
野栽渗入系水稻籽粒储藏蛋白质含量的QTL遗传解析
2015-07-31 17:20赵琳琳李楠吕志伟等
江苏农业科学 2015年3期
赵琳琳 李楠 吕志伟等


摘要:以云南元江普通野生稻为供体亲本、以优良籼稻品种特青为受体亲本构建了高代回交群体(BC3)。采用全自动凯氏定氮仪测定糙米中的总氮含量,用总氮含量乘以5.95估算储藏的蛋白质含量,蛋白质含量采用Map Manager QTXb17软件进行数量性状基因座(quantitative trait locus,QTL)定位分析,利用QTL检测分析2次测定的储存蛋白,结果共发现14个对蛋白质含量有较高相关性的QTL。在第1号染色体的RM272、RM243、RM23、RM5、RM212、RM306位点和第2号染色体的RM250位点附近发现了7个能提高蛋白质含量的QTL,它们均来源于野生稻;另外7个QTL位于第6、7、8、10号染色体上,分别是第6号染色体的RM345、第7号染色体的RM295、RM82、RM481、RM172,第8号染色体的RM25,以及第10号染色体的RM258附近的QTL,它们来源于栽培稻亲本特青的等位基因作用表现为提高蛋白质含量。在检测到的QTL中,有6个QTL在2次分析中均被检测到,经分析是较稳定的低蛋白QTL,它们分别为RM243、RM23、RM5、RM295、RM82、RM258。
关键词:野生稻基因渗入系;籽粒储藏蛋白质;凯氏定氮法;QTL分析
中图分类号: Q946.1 文献标志码: A
文章编号:1002-1302(2015)03-0050-03
水稻是一种重要的粮食作物,世界上约50%的人口以稻米为主食,稻米蛋白质被称作优质植物蛋白[1],通过与其他谷物进行比较可知,水稻中必需氨基酸的含量大体上要高于联合国粮食及农业组织(FAO)/世界卫生组织(WHO)建议的标准水平[2],其第一限制性氨基酸赖氨酸含量高于其他谷类,是人类蛋白质的主要来源之一。在发展中国家,约60%的蛋白来源于谷物,在未来的几十年里,谷物仍是世界上2/3人口的主要蛋白来源[3]。因此,提高稻米蛋白质含量已经成为当今水稻品质育种的一个重要内容;同时,随着慢性肾功能不全患者不断增加,培育低蛋白含量,特别是培育低可消化吸收蛋白(谷蛋白)含量水稻品种已成为功能水稻的新育种目标。
籽粒储存蛋白受多基因控制,属于数量遗传性状,开展控制籽粒储存蛋白含量的数量性状基因座(quantitative trait locus,QTL)定位与克隆,不仅对水稻种质资源创新和高蛋白米、低蛋白米的新品种选育有重要的理论意义,而且对当前调整农业结构、保持农业可持续发展同样具有重要的现实意义。
迄今为止,大量的研究主要是对水稻农艺性状的遗传分析和基因定位,并且对稻米品质性状的遗传分析和基因定位研究主要集中于加工品质、外观品质和蒸煮及食味品质等性状上[4-9],而关于水稻糙米储存蛋白定位的报道并不多见,仅限于个别报道。于永红等利用协青早B/密阳46的重组自交系群体对水稻糙米蛋白质含量的QTL分析表明,5个控制蛋白质含量的QTLs(qPc-3、qPc-4、qPc-5、qPc-6、qPc-10)均被检测到,单个QTL对群体表型变异的贡献率为4.17%~19.14%;其中第6号染色体的Wx基因区域对蛋白质含量具有主效作用[10]。Li等利用亚洲栽培稻和非洲栽培稻的回交高代群体,在第8号染色体上定位到了1个QTL[11]。Yoshida等利用DH群体(由花药单倍体诱导得到)定位到了分别位于第 8、9、11、12号染色体上的4个控制稻米蛋白含量的QTLs,其中第11号染色体RM206位点附近的QTL效应较大[12]。Aluko等利用非洲野生稻与籼稻BC3F1回交构建的DH群体,分别在第1、2、6、11号染色体上检测到4个与蛋白质含量相关的QTLs,其中位于第1号染色体RM226至RM297位点之间的QTL贡献率较大[13]。张涛等检测到6个QTLs (qPc-3、qPc-6、qPc-7、qPc-8-1、qPc-8-2、qPc-11)对糙米蛋白质含量进行控制,它们分别位于第3、6、7、8、11号染色体上,单个QTL对群体表型变异的贡献率为379%~19.41%[14]。在这些QTL的区间中,第8染色体的基因区域对糙米蛋白质含量具有主效作用。李晨等利用BC4群体共定位到了6个糙米蛋白QTLs,分别位于第1、2、8、9号染色体上,其中位于第1号染色体RM472位点附近的QTL具有主效作用[15]。
