
都江堰地区枹栎果实特征的比较
2015-08-22 05:44唐东民王珑颉郗晓丹
四川环境 2015年4期
郭 强,唐东民,梁 赟,王珑颉,郗晓丹
(眉山市环境监测中心,四川 眉山 620010)
· 试验研究 ·
都江堰地区枹栎果实特征的比较
郭强,唐东民,梁赟,王珑颉,郗晓丹
(眉山市环境监测中心,四川 眉山620010)
种子大小及其变化不仅涉及到植物生长过程中的生殖模式,而且是植物种的重要适应特征。本文选择了都江堰花溪森林公园内的3种不同生境,以果实长度,宽度,长宽比及重量等作为指标,对不同生境中的常绿阔叶林主要伴生树种枹栎(Quercusserrata)的果实性状进行了研究和分析。结果表明:不同生境间的枹栎果实性状存在差异,但3种生境内枹栎果实各性状指标的变异幅度都不大,果实性状与植株胸径和高度之间没有显著的相关关系(P>0.01),枹栎果实重量与其长度间有明显的相关关系,但果实重量与其宽度及长宽比之间却没有显著相关性。
常绿阔叶林;枹栎;果实性状;生境;变异
种子是种子植物的繁殖体,种子对后代的资源投入将会直接影响后代的适合度进而影响到植被群落的更新,种子在大小上的变异是不争的事实[1,2],此外,种子大小也影响了幼苗的存活,生长和建立[3~8]。植物种子大小变异的研究一直是植物生殖生态学研究的重要方面。Harper等[9]曾将植物生活史的早期过程分为种子、萌发期、出苗期、成苗期等阶段,并认为植物种在群落中的更新策略主要包括种子雨、种子库和幼苗库等。因此,对植物种子性状的研究与分析具有重要的生态学意义。本文通过对都江堰花溪森林公园常绿阔叶林主要伴生树种枹栎果实性状的研究和分析,以期了解不同生境中枹栎果实的性状特征,为枹栎种群的生殖生态学研究提供一定的理论依据。
1 研究地区与研究方法
1.1自然概况
研究地点位于都江堰市的花溪森林公园(30°45′N~31°22′N,103°25′E~103°47′E),四川盆地西缘山地,属于中亚热带湿润气候区,四季分明,夏无酷暑(极端最高气温32℃),冬无严寒(极端最低气温-5℃),年均气温15.2℃,1月份平均气温4.6℃,7月份平均气温24.7℃,≥10℃的年积温4677.1℃,雨量充沛,年降雨量1300~1800 mm,云雾多,日照少(年日照时数只有800~1000 h),湿度大(年平均相对湿度80%以上),年均无霜期280d。由于气候温暖湿润,地带性植被发育形成生物多样性丰富的常绿阔叶林[10]。该森林公园内约有9 hm2的原生林被较好的保存下来,地带性植被是常绿阔叶林。乔灌层植被主要由壳斗科,樟科,山茶科的常绿阔叶树组成。主要建群种有栲树(Castanopsisfargesii)、润楠(Machiluspingii)、青冈(Cyclcobalanopsisglauca)等,主要的伴生树种有枹栎、栓皮栎(Quercusvariabilis)、薯豆(Elaeocarpusjaponicus)、江南石栎(Lithocarpusleistocarpus)等。林下灌木层主要有油茶(Camelliaoleifera)、黄牛奶(Symplocoslaurina)、海桐(Pittosporumtobira)、山矾(Symplocossumuntia)、柃木(Euryajaponica)、川冬青(Ilexszechwanensis)等。
1.2研究方法
在花溪森林公园选择不同生境作为枹栎果实的收集点,大致分为3种生境条件:(1)馒头山,此处为退耕还林后形成的灌丛群落(简称灌丛Shrub,以S表示,下同),枹栎植株较小,无高大乔木,光照充足,属于植被演替的初期;(2)般若寺附近的林场,原生林保存比较好,郁闭度较高,属于植被演替的后期阶段(简称成熟林Mature forest,以MF表示,下同);(3)花溪村农户附近的次生林,此处受到较强人为干扰,乔木层以壳斗科为主,群落演替阶段介于生境S和生境MF之间(简称次生林Second forest,以SF表示,下同)。3个收集点间距大约2 km,分别在3种生境内标记枹栎12株(同生境中植株间间隔大约10~30 m),在枹栎果实成熟与凋落期间(2006年9月底至11月初),从10月3日开始,每隔2 d或3 d在不同生境中收集果实,到10月15日止,共在每个生境内采种5次。每次每株枹栎收集果实15粒,同时记录所收集种子的母株的胸径(cm),高度(m)。
