
有氧耐力可训练性差异与运动适应性循环microRNA表达谱特征的关联性
2015-08-04 06:51薛源李岩徐思
体育学刊 2015年4期
薛源++李岩++徐思
摘 要:miRNA调控机体对训练应激的适应能力,其调控模式影响有氧耐力可训练性。循环miRNA(c-miRNA)与miRNA密切相关,分析不同有氧耐力可训练性水平群体的运动适应性c-miRNA表达变化差异,可获得与训练敏感性相关的c-miRNA表达谱特征。结果显示:高可训练性表型在有氧训练应激诱导下差异表达17条c-miRNA,其中11条上调,6条下调或平稳,其调控功能主要涉及低氧适应通路和脂肪酸β氧化代谢的关键基因表达。提示:有氧耐力可训练性差异与机体低氧适应能力和脂肪酸β氧化供能能力,对有氧训练应激诱导作用的反应性密切相关。miRNA调控并整合机体应激适应性基因表达。c-miRNA差异表达谱可用来评估有氧耐力可训练性,预测有氧能力发展潜力。
关 键 词:运动生物化学;有氧耐力;可训练性;循环微核苷酸;最大摄氧量;关联模型;
运动性适应
中图分类号:G804.7 文献标志码:A 文章编号:1006-7116(2015)04-0139-06
Correlation between aerobic endurance trainability difference and the expression
spectrum characteristics of sport adaptive circulating microRNA
XUE Yuan1,LI Yan2,XU Si3
(1.School of Physical Education,Sichuan Normal University,Chengdu 610101,China;
2.School of Physical Education,Ludong University,Yantai 264025,China;
3.Sichuan Academy of Medical Science,Sichuan Provincial Hospital,Chengdu 610031,China)
Abstract: miRNA regulates the bodys ability to adapt to training stress; its regulating pattern affects aerobic endurance trainability. Circulating miRNA (c-miRNA) is closely related to miRNA; by analyzing differences in the changing of sport adaptive c-miRNA expression of groups of people at different aerobic endurance trainability levels, the authors can acquire the expression spectrum characteristics of c-miRNA sensitively correlative with training. Results: induced by aerobic training stress, the phenotype with high trainability differentially expressed 17 c-miRNAs, 11 of which were up-regulated, 16 of which were down-regulated or steady, its regulating function mainly involved with hypoxia adaptation pathway and the key gene expression of fatty acid β-oxidation metabolism. Hint: