF-κB,进而诱导与癌症相关的基因表达。我们的研究也发现吸烟的肺癌患者中TLR5表达高于非吸烟者,说明吸烟确实提高了支气管炎症的发生。
因此,从我们的实验结果可以推测出鞭毛蛋白刺激NSCLC细胞TLR5活化,通路下游具有活性的磷酸化p-IKBα、p-ERK1/2及p-JNK蛋白表达增多,在30 min达到高峰,NF-κB入核率增加,导致NSCLC细胞活化,但对其生物行为的影响还有待进一步研究。
猜你喜欢
材料与冶金学报(2022年2期)2022-08-10
天津医科大学学报(2021年4期)2021-08-21
昆明医科大学学报(2021年8期)2021-08-13
昆明医科大学学报(2021年3期)2021-07-22
科学与财富(2021年33期)2021-05-10
中日友好医院学报(2021年1期)2021-04-14
广东蚕业(2021年1期)2021-03-18
中西医结合肝病杂志(2020年2期)2020-10-27
作文成功之路·小学版(2020年6期)2020-07-27
作文成功之路·小学版(2020年5期)2020-06-11