每次将收集到的枹栎果实带回实验室,从每个生境所采集的180粒果实中随机抽取90粒(30粒1组,3次重复)。在室内用游标卡尺测定果实最长处长度(cm),最宽处宽度(cm),并在80℃下烘36h后,用电子天平(精确度0.01g)称取每30粒果实的干重。
枹栎的果实属于坚果,长卵形,果实的性状指标一般包括其长度(cm),宽度(cm),重量(g)等,果实的长度与果实宽度的比值在一定程度上可反映果实的形态特征[11],作为一个衡量指标,可以把它称为果实的长宽比。果实性状指标以平均值±标准差表示。
1.3数据处理
每种生境选取450粒果实测定,共统计3个生境36株枹栎的1350粒果实。对所有果实性状数据作频度分析,单因素方差分析不同生境间的果实性状,并用邓肯氏多重比较检验方差分析中有显著差异的指标间的差异,并对果实性状与母株胸径,高度之间进行Pearson相关性分析,以上分析均用spss12.0软件包进行,作图用Excel完成。
2 结果与分析
2.1枹栎果实大小的变异
2.1.1枹栎果实长度分析
该地区枹栎果实长度分布呈正态分布(图1),其Skewness(即正态分布的偏度)值为-0.143,Kurtosis值为0.124,比标准正态峰略高。约有85.1%枹栎果实长度在1.62~2.22 cm之间。该地区中枹栎果实的平均长度为1.85±0.2cm。果实长度的变异系数较小,仅为4.0%。生境S,MF,SF的枹栎果实的长度也呈正态分布,生境S的枹栎果实的平均长度最小(表1),为1.82±0.20cm,其果实长度的变异幅度为3.8%。生境MF果实的平均长度最大,为1.87±0.18cm,但其变异系数也仅为3.3%。
2.1.2枹栎果实宽度
3种生境中枹栎果实宽度分布在0.67~1.87 cm之间,呈正态分布(图2),其Skewness值为0.281,Kurtosis值为0.404,比标准正态峰略高。大约有83.6%的果实分布在0.77~1.07 cm的宽度范围内。枹栎果实的平均宽度为0.95±0.108cm,变异幅度为1.2%。不同生境中枹栎果实宽度的变化情况见表1。生境S,MF,SF的枹栎果实的宽度也呈正态分布,平均宽度最大的是生境MF中的枹栎果实,为0.97±0.11cm,变异系数为1.2%,范围在0.69~1.77 cm之间。生境S和生境SF中的果实平均宽度分别为0.92±0.11cm和0.96±0.10cm,变异系数均较小,分别为1.2%和1.0%。

坐标数值范围(cm) 1(≤0.77),2(0.77~0.87),3(0.87~0.97),4(0.97~1.07),5(1.07~1.17),6(1.17~1.27),7(1.27~1.37),8(≥1.37)。图2 该地区枹栎果实宽度分布Fig.2 Distribution of Quercus serrata fruit width in this region
2.1.3枹栎果实长宽比
果实的长宽比值在一定程度上反映出果实的形状特征。花溪地区枹栎果实的平均长宽比为1.956±0.214,其Skewness值为-0.002,Kurtosis值为0.037,比标准正态峰略高(图3),约84.2%的果实长宽比值集中于1.70~2.30之间,变异系数为4.6%。生境S,MF,SF的枹栎果实的长宽比也呈正态分布,3种生境中的果实长宽比平均值都接近2(表1),分别为1.990±0.194,1.939±0.211,1.940±0.233。生境S的果实长宽比最大,果实显著偏长,生境SF中枹栎果实的长宽比变异幅度最大,为5.4%。

横坐标数值范围 1(≤1.40),2(1.40~1.55),3(1.55~1.70),4(1.70~1.85),5(1.85~2.00),6(2.00~2.15),7(2.15~2.30),8(2.30~2.45),9(2.45~2.60) ,10(≥2.60)。图3 该地区枹栎果实长宽比分布Fig.3 Distribution of ratio of Quercus serrata fruit length to width in this region