aerobic endurance trainability difference is close related to the reactivity of the bodys hypoxia adaptation ability and fatty acid β-oxidation energy supply ability to the induction function of aerobic training. miRNA regulates and integrates the bodys stress adaptive gene expression. C-miRNA differential expression spectrum can be used to evaluate aerobic endurance trainability, and to predict aerobic ability development potential.
Key words: sports biochemistry;aerobic endurance;trainability;c-miRNAs;maximum oxygen uptake;correlation model;sports adaption
杰出运动能力的形成与运动天赋和长期刻苦训练密不可分。运动性适应能力好的运动员的可训练性更强,成绩提高更快,训练效果更好[1]。表观遗传学机制调控营养、训练等环境应激因素诱导的适应过程的基因选择性表达,其基因调控模式具有个体差别并可稳定遗传[2],可能是产生运动能力及可训练性个体差异的原因。目前对于表观遗传学基因调控模式与运动能力的可训练性的具体关联所知甚少,因而无法将其作为指标应用于运动员选材和个性化训练。
microRNA基因表达调控作用是环境应激适应性基因表达的主要表观遗传学调控方式[3],其调控模式与环境应激适应能力的个体差异有关,应激适应过程中的microRNA表达谱反映了其调控模式和强度[4]。研究表明,microRNA分子(以下简称miRNA)参与了运动性适应基本生理过程的调控,如肌组织肥大和心肌/骨骼肌收缩力增强[5]、血管增生[6]、线粒体合成与酶活性提高[7]等。组织内miRNA分泌进入血液循环成为循环miRNAs(circulating miRNAs,以下简称c-miRNA),二者在表达谱上具有稳定关联[8]。c-miRNA可能是运动应激诱导机体应答过程中整合多系统适应性反应的关键因子[9],其表达谱特征与运动训练方式和训练效果存在关联,在运动性适应的不同状态下,c-miRNAs表达谱发生特征性改变[10],因此推测:运动适应性c-miRNAs表达谱可以作为反映运动能力的可训练性的“分子指纹”,在选材阶段准确预估运动能力的发展潜力,或用来评估训练效果/竞技状态,作为制订个性化训练方案的依据。
有氧耐力水平是决定运动能力的主要因素之一,不同个体的有氧耐力训练效果存在显著差异[11]。本研究以最大摄氧量(VO2max)为主要指标评估有氧耐力训练效果和训练敏感性,同步检测运动适应性c-miRNA表达谱,提取和分析有氧耐力训练的高敏感表型个体的运动性适应c-miRNA表达谱特征,建立运动性适应c-miRNA表达谱与有氧运动能力训练敏感性的关联模型,作为早期预测和评估有氧运动能力发展潜力和适应特点的指标应用于运动员选材和个性化训练方案制订。
1 研究对象与方法
1.1 研究对象
四川省某文武学校初中部学员92人,男性,平均年龄(13.6±0.5)岁;身高(171.85±6.28)cm;体重(59.60±4.55) kg。既往健康,无伤病史,参加试验前未受过系统的有氧运动能力训练。
1.2 有氧耐力训练方案
参加实验学员进行为期15周的系统性有氧运动能力训练。每周三、五下午16:00-17:30进行5 000 m长跑训练,负荷强度:1~3周65%个人最大心率±3次/min;4~10周70%个人最大心率±3次/min;11~15周75%个人最大心率±3次/min。以polar表(瑞典产)监控靶心率维持速度。训练方案由教练监督执行。