野生稻中蕴含着丰富的遗传信息,是栽培稻的祖先,因此发掘野生稻中的有益基因不仅有利于解决当前育种资源狭窄的现状,而且是突破育种瓶颈的有效方法之一。但是,目前关于从野生稻中定位QTL的报道并不多,各个方面都待完善,因此今后需要进一步强化这方面的工作。本研究通过采用凯氏定氮法对野生稻染色体片段代换系(简称水稻野栽渗入系)的籽粒储存蛋白质含量进行分析,并对控制籽粒储存蛋白质含量的QTL进行了检测。
1 材料与方法
1.1 群体构建
本研究所构建的野栽渗入系由中国农业大学孙传清博士提供,是以元江普野为供体亲本、籼稻品种特青为受体亲本培育而成的。在BC3F2群体的基础上,连续自交3次得到BC3F3、BC3F4群体。根据BC3F2株系表型鉴定和基因分析结果,渗入系构建的基本材料选取了263个BC3F3、BC3F4单株,最终渗入系由106个BC3F3、BC3F4候选单株构成。
1.2 SSR引物扩增及数据处理
根据Temnykh等发表的水稻SSR序列合成引物,用在亲本间有多态性的112个SSR标记,调查上述野栽渗入系群体106个系的基因型[16]。在基因型分析中,“B”表示与亲本特青带型相同,“A”表示与野生稻亲本带型相同,“H”则表示具杂合带型,“-”表示带型模糊或由于某种原因缺失。采用MAPMAKER/EXP3.0软件并参考Temnykh等发表的水稻SSR连锁图谱为框架,构建连锁图谱,可见112个SSR标记分布于12条染色体上,各染色体上最多的有13个标记,最少的有7个标记,平均每条染色体有9.3个标记,标记平均间距17.8 cM,基本能满足QTL定位需要[16]。
1.3 糙米储藏蛋白质含量的测定及QTL分析
选用2006年于北京采收的种子,挑选表型一致(如大小、颜色)的水稻籽粒,剥去颖壳后于干燥箱(105 ℃)中干燥5 h,然后以8粒种子为1组称质量,每个家系重复5次。采用意大利产全自动凯氏定氮仪(UDK 142)测定糙米中的总氮含量,用总氮含量乘以5.95估算储藏的蛋白质含量[17-18]。为了确保测定结果的准确性,同时选用2007年采收于北京的种子进行同样的测定,重复3次。
采用Map Manager QTXb17软件,取P<0.05作为判断QTL存在的阈值,并用单位点分析法对籽粒储藏蛋白质含量进行QTL定位与遗传分析[19]。
2 结果与分析
2.1 籽粒储藏蛋白质含量的分布
图1是不同家系的蛋白质含量分布情况。可见渗入系群体的蛋白质含量趋于正态分布,符合数量性状的特征,适于QTL定位分析。
2.2 QTL解析
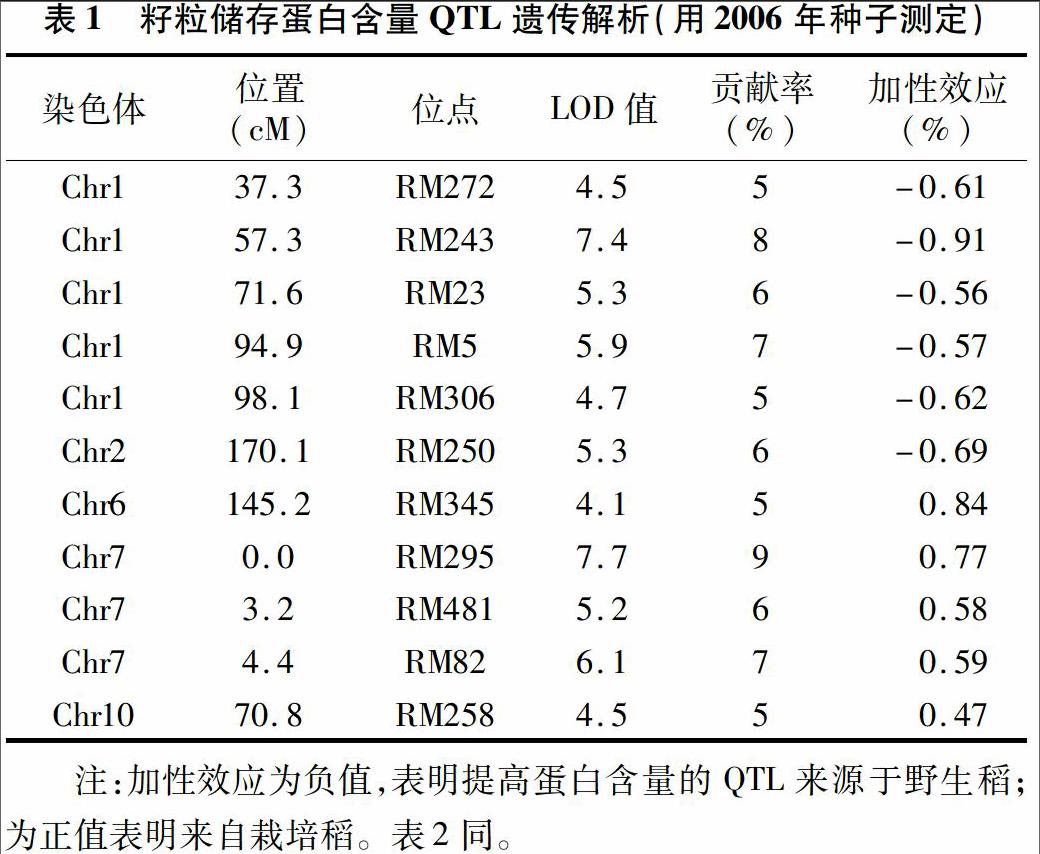
将渗入系群体的籽粒蛋白质含量数据采用Map Manager QTXb17软件进行QTL定位分析,共检测到11个与蛋白质含量相关的QTL(表1)。其中位于第1号染色体RM272、RM243、RM23、RM5、RM306位点附近以及位于第2号染色体RM250位点附近的计6个QTL,贡献率分别为5%、8%、6%、7%、5%、6%,加性效应分别为-0.61%、-0.91%、-0.56%、-0.57%、-0.62%、-0.69%,来源于野生稻的等位基因能够提高糙米籽粒储藏蛋白质含量。另外5个QTL位于第6、7、10号染色体上,分别是第6号染色体RM345位点,第7染色体RM295、RM481、RM82位点,以及第10染色体RM258位点附近的QTL,贡献率分别为5%、9%、6%、7%、5%,加性效应值分别为084%、0.77%、0.58%、0.59%、047%,来源于特青的等位基因表现为提高糙米籽粒储藏蛋白质含量。