2.1.4枹栎果实重量
由于不同采摘期果实的含水量不同,本文以果实干重量作为衡量指标。3种生境中枹栎果实重量分布在0.24~0.49g之间,呈正态分布(图4),其Skewness值为0.854,Kurtosis值为0.419,比标准正态峰略高。大约有77.8%的果实分布在0.26~0.38g之间。枹栎果实的平均重量为0.34±0.06g,变异幅度为0.3%。不同生境中枹栎果实重量的变化情况见表1。生境S,MF,SF的枹栎果实的重量也呈正态分布,平均重量最大的是生境MF中的枹栎果实,为0.38±0.07g,变异系数为0.5%,范围在0.26~0.49g之间。生境S和生境MF中的果实平均质量分别为0.31±0.02g和0.33±0.04g,变异系数均较小,分别为0.0%和0.2%。
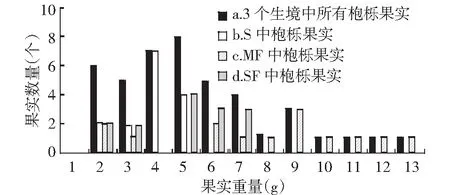
横坐标数值范围(g) 1(0.24~0.26),2(0.26~0.28),3(0.28~0.30),4(0.30~0.32),5(0.32~0.34),6(0.34~0.36),7(0.36~0.38),8(0.38~0.40),9(0.40~0.42),10(0.42~0.44),11(0.44~0.46),12(0.46~0.48),13(0.48~0.50)。图4 该地区枹栎果实重量分布 Fig.4 Distribution of Quercus serrata fruit weight in this region
2.2不同生境中枹栎产生的果实性状差异
邓肯氏多重比较显示(表1),生境S中枹栎果实的平均长度和平均宽度均为最小,分别为1.82±0.20cm和0.92±0.11cm,而且与生境MF和SF之间存在着显著差异。果实长宽比平均值最大的是生境S,为1.990±0.194,也与其它2个生境间表现出显著差异。生境MF和SF在果实长度,宽度以及长宽比等性状上未表现显著差异,但在果实重量上有显著差异。在3种生境中,果实重量的变异系数最小,分别为0.0%,0.5%,0.2%,而变异幅度最大的是果实长宽比,达到了3.8%,4.5%,5.4%。

表1 不同生境间枹栎果实长度、宽度、长宽比以及重量的比较
S为灌丛,SF为次生林,MF为成熟林;不同字母为差异显著(P<0.01)。
方差分析的结果(表2)表明,在不同生境条件下枹栎果实的性状指标中,果实长度,果实长宽比及果实重量有一定的差异(P<0.01),其中果实宽度在3种生境间的差异很显著,F值为30.453,自由度为2。而果实长度,长宽比和重量的F值仅分别为7.402,8.437和7.881(自由度均为2)。

表2 不同生境间枹栎果实性状的方差分析
2.3枹栎果实性状与植株个体大小间的关系
通过相关性分析,对枹栎果实性状而言,无论是果实平均长度,平均宽度,平均重量,还是平均长宽比,与植株的胸径和高度之间没有显著的相关关系(表3)。果实长度和果实长宽比平均指标与植株胸径,高度之间的Pearson相关系数小于0.1,果实宽度,果实重量与植株胸径,高度间的Pearson相关系数稍大,但也不明显,分别为0.193和0.231,0.184和0.360(P>0.05)。
2.4枹栎果实性状间的相关关系
枹栎果实的平均重量与它的平均长度之间有明显的相关性(P<0.05),其中生境S果实重量与长度之间的Pearson相关系数为0.574(P<0.05)(图5a),其方程表示为W重量=-0.28+0.32L长度,生境MF果实重量与长度之间的相关系数为0.721(P<0.01)(图5b),其方程表示为W重量=-1.16

表3 枹栎果实性状与植株胸径,高度间的Pearson相关分析
