
饲粮精粗比对犊牛组织中基因GLUT1 和FATP4 及PepT1 mRNA 表达的影响
2015-07-15 05:47:12占今舜杨宏波詹康刘红刘明美赵国琦
湖南农业大学学报(自然科学版) 2015年5期
占今舜,杨宏波,詹康,刘红,刘明美,赵国琦
(扬州大学动物科学技术学院,江苏 扬州 225009)
豆科牧草紫花苜蓿在世界上广泛分布,且享有盛誉,被称为“牧草之王”。紫花苜蓿含有蛋白质等营养物质,是较好的饲料原料之一[1–2]。在西方,小黑麦草是主要的饲草之一;在中国,随着农业结构的调整,小黑麦草也被作为饲料作物进行开发。饲用小黑麦草具有相对较强的抗逆性,能够于干旱瘠薄地、闲散废弃地生长,对畜牧业的发展具有一定的促进作用[3]。
葡萄糖转运蛋白(GLUTs)是细胞转运葡萄糖的载体,介导葡萄糖在细胞膜两侧的转运,广泛分布于体内各种组织。GLUT1 是一种易化扩散的葡萄糖转运蛋白,分布广泛,具有维持基础状态下组织细胞葡萄糖摄入的功能。GLUT1 是反刍动物乳腺中主要的葡萄糖转运蛋白[4–6]。细胞膜脂质和细胞内部多种信号分子等均由脂肪酸构成,而脂肪酸转运蛋白(FATP)是促进脂肪酸转运的蛋白之一。FATP4 是动物肠道中唯一存在的FATPs家族成员,存在于肠绒毛细胞,被认为是调控肠道细胞吸收脂肪酸的重要因子[7–8]。小肽转运载体(PepT) 在肠道内参与二肽和三肽的跨膜转运。组织和细胞中的PepT1 通过跨膜转运肽类物质,对营养物质的吸收具有重要作用[9–10]。
饲料营养水平对葡萄糖、脂肪和小肽转运蛋白具有调控作用。笔者将质量比3∶1 的苜蓿和小黑麦草制成不同精粗比的颗粒全价饲料,研究其对犊牛不同组织GLUT1、FATP4、PepT1 基因的影响,以从分子水平探明不同营养水平对动物营养物质吸收的影响。
1 材料与方法
1.1 试验日粮和犊牛
根据NRC(2001)营养需要,采用20 型颗粒饲料机组,将质量比3∶1 的苜蓿和小黑麦草分别制成精粗比75∶25、70∶30、65∶35 和60∶40 的颗粒全价饲料。
将日龄(104.00±1.50) d、平均体重(121.25± 4.12) kg 的中国荷斯坦断奶公犊牛12 头分为4组,分别饲喂精粗比75∶25、70∶30、65∶35、60∶40 的颗粒全价饲料(分别称其为试验Ⅰ组、Ⅱ组、Ⅲ组、Ⅳ组),预试期14 d,正试期 56 d。每天饲喂3次,饲喂时间为8:30、14:30 和20:30,每周调整1次饲喂量,但保证每组犊牛的干物质采食量基本一致。犊牛自由饮水,每天有6~7 h户外活动。每日清晨打扫圈舍,每周至少对牛舍消毒2次。各试验组饲粮的组成和营养水平见表1。