1.3 有氧运动能力相关指标检测
所有指标在15周有氧耐力训练前后各检测1次,分别记为基础值(baseline)和训练值(post-ex)。
1)最大摄氧量相对值(R-VO2max)测试采用12 min跑法[12],在400 m标准跑道上进行,受试者佩戴polar表,按指令起跑,尽最大努力跑完12 min,读取跑动距离s并换算成英里,按以下公式折算成R-VO2max:
R-VO2max(mL/(min·kg))=(s-0.318)/0.0278 ①
2)个体乳酸阈(ILT)测定采用递增负荷法[13],功率自行车(瑞典产Monark894E、839E)起始负荷为50 W,每2 min递增40 W,转速维持在50 r/min,直至力竭(力竭标准:连续2次不能维持规定转速)。取中指指尖血20 μL,采用YSI1500便携式血乳酸自动分析仪测定血乳酸值。分别采取安静状态下、每级负荷后即刻及恢复期第2、5、8、10、15 min血样。测试结果记录在表中,采用Stegmann方法判定ILT[14]。
1.4 c-miRNA表达谱检测和实时荧光定量PCR法验证
1)c-miRNAs表达值测定。
15周有氧耐力训练开始前清晨空腹安静状态下肘静脉取血5 mL检测c-miRNAs表达值,记为基线表达值。完成VO2max基础值测试后分别于1 h内、恢复24和48 h肘静脉取血5 mL检测c-miRNAs表达值,记为测试后0、恢复24 h、恢复48 h表达值。
2)血浆总RNA提取。
TRIZOL法提取血浆总RNA,RNase-free ddH2O溶解提取的总RNA分子。紫外分光光度仪测定RNA纯度和浓度,OD 260/280>1.9说明总RNA纯度较好,②甲醛变性琼脂糖电泳检测总RNA完整性较好,未发生降解。
3)microarray检测c-miRNA差异表达谱。
总RNA按照Ambions miRNA Isolation Kit说明书分离miRNA。使用miRCURYTM Array Labelling kit(Ambion)进行miRNA标记操作。microRNA芯片杂交按照miRCURYTM Array microarray kit (Exiqon)说明书进行。Genepix 4000B双通道激光图像扫描仪(Molecular Devices公司)以635 nm单波长进行扫描,Genepix Pro 6.0软件分析数据。数据预处理以后,根据各张芯片的global mean 进行片间校正,用SAM (significance analysis of microarrays,SAM .version2.1)筛选差异表达miRNAs。筛选条件:FDR (false discovery rate)<5%,Fold change>2或<0.5。
4)实时荧光定量PCR(qRT-PCR)验证差异表达c-miRNA相对表达量。
(1)c-miRNA茎环反转录合成cDNA:根据mirBASE提供的miRNA种子区序列,设计茎环反转录引物(北京六合华大基因公司合成),20 μL反应体系(5×First Strand synthesis Buffer,4 μL;dNTPmi 1uL;RNase Inhibitor,40units;Reverse Transcriptase(M-MLV),1μL;Stem-loop RT Primer,1μL;Total RNA 500 ng;RNase-free ddH2O),反应条件:65 ℃加热5 min,放入冰中2 min。然后放入PCR仪16 ℃热激30 min,42℃ 60 min,85 ℃,5 min停止反转录。