+0.85L长度,生境SF果实重量与长度之间的相关系数为0.606(P<0.05)(图5c),其方程表示为W重量=-0.57+0.48L长度,而综合3个生境中所有枹栎果实的重量与长度间的相关系数为0.310(P<0.05)(图5d),方程为W重量=-0.27+0.33L长度。通过分析表明,3个生境中的枹栎果实重量与其宽度和长宽比之间没有显著的相关性(P>0.05)。

a:生境S果实质量与其长度间的相关关系; b:生境MF果实质量与其长度间的相关关系c:生境SF果实质量与其长度间的相关关系;d:3个生境中果实质量与其长度间的相关关系图5 枹栎果实重量与其长度间的相关关系Fig.5 Correlation between fruit weight of Quercus serrata and its fruit length
3 讨 论
在较大地理尺度范围内,种子大小变化与气候等因素有密切关系[12,13]。蔡永立等[11]对东部亚热带青冈果实的研究发现,青冈果实大小和性状的变异与纬度和经度均表现较高的负相关,南部地方青冈果实常大而偏圆,而较北部地区的果实相对小而偏长,并认为这可能是对不同环境所采取的繁殖策略。植物种子的大小不仅受遗传机制控制,同时也受不同环境因子的影响。植物种子的大小变化被认为是自然选择下的一种“妥协”[14]。
本文研究了较小尺度范围内生境各异的枹栎种群,结果表明不同生境条件中的枹栎个体产生的果实性状间有差异,生境S即馒头山退耕地灌丛群落中的枹栎果实平均长度,宽度及重量最小(1.82±0.20cm,0.92±0.11cm,0.31±0.02g),并与其他2种生境中的果实形态特征有显著的差异(P<0.01)(生境S与生境SF的果实重量没有显著差异可能与采集方式不同有关,生境S植株较小,果实主要从植株上摘取,生境SF中则主要是从地上拾取较新鲜的果实),而该生境条件中枹栎的果实平均长宽比却最大,也与其它生境存在明显的差异,相对于其他2种生境中的枹栎果实表现出小而偏长。
果实的大小与母株所处立地生境中土壤的养分,水分以及光照,温湿度等因素有关[15]。在花溪森林公园常绿阔叶林中,生境MF植被是以壳斗科,樟科,山茶科等组成的群落地段,以栲树,润楠,青冈栎为建群种,该地区属植被演替的后期阶段,栲树,润楠,马尾松等个体占据着上层乔木层,枹栎,栓皮栎,薯豆,江南石栎等伴生种位居乔Ⅱ层,枹栎虽然不是优势树种,但数量较多,分布密集,此外,土壤营养和水分状况较好,树冠郁闭度较高。
生境SF是受人为干扰较强的次生林,在该生境中,土壤条件与生境MF相似,但建群种不明显,枹栎数量相对较多,林冠稀疏,郁闭度较低。而生境S是退耕还林地,立地生境无高大乔木,以灌丛为优势群落,光照强烈,但土壤肥力较差,保水性弱,属于植被演替的初期。该生境中,枹栎植株较小,数量较少且分布间隔大。
分析结果表明,生境MF和SF之间,枹栎果实的各性状指标(除重量外)均没有明显差异。推断原因可能是:1.两种生境中土壤条件相似,养料和水分充足;2.与枹栎的繁殖方式有关,其传粉方式为风媒和虫媒兼有,在生境MF和生境SF群落内,枹栎个体分布相对密集,传粉昆虫种类也较多,在花期,个体间极易互相传粉,有利于提高受粉率,传粉的水平与效率也会影响到果实大小,大量花粉的存在有助于提高生殖个体之间的传粉效率,并有可能保证胚珠的良好发育和生长,在花粉数量较少的条件下,植物易产生干瘪或发育不好的果实[16~19]。而处于生境S中的枹栎个体虽能接受较好的光照,但土壤营养状况和水分相对较差,在不太有利的自然条件下可能会减少植株对有性生殖的投资,同时植株个体较小,数量少,且相距较远,不利于传粉,使其受精的效率和果实的发育受到影响。
Winn[20]的研究表明,在具有较高草本覆盖的生境中,其中的植物均生产较小的种子,而在低草本覆盖的生境移栽的和当地种会生产较大的种子。本文所研究的3种生境中,S的草本覆盖率很高,而MF和SF内则较少草本分布,得到的结果显示,就枹栎而言,与Winn的这一规律相符。
一般认为平均果实的大小与亲本能力有关,一些植物的株高,胸径与果实大小间有很强的相关关系[21],但本研究结果表明,无论是果实长度,宽度,重量还是长宽比与植株的胸径和植株高度之间的Pearson相关系数均较小,这同早期对Quercus和Pinus等属中植物果实的研究结果相同[22]。从枹栎果实的大小与植株间没有明显关系这点推测,果实大小差异可能与植株所处的不同生境以及在不同生境中采取的生殖方式有关。