2007年批次种子的重复试验结果见表2,共检测到9个与蛋白质含量相关的QTL。分别位于第1、7、8、10号染色体上,贡献率为5%~10%。与表1相比可知,在第2、6号染色体上没有检测到QTL,而在第8号染色体上检测到的来源于特青的、提高蛋白质含量的QTL在第1次的结果中并没有检测到。
2次结果均检测到的QTL位点见表3,可见有位于第1号染色体的RM243、RM23、RM5位点附近的3个QTL,表现为野生稻等位基因提高糙米籽粒储藏蛋白质含量;还有位于第7号染色体RM295、RM82位点和第10号染色体RM258位点附近的3个QTL,表现为特青等位基因提高糙米籽粒储藏蛋白质含量。
3 结论与讨论
本研究以元江普野为供体亲本、籼稻品种特青为受体亲本构建的野栽渗入系群体的种子为材料,在全自动凯氏定氮仪中消化并测定剥掉颖壳后糙米的含氮量,用含氮量乘以595作为糙米蛋白质含量的计算指标。用Map Manager QTXb17软件,取P<0.05作为判断QTL存在的阈值,并用单位点分析法,对籽粒储藏蛋白质含量进行QTL定位与遗传分析,2次共定位到14个QTL位点。其中来自野生稻亲本的高蛋白QTL有7个,分别分布于第1、2号染色体上,第1号染色体上有6个,第2号染色体上有1个。来源于栽培稻亲本特青的高蛋白QTL也有7个,分别分布于第6、7、8、10号染色体上,第7号染色体上有4个,其余染色体上各有1个。
通过与以前的研究结果相比较,我们发现有5 个QTL位点与前人所报道的QTL位点在同一区域或者在相临区域。位于第1号染色体上RM23位点附近的QTL与李晨等报道的qCP1-1[15]在同一区域。位于第1号染色体上RM212位点附近的QTL,以及位于第2号染色体上RM250位点附近的QTL分别与Aluko等报道的Pro1、Pro2[13]在同一区域。位于第7号染色体RM172位点附近的QTL与吴长明等报道的Pr3[20]处于相临区域。位于第8号染色体RM25位点附件的QTL与张涛等报道的qPc-8-2[14]在相近区域,经分析是同一位点。
此外,经过分析有9个QTL位点与以前报道的位置不一样,是检测到的新QTL;这9个QTL位点分别位于第1号染色体RM272、RM243、RM5、RM306位点附近,第6号染色体RM345位点附近,第7号染色体RM295、RM481、RM82位点附近,以及位于第10号染色体RM258位点附近。
在本研究中,没有检测到主效QTL,检测到的QTL贡献率最大的为10%,这一结论与Li等的研究结果[11,21-22]类似,而与于永红等的研究结果[10,12-15,23]有所差异,认为这种差异可能是由于所用的材料不同造成的。
本研究共从野生稻中检测到7个控制籽粒储存蛋白质的QTL,其中在第1号染色体上有6个,在第2号染色体上有1个,这为今后进一步研究分析和利用这些QTL打下了基础。
参考文献:
[1]周丽慧,刘巧泉,张昌泉,等. 水稻种子蛋白质含量及组分在品种间的变异与分布[J]. 作物学报,2009,35(5):884-891.
[2]王治平. 大米品质与主食米饭食用品质相关性的研究[D]. 长沙:湖南农业大学,2000:2-4.
[3]姚惠源. 稻米深加工[M]. 北京:化学工业出版社,2004.
[4]Tan Y F,Li J X,Yu S B,et al. The three important traits for cooking and eating quality of rice grains are controlled by a single locus in an elite rice hybrid,Shanyou 63[J]. Theoretical and Applied Genetics,1999,99(3/4):642-648.