(2)qRT-PCR检测c-miRNA表达水平:miRNA定量引物(北京六合华大基因公司)。反应体系按照SYBR? Premix Ex TaqTMⅡ(Perfect Real Time)说明书进行配置。使用Bio-Rad iCycler PCR System进行Real Time PCR反应,程序:95 ℃ 5 min;95 ℃ 10 s;60℃ 30 s,40cycle。以上循环结束后进行65~95 ℃的融解曲线分析。每个样品平行3次,溶解曲线为单一峰。按照2-ΔΔCt计算样品的相对表达丰度。
1.5 c-miRNA差异表达谱的获得及其与有氧耐力指标的相关性
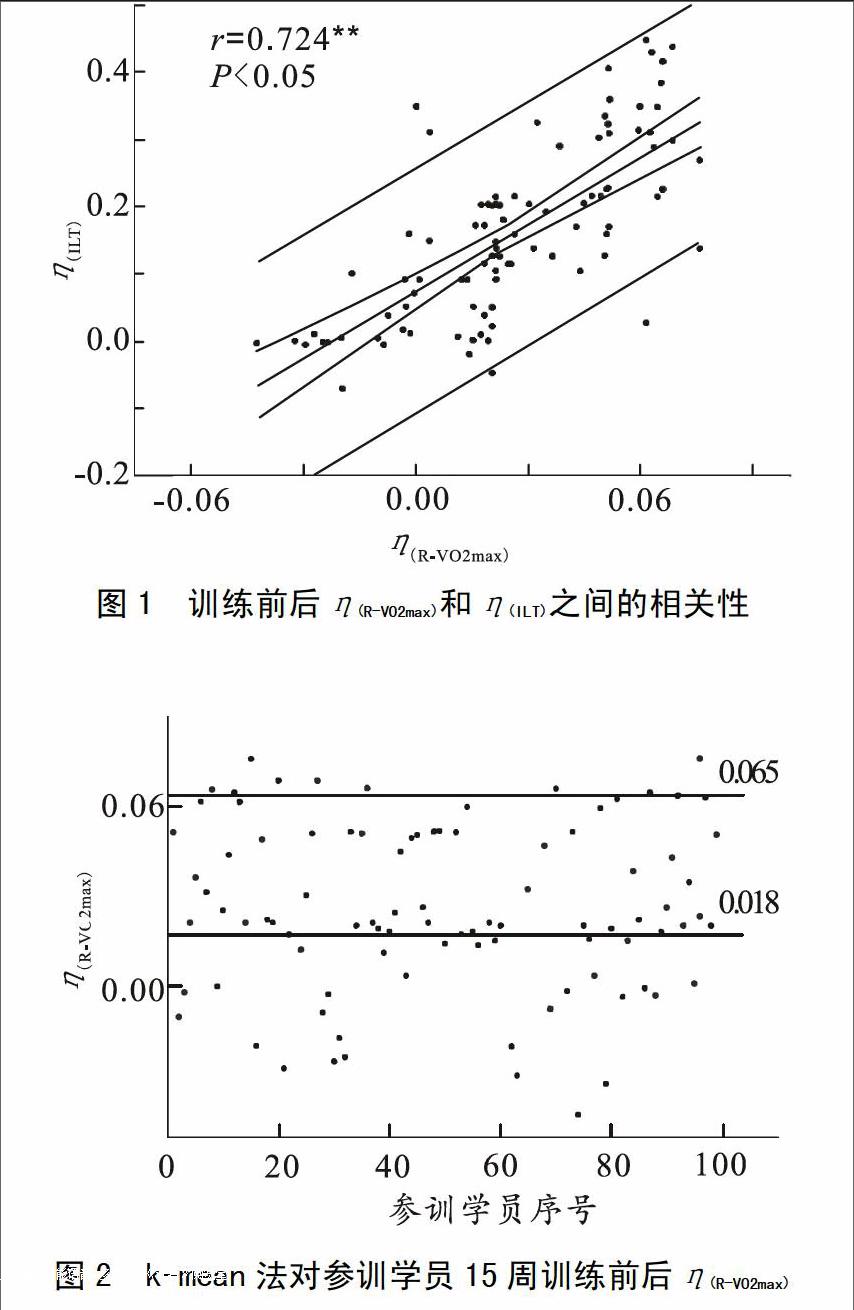
1)计算训练前后R-VO2max变化率η(R-VO2max)并采用K-means法进行聚类分析,根据聚类分析结果将学员分为对有氧训练的高敏感表型组(High Response genotype Group,简称HR组)和普通敏感表型组(Common Response genotype Group,简称CR组)。比较两组c-miRNA表达水平,参照Davidsen标准[10]提取HR组c-miRNA差异表达谱。
2)计算HR组差异表达的c-miRNAs在基线值-0 h阶段(以下简称为阶段Ⅰ)和0~24 h阶段(以下简称为阶段Ⅱ)的表达量变化率η(c-miRNA):
η(c-miRNA)=[(阶段终点表达量-阶段起点表达量)/阶段起点表达量]×100%
3)计算η(c-miRNA)与η(R-VO2max)和η(ILT)之间的Pearson相关系数,分析其相关性。
1.6 数据处理与分析
采用SPSS 17.0软件对结果进行处理。各项指标数据均采用(±s)表示。有氧耐力训练前后各项指标的变化率η(Χ)=[(训练值-基础值)/基础值]×100%。均数间比较采用两独立样本的t检验,P<0.05表示两组间的差异有统计学意义。聚类分析采用k-means法。采用Bivariate过程进行两组数据之间的相关性分析,计算Pearson相关系数并用r表示。
2 研究结果及分析
2.1 训练前后R-VO2max与ILT变化率及其相关性
3 讨论
本研究探讨了运动适应性c-miRNA表达谱与有氧耐力的可训练性之间的关联,结果发现:η(R-VO2max)显著高于普通水平的表型个体有17个c-miRNA差异表达,其功能主要富集于低氧适应通路和脂肪酸β氧化代谢途径,调控毛细血管增生、血红素生成、糖酵解相关酶功能、线粒体脂肪酸β氧化功能及骨骼肌重塑等适应性生理反应的关键基因表达,影响机体对训练应激的适应性。其作用特点为协同性调控多系统对耐力训练的应激适应。本研究结果提示:运动能力及其可训练性与miRNA调控基因选择性表达的模式有关。运动应激诱导的特征性c-miRNA表达谱与杰出运动能力表型存在关联,可以用来预测运动能力的发展潜力,将具有相关运动天赋的运动员早期选拔出来加以系统培养,或用以评估运动员的训练适应状态/竞技状态,制订个性化训练方案。
遗传选材的一个主要问题是如何评估个体的运动能力可训练性,预测其运动能力发展潜力[15]。杰出运动能力是遗传优势和营养、训练等环境因素共同作用下的积累效应。机体对训练应激的适应能力有条件地影响遗传优势的表型化程度,决定了运动能力的可训练性。应激诱导miRNA表达,调控基因表达强度,产生适应性的个体差异[17]。比较分析不同可训练性群体的适应性miRNA表达谱差异,不仅可以建立高可训练性群体的c-miRNA表达特征关联模型,还提供了训练应激与机体相互作用影响运动能力的有价值的信息。
最大摄氧量是评价有氧耐力的主要指标。个体VO2max主要受遗传影响,耐力训练可以在一定程度上改变VO2max水平,相同训练条件下VO2max变化率反映了心肺有氧机能可训练性的个体差异[18]。本研究采用VO2max变化率作为区别有氧能力可训练性的指标分析运动应激诱导的c-miRNA表达差异,采用Targetscan、miRanda和mirBASE联合分析差异表达的miRNA的靶基因,并进行GO分析和KEGG pathway分析,结合靶基因主要生理机能和代谢通路的共性特征研究HR组c-miRNA表达谱的调控特征。结果显示:差异表达的17条miRNA的靶基因主要富集于低氧适应通路和PPAR信号通路,HR组miRNA表达模式促进相关代谢通路的应激适应能力,并显示出多靶整合效应特征。