Salisbury[23]的研究表明, 群落演替高级阶段中的物种比低级阶段中的物种种子较重。王伟伟等[24]的研究也表明,植物在郁闭的生境中倾向于产生较大的种子。枹栎个体在生境MF和SF中产生的果实,显著大于生境S中的果实,这种变异也可能与它们所处的演替阶段以及立地生境完全不同的光照条件有关。大的种子固然有利于萌发和幼苗生长,小种子却容易逃避动物的捕食,传播范围广,利于种群的扩散,并且增加与土壤缝隙的接触,对于处于群落演替初期的物种而言,较小的种子更有利于种群的扩张。因此推测枹栎果实的性状变异是为了保证其在异质生境中的萌发,提高幼苗的竞争力与生存能力,从而确保有较大数量幼苗库,维持种群更新和稳定发展。
[1]武高林,杜国桢,尚占环. 种子大小及其命运对植被更新贡献研究进展[J]. 应用生态学报,2006,17(10): 1969-1972.
[2]Rees M. Trade-offs among dispersal strategies in British plants[J]. Nature, 1993,366: 150-152.
[3]Howe HF,Richter WM. Effects of seed size on seedling size in Virola surinamensis; a within and between tree analysis[J]. Oecologia, 1982,53: 347-351.
[4]Winn A A. Ecological and evolutionary consequences of seed size in Prunella vulgaris[J]. Ecology, 1988,69: 1537-1544.
[5]Houssard C,Escarré J. The effects of seed weight on growth and competitive ability of Rumex acetosella from two successional old-fields[J]. Oecologia, 1991,86: 236-242.
[6]Mendez M. Sources of variation in seed mass in Arum italicum[J]. International Journal of Plant Sciences, 1997,158: 298-305.
[7]Eriksson O. Seed size variation and its effect on germination and seedling performance in the clonal herb Convallaria majalis[J]. Acta Oecologica, 1999, 20: 61-66.
[8]Terry J L,Robert H J. Seedling growth strategies and seed size effects in fourteen oak species native to different soil moisture habitats[J]. Trees, 1996, 11: 1-8.
[9]Harper J L,Lovell P H,Moore K G. The shapes and sizes of seeds[J]. Annual Review of Ecology and Systematics, 1970, (1): 327-356.
[10]陈昌笃. 都江堰地区――横断山北段生物多样性交汇、分化和存留的枢纽地段[J]. 生态学报, 2000,20(1):28-34.
[11]蔡永立, 王希华, 宋永昌.中国东部亚热带青冈果实形态变异的研究[J]. 生态学报, 1999,19(4): 580-586.