表1 饲粮组成和营养水平(干物质基础) Table 1 Composition and nutrient levels of basal diets (DM basis)
1.2 方法
试验于2014年3月至2014年5月在扬州大学实验农牧场进行。
1.2.1 样品采集
于试验第70天,每组犊牛禁食12 h 后进行屠宰,沿纵向剖开腹腔,分离出肝脏、瘤胃、十二指肠和肌肉。用高温灭菌后的镊子和手术剪刀剪取各组织,在灭菌生理盐水中清洗,装入1.5mL 的离心管中,液氮速冻后转移到–80℃的冰箱冷冻保存。
1.2.2 组织总RNA 的提取和cDNA 的合成
根据RNA 提取试剂盒(天根生化科技有限公司)说明书进行组织总RNA 的提取,采用One Drop 仪测定总RNA 的浓度和纯度。根据1%琼脂糖凝胶电泳结果判断RNA 是否有降解。采用Takara 反转录试剂盒进行cDNA 合成。试验在冰上进行。反应体系10 μL。反应条件:37℃反转入反应15min,85℃反转入酶失活反应5 s;4℃保存。所得cDNA 保存于–20℃。
1.2.3 引物设计
根据Genbank 中的GAPDH、GLUT1、FATP4、PepT1 基因的序列,采用Primer 5.0 软件设计。引物(表2)由Invitrogen 公司合成。

表 2 设计引物 Table 2 Primer design
1.2.4 Real Time–PCR 反应条件
根据TaKaRa 试剂盒说明书配制反应液(表3)。在罗氏Light Cycler® 96 PCR 仪上进行检测。条件:95℃预变性30 s;95℃变性5 s,60℃退火20 s,40个循环;65℃延伸15 s。RT–PCR 反应液总体积20 μL,其中, SYBR 预混合试剂Ex TaqⅡ(2×)10 μL(终浓度1×);10 μmol/L PCR 正向引物0.8 μL(终浓度0.4 μmol/L),10 μmol/L PCR 反向引物0.8 μL(终浓度0.4 μmol/L),cDNA 模板2 μL,灭菌蒸馏水6.4 μL。
1.3 数据分析
先用Excle 2007 对试验数据进行预处理,根据公式2–△Ct计算出目的基因的相对表达量,其中△Ct=Ct目的基因-Ct管家基因。采用SPSS 17.0 进行单因素方差分析,用LSD 法进行多重比较。
2 结果与分析
2.1 各试验组犊牛不同组织中GLUT1 mRNA 的相对表达量
从表3 中可以看出,试验Ⅲ组犊牛肝脏、瘤胃、十二指肠和盲肠的GLUT1 mRNA 相对表达量显著高于试验Ⅳ,试验I组皱胃的GLUT1 mRNA 相对表达量极显著高于试验Ⅲ和Ⅳ组。