低氧应激激活HIF信号通路适应是获得有氧耐力训练效果的重要途径[19]。常氧分压条件下,有氧耐力训练引起的低氧应激主要发生在心肌/骨骼肌等组织内[20]。有氧工作引起耗氧量增加,组织内氧分压显著降低,抑制HIF-1a降解,上调HIF-1a活性,调控下游EPO、VEGF等低氧适应基因表达水平上调,产生心肌和骨骼肌肌纤维毛细血管密度增加、Mb及血红蛋白水平提高等适应效应,改善心肌和骨骼肌供血供氧,提高心脏和骨骼肌有氧工作能力。
运动应激诱导PPARδ和PPARα基因表达上调促进心肌骨骼肌脂肪酸β氧化对有氧运动能力具有重要作用。线粒体脂肪酸β氧化供能是心肌有氧代谢的主要供能方式,占心肌耗能的70%以上,对于维持心泵功能具有重要作用[21]。PPARδ调节PPARα促进心肌细胞脂肪酸β氧化,提高心泵功能[22]。进行有氧耐力训练或在骨骼肌细胞过表达PPARδ都可以使PPARδ蛋白含量增加并获得相似的表型改变和骨骼肌重塑:慢肌纤维收缩蛋白基因和脂肪酸β氧化相关基因表达上调,氧化型肌纤维增多,氧代谢效率提高,有氧耐力水平提高。阻断PPARδ的作用,则不会产生上述改变[23]。
HR组差异表达的miRNA的功能主要是调控HIF-1a信号通路和PPAR信号通路对有氧训练应激的敏感性和响应强度,其综合适应效应与其运动适应表型特征是一致的。上调HIF-1a信号通路对低氧应激的敏感性可以使循环系统携氧能力和心肌/骨骼肌的供氧水平提高更显著。PPARδ和PPARα对运动应激诱导作用的反应性更好,可以更显著上调心肌/骨骼肌脂肪酸β氧化代谢能力,心肌/骨骼肌有氧工作能力提高更明显。心泵功能增强提高心输出量,骨骼肌有氧代谢效率提高,可以增加提高动静脉氧分压差。VO2max水平取决于心输出量和动静脉氧分压差[12],因此上调HIF-1a信号通路和PPAR信号通路都可以使VO2max的可训练性更好。同时,骨骼肌氧化型肌纤维增多,脂代谢供能比例增加还可以提高ILT水平[24],这与HR组具有较高的η(ILT)是一致的。
HR组miRNA差异表达谱特征显示出对多个应激适应通路的整合作用。差异表达的17条miRNA中,除miR210、miR423-5p和miR126专性调控低氧应激适应外,其余14条miRNA均同时作用于低氧应激适应通路和PPAR信号通路,在调控特点上显示出整合效应。本研究结果显示VO2max与ILT具有高度相关性,一方面证明了骨骼肌有氧代谢能力提高对VO2max的可训练性具有重要的影响[18],同时也提示我们miRNA对多种应激适应途径的整合作用可能是有氧耐力的心肺机能和骨骼肌代谢的协调一致的重要调节机制。
分阶段分析η(c-miRNA)与η(R-VO2max)和η(ILT)相关性结果表明:运动应激诱导的脂肪酸β氧化供能能力的提高对有氧耐力的可训练性十分重要。在阶段Ⅰ,η(miR183)与η(R-VO2max)和η(ILT)均呈显著负相关,miR183[25]负性调节HIF-3a[20]和CROT[23]基因表达,促进HIF-1a的功能,同时抑制线粒体脂肪酸β氧化,研究表明:低氧应激提高糖酵解供能水平,抑制脂肪酸β氧化[19]。该结果提示:训练过程中低氧应激对脂肪酸β氧化代谢功能的抑制作用不利于有氧耐力可训练性的提高。在阶段Ⅱ,η(miR141)和η(miR15a)均与η(R-VO2max)和η(ILT)正相关。研究表明:低氧应激诱导的HIF-2a[19]的高表达在应激结束后仍持续抑制PPARa和CPT1的表达和功能,损害线粒体脂肪酸β氧化能力[23],miR141[26]和miR15a[27]均可上调PPARa的表达,PPARa又上调CPT1的表达,该结果提示:抑制低氧应激对脂肪酸β氧化能力的损害有助于提高VO2max和ILT的可训练性。