[12]Foster S A. On the adaptive value of large seeds for tropical moist forest trees: a review and synthesis[J]. Botanical Review, 1986, 52:261-299.
[13]Mazer S J. Ecological, taxonomic and life history correlates of seed mass among Indiana dune angiosperms[J]. Ecological Monographs, 1989, 59:153-175.
[14]Silvertown J W,Doust J L. Introduction to Plant Population Biology[A]. London:Blackwell Scientific Publication,1993,105-121.
[15]Wulff R D. Seed size variation in Desmodium paniculatum. I. Factors on affecting seed size[J]. The Journal of Ecology, 1986, 74: 87-97.
[16]Houle G. Mast seeding in Abies balsamea, Acer saccharum and Betula alleghaniensis in an old growth, cold temperate forest of north-estern North America[J]. Journal of Ecology, 1999,87: 413-422.
[17]Quesada M, Winsor J A, Stephenson A G. Effects of pollen competition on progeny performance in a heterozygous cucurbit[J]. The American Naturalist, 1993,142: 694-706.
[18]Sork V L. Evolutionary ecology of mast-seeding in temperate and tropical oaks(Quercus spp.) [J].Plant Ecology, 1993,107: 133-147.
[19]何承刚,毕玉芬,姜华,等.紫花苜蓿的花蜜量和访花蜜蜂数量对种子产量的影响[J]. 生态学杂志,2005,24(12):1388-1391.
[20]Winn A A. Effects of seed size and microsite on seedling emergence of Prunella vulgaris in four habitats[J]. The Journal of Ecology, 1985,73(3): 831-840.
[21]Stocklin J,Favre P. Effects of plant size and morphological constraints on variation in reproductive components in two related species of Epilobium[J]. The Journal of Ecology, 1994, 82:735-746.
[22]Thompson K,Rabinowitz D. Do big plants have big seeds? [J]. The American Naturalist, 1989,133(5): 722-728.
[23]Salisbury E J. The reproductive capacity of plants[J]. Ecology, 1942, 24:269.
[24]王伟伟,苏智先,胡进耀,等.珍稀濒危植物珙桐不同采收期的种子特性研究[J]. 广西植物,2006,26(2): 178-182.
Comparison of the Fruit Characteristics of Quercus Serrata in Dujiangyan Region
GUO Qiang, TANG Dong-min,LIANG Yun,WANG Long-jie,XI Xiao-dan
(MeishanEnvironmentalMonitoringCenter,Meishan,Sichuan620010,China)
Size and variation of the seed were not only related to reproductive pattern of plant but also the important characteristic of plant adaptation. This article had chosen 3 different habitats in Hua-xi Forest Park of Dujiangyan as the study sites and analyzed the fruit characteristics of evergreen broad-leaved species- Quercus serrata with fruit length, width, aspect ratio and weight as indicators. The results showed a significant difference among the fruits from different habitats, but with small variation range of the fruit characteristics indexes among 3 habitats, and there were no obvious correlation between fruit characteristic and the height and diameter of the plant(P>0.01). The weight of fruit presented a significant correlation with its length but a non-significant correlation with its width and length /width ratio.
Evergreen broad-leaved forest; quercus serrata; fruit characteristics; habitats; variance
2014-12-26
郭 强(1981-),男,四川大邑人,2008年毕业于西华师范大学环境科学专业,硕士,工程师,主要从事生态环境、生态恢复研究。
X173
A
1001-3644(2015)04-0017-06
猜你喜欢
落叶果树(2021年6期)2021-02-12
趣味(数学)(2020年4期)2020-07-27
支部建设(2020年15期)2020-07-08
文苑(2020年6期)2020-06-22
体育科技(2016年2期)2016-02-28
百科知识(2015年18期)2015-09-10
西华师范大学学报(自然科学版)(2015年3期)2015-02-27
应用海洋学学报(2014年2期)2014-11-26
筑路机械与施工机械化(2014年2期)2014-03-01
诗潮(2014年7期)2014-02-28