表3 不同组织中GLUT1 mRNA 的相对表达量 Table 3 Relative expression levels of GLUT1 mRNA in different tissues
2.2 各试验组犊牛不同组织中FATP4 mRNA 的相对表达量
由表4 可见,试验Ⅲ组犊牛肝脏中FATP4 mRNA 的相对表达量极显著高于试验Ⅳ组,显著高于试验I 和Ⅱ组;试验I组瘤胃和十二指肠中FATP4 mRNA 的相对表达量显著高于试验Ⅲ组;试验I组皱胃中FATP4 mRNA 的相对表达量极显著高于试验Ⅳ组;试验Ⅲ组盲肠中FATP4 mRNA 的相对表达量显著高于其他各组。
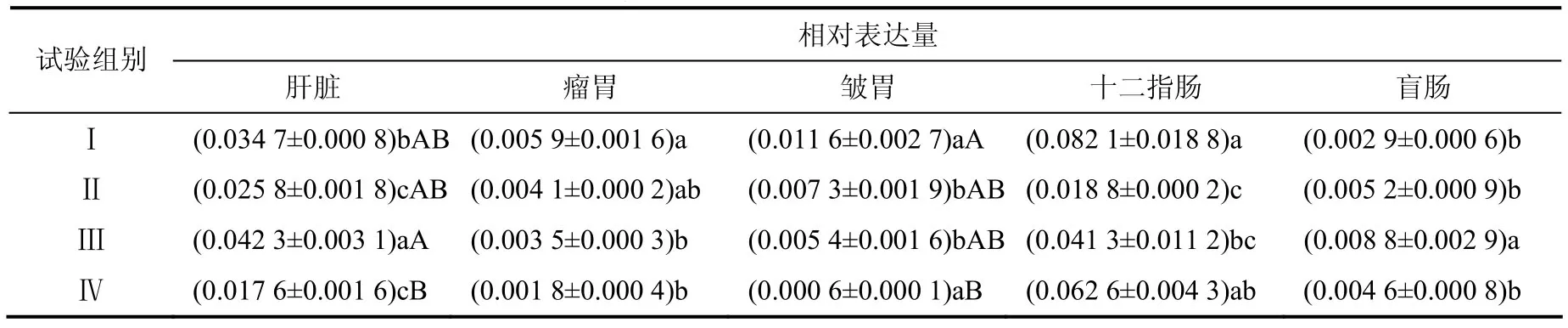
表4 不同组织中FATP4 mRNA 的相对表达量 Table 4 Relative expression levels of FATP4 mRNA in different tissues
2.3 各试验组犊牛不同组织中PepT1 mRNA 的相对表达量
从表5 中可以看出,试验Ⅱ和Ⅳ组犊牛肝脏中PepT1 mRNA 的相对表达量显著高于试验Ⅰ和Ⅲ组,试验I 和Ⅳ组瘤胃中PepT1 mRNA 的相对表达量显著高于试验Ⅱ和Ⅲ组,试验I组皱胃和十二指肠中PepT1 mRNA 的相对表达量显著高于试验Ⅳ组,试验Ⅲ组盲肠中PepT1 mRNA 的相对表达量显著高于试验I 和Ⅳ组。
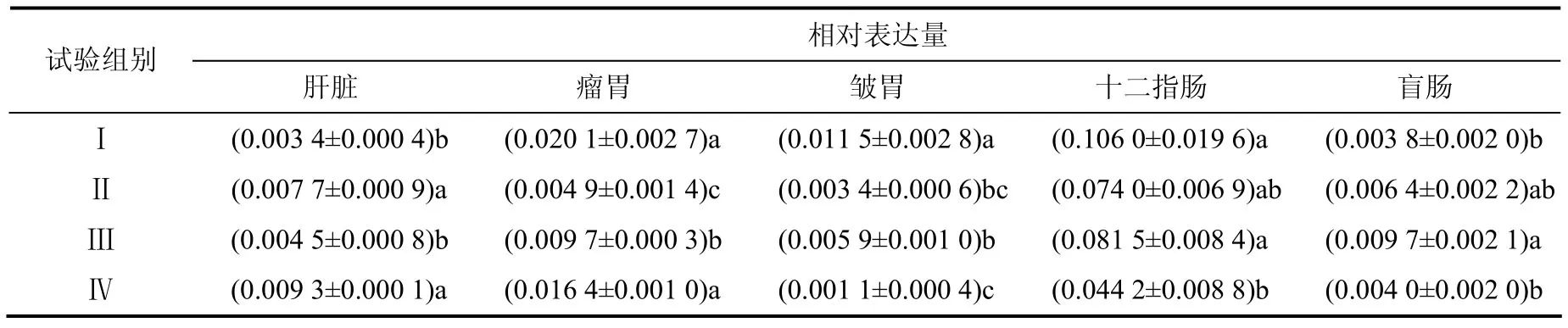
表5 不同犊牛组织中PepT1 mRNA 的相对表达量 Table 5 Relative expression levels of PepT1 mRNA in different tissues
3 结论与讨论
不同精粗比饲粮显著或极显著地影响着绵羊瘤胃纤维素酶的活性[11]。采食精粗比20∶80、30∶70、和40∶60 的日粮,小尾寒羊纤维素和半纤维素消化量呈先上升后下降的趋势[12],说明适当的精粗比日粮可以促进纤维素的分解,进而提高葡萄糖的生成。潘巧等[13]发现,饲喂高精料日粮,奶山羊门静脉吸收丙酸的量增加,导致肝脏增强吸收和利用丙酸,使肝脏糖异生相关酶基因的表达上调。当细胞外葡萄糖浓度增加时,细胞膜上GLUT1 的表达增加[14–15]。本试验结果表明,除皱胃外,试验Ⅲ组各组织中GLUT1 mRNA 的相对表达量最高,说明精粗比为65∶35 的日粮更利于犊牛吸收葡萄糖。试验Ⅳ组皱胃的GLUT1 mRNA 相对表达量低于其他各组,可能是因为高精料日粮中淀粉的含量高,瘤胃淀粉分解菌占优势,淀粉被大量降解,未被瘤胃降解的部分淀粉则在皱胃中被消化酶分解成葡萄糖,导致GLUT1 基因表达升高,促进葡萄糖的吸收。