此外,本研究结果还提示:机体抗损伤能力对于有氧耐力可训练性也有重要的影响。在阶段Ⅰ,η(R-VO2max)和η(c-miR133)呈负相关关系。miR133是肌肉特异性microRNA,运动过程中肌肉损伤漏出是其血浆水平升高主要原因[28],该结果提示:骨骼肌抗运动损伤能力越强,训练效果越好。
综上所述,本研究发现:在典型有氧训练周期的运动性适应过程中,17条c-miRNAs的表达特征与有氧耐力的可训练性之间存在密切的联系并初步建立了关联模型,同时也提出了一种采用表观遗传学指标进行运动员选材和指导训练的方法,以c-miRNAs表达特征作为生物标记评估运动能力的发展潜力和训练适应状态。进一步的研究将对c-miRNAs特征表达谱在运动员选材、训练效果/竞技状态评估等方面的应用价值做出更为全面和深入的探索。
注释:
① 该公式是根据同一受试者的R-VO2max直接测量值与12 min跑距离数值之间配对归纳得出的数值推算公式,反映的是R-VO2max与12 min跑距离之间的数值换算关系,如果采用公制单位该公式可以写成
R-VO2max(mL/(kg·min))=(s/1 609.34-0.318)/0.027 8
由于原始文献和相关文献中引用该公式时12 min跑距离数值均采用英制,所以本研究也采用相同的表述方式。该公式是数值推算公式,仅表示R-VO2max与12 min跑距离之间的数值换算关系,不能从公式中得出单位。
② “OD260/280值”是提取核酸后检验其纯度和完整性的指标,利用紫外分光光度计在260 nm和280 nm处的光密度比值判断核酸提取物的纯度和完整性,一般以OD260/280值在1.9~2.0范围内表示提取物的纯度和完整性较好,过高说明杂蛋白含量较高,过低说明RNA降解明显。
参考文献:
[1] 马力宏. 人类体质与运动能力的遗传学基础[J]. 天津体育学院学报,2001,16(4):1-5.
[2] 于红. 表观遗传学:生物细胞非编码RNA调控的研究进展[J]. 遗传,2009,31(11):1077-1086.
[3] 郑永霞,焦炳华. miRNA的生物形成及调控基因表达机制[J]. 生命的化学,2010,30(6):821-826.
[4] Leung A K L,Sharp P A. MicroRNA functions in stress responses[J]. Molecular Cell,2010,40(2):205-215.
[5] Williams A H,Liu N,Van Rooij E,et al. MicroRNA control of muscle development and disease[J]. Curr Opin Cell Biol,2009,21(4):461–469.
[6] Zhang C. MicroRNAs in vascular biology and vascular disease[J]. J Cardiovasc Transl Res,2010,3(3):235–240.
[7] Dang C V. Re-thinking the Warburg effect with Mycmicroma naging glutamine metabolism[J]. Cancer Res,2010,70(5):859–862.
[8] Kosaka N,Iguchi H,Ochiya T. Circulating microRNA in body fluid: a new potential biomarker for cancer diagnosis and prognosis[J]. Cancer Sci,2010,101(10):2087–2092.
[9] Baggish A L,Hale A,Weiner R B,et al. Dynamic regulation of circulating microRNA during acute exhaustive exercise and sustained aerobic exercise training[J]. J Physiol,2011,589(16):3983-3994.