给COS1 细胞转染FATP4 cDNA 导致棕榈酰– CoA 合成酶和二十四烷基–CoA 合成酶升高[16],表明FATP4 基因能够编码酰–CoA合成酶,说明FATP4在长链脂肪酸的摄取和代谢调节中发挥着重要作用。长链脂肪酸的合成原料是乙酰辅酶A,而反刍动物脂肪酸的合成是由瘤胃中形成的乙酸和丁酸转化为乙酰辅酶A 和丁酰辅酶A 的。随精料水平的增加,瘤胃乙酸、丁酸含量降低,丙酸含量增加[17];随采食时间的延长,挥发性脂肪酸含量呈先增加后降低趋势。随精粗比的升高,奶牛瘤胃的乙酸、丁酸含量降低[18]。本试验中,试验Ⅲ组肝脏FATP4基因的表达高于其他各组,可能是因为该组日粮使瘤胃产生的乙酸、丁酸含量高,导致肝脏合成脂肪酸多,促进肝脏对脂肪酸的转运和代谢。饲料中的脂肪在瘤胃微生物作用下水解产生甘油和各种脂肪酸。在本试验中,试验I组瘤胃、皱胃和十二指肠FATP4 基因的表达量最高,这可能是因为该组日粮中含量较高的蛋白和脂肪在瘤胃微生物的作用下大量合成脂肪酸。
在本试验中,试验I组的日粮蛋白质水平最高,因此,在瘤胃降解成的小肽较多,进而导致瘤胃、皱胃和十二指肠的PepT1 mRNA 相对表达量最高。盲肠是反刍动物微生物消化的另一主要场所。王东升[19]等发现,高精粗比能显著提高山羊盲肠内容物和盲肠静脉的生物胺含量。本试验中,试验I组PepT1 基因的表达量最低,可能是由盲肠中产生的大量生物胺导致小肽生成较少引起。张树坤等[20]发现,高精料饲喂泌乳期山羊,其肝脏中氨基酸消耗增多,同时小肽合成减少。本试验中试验I组肝脏的PepT1 mRNA 的相对表达量最低,可能是因为高精料引起肝脏合成小肽减少导致的。
综合分析结果表明,精粗比为65∶35 的全价颗粒饲料能促进消化道GLUT1 基因的表达,精粗比为75∶25 的全价颗粒饲料能促进消化道FATP4、PepT1 基因的表达。从整体上看,精粗比为65∶35的全价颗粒饲料更有利于犊牛对葡萄糖、脂肪酸和蛋白质的吸收。
[1] 占今舜,陈佩佩,张彬.苜蓿在养猪生产上的应用研究进展[J].养猪,2012(6):27–30.
[2] 卢欣石.中国苜蓿产业发展问题[J].中国草地学报,2013,35(5):1–5.
[3] 谢楠,李源,赵海明,等.饲用黑麦、小黑麦品种的抗旱性评价[J].中国草地学报,2011,33(6):82–88.
[4] 刘越素,任跃忠.葡萄糖转运蛋白–1 与糖尿病肾病[J].国外医学泌尿系统分册,2004,24(3):388–391.
[5] Christina P,Alexandra C,Jordan S,et al.Effects of cinnamaldehyde on theglucose transport actⅣity of GLUT1[J].Biochimie,2011,93:339–344.
[6] 齐利枝,闫素梅,生冉,等.奶牛乳腺中乳成分前体物对乳成分合成影响的研究进展[J].动物营养学报,2011,23(12):2077–2083.
[7] 齐仁立,黄金秀,杨飞云,等.脂肪酸转运蛋白家族及其介导的脂肪酸跨膜转运[J].动物营养学报,2013,25(5):905–911.
[8] Blackburn C,Guan B,Brown J,et al.Identification and characterization of 4–aryl–3 , 4–dihydropyrimidin–2 (1H)–ones as inhibitors of the fatty acid transporter FATP4[J].Bioorganic & Medicinal Chemistry Letters,2006,16:3504–3509.
[9] Fei Y J,Sugawara M,Liu J C,et al.cDNA structure,genomic organization,and promoter analysis of the mouse intestinal peptide transporter PEPT1[J]. Biochimica et Biophysica Acta,2000,1492:145–154.
[10] 朱宇旌,王秉玉,张勇,等.小肽转运载体1 的生物学特性及其功能[J].动物营养学报,2012,24(10):1847–1853.
[11] 霍鲜鲜,侯先志,赵志恭,等.不同精粗比日粮对绵羊瘤胃内纤维素酶活的影响[J].甘肃畜牧兽医,2003(5):16–20.
[12] 门小明,雒秋江,唐志高,等.3 种不同精粗比日粮条件下空怀小尾寒羊母羊的消化与代谢[J].中国畜牧兽医,2006,33(10):13–16.
[13] 潘巧.日粮精粗比对奶山羊肝脏糖异生的影响及其机理研究[D].南京:南京农业大学,2012.
[14] 丁红,马健飞,樊怡,等.高糖对大鼠腹膜间皮细胞葡萄糖转运蛋白1 调节作用的研究[J].中国医科大学学报,2006,35(2):151–152.
[15] 任蕾.高血糖对大鼠肾皮质葡萄糖转运蛋白l mRNA表达的影响[D].沈阳:中国医科大学,2008.
[16] Thomas H,Florian B,Isabella G,et al.Mouse fatty acid transport protein 4 (FATP4): Characterization of thegene and functional assessment as a very long chain acyl–CoA synthetase[J].Gene,2001,270:31–40.
[17] 华金玲,郭亮,王立克,等.不同精粗比日粮对黄淮白山羊瘤胃挥发性脂肪酸影响[J].东北农业大学学报,2013,44(6):58–61.
[18] 贾玉东,王振勇,柴同杰,等.日粮粗精比对奶牛瘤胃液和血清乙酸、丙酸、丁酸的影响[J].西北农林科技大学学报:自然科学版,2008,36(7):27–32.
[19] 王东升.日粮模式对瘤胃与后肠中生物胺生成的影响[D].南京:南京农业大学,2012.
[20] 张树坤,姜雪元,倪迎冬,等.不同精粗比日粮对泌乳山羊肝脏蛋白质代谢及乳蛋白含量的影响[J].农业生物技术学报,2013,21(4):413–420.
猜你喜欢
畜牧与兽医(2021年8期)2021-08-06 02:58:34
现代畜牧科技(2021年5期)2021-07-20 08:07:48
中国兽医杂志(2020年3期)2020-09-07 08:25:04
今日畜牧兽医(2020年6期)2020-02-15 12:23:15
饲料博览(2017年8期)2017-04-04 12:11:59
兽医导刊(2016年12期)2016-05-17 03:51:34
兽医导刊(2016年12期)2016-05-17 03:51:15
动物营养学报(2015年9期)2016-01-07 11:29:38
当代畜禽养殖业(2014年5期)2014-08-31 02:50:52
云南畜牧兽医(2014年4期)2014-02-28 21:25:32