[10] Davidsen P K,Gallagher I J,Hartman J W,et al. High responders to resistance exercise training demonstrate differential regulation of skeletal muscle microRNA expression[J]. J Appl Physiol,2011,110(2):309–317.
[11] 李义志,骆军保,汤长发. HSP1A1基因+190G>C多态与有氧运动能力表型指标的关联性研究[J]. 天津体育学院学报,2013,28(2):180-184.
[12] 李俊勇,任晋军,曹峰锐. 最大摄氧量、无氧阈和最大摄氧量平台同高校男生12 min跑和1 000 m跑的相关性分析[J]. 北京体育大学学报,2010,33(08):65-67.
[13] 林文弢,郑选梅,徐国琴,等. 不同氧浓度环境下个体乳酸阈的比较[J]. 广州体育学院学报,2012,32(6):81-84.
[14] Stegman H,Kindermann W,Schnabel A. Lactate kinetics and individual anaerobic threshold[J]. International Journal of Sports Medicine,1981,2(3):160-165.
[15] 杨若愚,王予彬,沈勋章,等. 基因多态性与杰出运动能力[J]. 中国组织工程研究,2014,18(7):1121-1128.
[16] 李江华,王智慧,朱小娟. 代谢组学运动选材与传统生化选材方法的比较[J]. 体育学刊,2013,20(1):123-128.
[17] Joshua T. Mendell Eric N. Olson. MicroRNAs in stress signaling and human disease[J]. Cell,2012,48(6):1172–1187.
[18] 王国军,温含,武雅琼. 跑节省化、最大摄氧量评价普通人有氧耐力的效果比较[J]. 中国组织工程研究,2013,17(7):1265-1272.
[19] Mounier R,Pialoux V,Schmitt L,et al. Effects of acute hypoxia tests on blood markers in high-level endurance athletes[J]. Eur J Appl Physiol.,2009,106(5):713-720.
[20] 赵琛,谭斐. 低氧诱导因子-1激活的调节[J]. 生命科学,2013,25(1):40-46.
[21] Cheng L,Ding G,Qin Q,et al. Cardiomyocyte-restricted peroxisome proliferator-activated receptor-delta deletion perturbs myocardial fatty acid oxidation and leads to cardiomyopathy[J]. Nat Med,2004,10(11):1245-1250.
[22] Barger P M,Brandt J M,Leone T C,et al. Deactivation of peroxisome proliferator-activated receptor αduring cardiac hypertorphic growth[J]. J Clin Invest,2000,105(12):1723-1730.
[23] 胡琴,李隆贵. 心肌脂肪酸氧化酶的基因调控机制及PPARα的作用[J]. 中国病理生理杂志,2002,18(12):1552-1555.
[24] 刘昭强,朱寄天. 心肌乳酸代谢及乳酸对心脏功能的影响[J]. 成都体育学院学报,1999,25(3):11-16.
[25] Vohwinkel C U,Lecuona E,Sun H,et al. Elevated CO(2) levels cause mitochondrial dysfunction and impair cell proliferation[J]. J Biol Chem,2011,286(43):37067-37076.
[26] Mangala L S,Zhang Y,He Z,et al. Effects of simulated microgravity on expression profile of microRNA in human lymphoblastoid cells[J]. J Biol Chem,2011,286(37):32483-32490.
[27] Sun L L,Jiang B G,LI W T,et al. MicroRNA-15a positively regulates insulin synthesis by inhibiting uncoupling protein-2 expression[J]. Diabetes Res Clin Pract,2011,91(1):94-100.
[28] Chen J F,Mandel E M,Thomson J M,et al. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation[J]. Nat Genet,2006,38(2):228-233.
猜你喜欢
中老年保健(2022年3期)2022-11-21
中老年保健(2022年2期)2022-08-24
中老年保健(2022年4期)2022-08-22
天津医科大学学报(2021年3期)2021-07-21
国外医药(抗生素分册)(2016年5期)2016-07-12
中国运动医学杂志(2016年3期)2016-07-10
中国运动医学杂志(2016年3期)2016-07-10
医学研究杂志(2015年5期)2015-06-10
中国体育教练员(2015年2期)2015-04-16
实验动物与比较医学(2014年5期)2014-02-28
