
青藏高原草地双子叶植物叶片的气孔特征研究
2015-07-05 07:03:24胡选萍吉成均安丽华
西北植物学报 2015年7期
胡选萍,吉成均,安丽华
(1 陕西理工学院生物科学与工程学院,陕西汉中723000;2 北京大学城市与环境学院,北京100871;3 北京大学医药卫生分析中心,北京100191)
青藏高原平均海拔4 000m 以上,具有太阳辐射强、日照时间长、气温低、昼夜温差大、气压低、氧气与二氧化碳含量少、湿度和降水量不均衡、风力大、气候频变等独特的高原气候特点[1-2]。植物能在如此极端的环境中生存,一定有其特殊的结构特征及生理、生态适应机制[3-7]。叶片作为植物暴露在环境中面积最大的器官,对外界环境的变化敏感,最易受环境影响,且能快速反映环境的变化,因而已成为植物生态解剖研究的主要指标之一[8-9]。前人已经总结出青藏高原草地植物叶解剖特征的一系列规律:如叶表皮细胞和角质层较厚,且有一定程度的角质化,栅栏组织多层,叶肉细胞排列较紧密,异细胞丰富,具有发达的通气组织,许多植物还发育出复杂的表皮毛及各种附属物等[10-13]。气孔作为植物表皮的特殊结构,是植物与外界环境进行气体交换的门户,也是植物体内水分蒸腾散失的通道,易受环境条件的修饰,其形状、大小、分布以及气孔保卫细胞厚度等对外界环境变化较为敏感,因此用植物气孔参数反映植物对气候环境变化的响应,已成为近年来国际生态学研究的热点。目前关于高寒极端环境下气孔形态、分布及相关数量特征的研究很少,区域尺度上气孔特征的适应性变化还没有系统报道。本研究通过对青藏高原草地主要双子叶植物气孔特征的观察,试图诠释草地植物气孔在高原高寒缺氧环境下形态结构及分布特点,分析青藏高原草地植物气孔数量特征的共性规律,揭示青藏高原草地植物对高原极端严酷环境的内在适应机制,对于了解植物对环境的响应和适应关系,以及探索植物气孔参数作为辅助分类依据提供参考。
1 材料和方法
1.1 实验材料
实验材料由北京大学城市与环境学院提供,样品于2003~2005年采至青藏高原,用FAA 溶液固定。这些草地植物隶属于9个科(蝶形花科、菊科、蔷薇科、蓼科、玄参科、罂粟科、龙胆科、瑞香科、十字花科),12个属(棘豆属、黄耆属、凤毛菊属、橐吾属、蒿属、委陵菜属、蓼属、马先蒿属、绿绒蒿属、龙胆属、狼毒属、燥原芥属),29个种(表1)。
1.2 实验方法
1.2.1 样品制备 用10%铬酸对叶片离析12h,清水冲洗并浸泡,采用透明胶带粘取法制备装片,在光学显微镜下观测叶片表皮气孔特征。透明胶带为普通型胶带,宽度13 mm,厚度0.05 mm。将胶带拉开8~10cm,胶面朝上平放在实验台面上。将叶片从清水中取出,用吸水纸迅速吸干表面水分。用镊子夹住材料,分清上下表面,将下表面顺长贴于胶带约1/3处,再将胶带从1/2处对折,平整地粘贴于材料的上表面,用镊子柄沿同方向轻刮数下,使胶带与叶片两面充分粘着。从对折处将胶带剪断,将胶带缓慢撕开,表皮已粘附于胶带之上。分清上下表皮胶面,用清水冲洗除去附在表皮上的叶肉细胞残留物。用吸水纸吸干表面水分,将粘有上下表皮的胶带成对贴于载玻片上,封片后用MoticB5光学显微镜观察,Motic Images Plus 2.0显微成像系统照相和测定。
1.2.2 观察测量指标 气孔长度(SL)测定:每片叶在显微镜下随机选取10个视野,每个视野随机选取5个气孔,测量其保卫细胞长度,即为气孔长度(μm)。

表1 实验材料来源Table 1 Sources of materials
续表1 Continued Table 1
气孔密度(SD)测定:每片叶在显微镜下随机选取10个视野计算气孔数目,并测量每个视野的面积,将其平均值换算为单位面积的气孔数,即为叶片的气孔密度(个/mm2)。
气孔指数(SI)测定:随机观察10个视野,统计单位视野气孔个数(A)和单位视野表皮细胞个数(B),计算气孔指数。气孔指数(%)=[A/(A+B)]×100%。
1.2.3 统计分析方法 采用SPSS19.0统计软件对实验数据进行统计分析。对叶片上、下表皮气孔数量指标进行one-way ANOVA 分析与Pearson相关分析。
2 结果与分析
2.1 青藏高原草地双子叶植物叶片气孔形态特征
青藏高原草地双子叶植物长期受高原气候环境的筛选,在气孔分布及形态等方面形成了一些特有的共性特征:(1)从气孔分布来看,气孔多随机排布,没有固定方向,且气孔在叶片上下两面均有分布。实验观察的29种双子叶植物,除小叶金露梅(Potentilla parvifolia)上表皮没有观察到明显可见气孔,银 背 凤 毛 菊(Saussurea nivea)、禾 叶 凤 毛 菊(Saussurea graminifolia)下表皮没有观察到气孔,其他26 种植物上下表皮均有气孔分布(图版Ⅰ、Ⅱ)。(2)从气孔形态来看,气孔呈长椭圆形或宽椭圆形,保卫细胞呈肾形,排列整齐,气孔通道明显,呈纺锤形。气孔与表皮细胞基本处于同一平面,大多种类气孔外缘角质层相对平坦,少数如青藏蒿(Artemisia duthreuil-de-rhinsii)、藏 沙 蒿(Artemisia wellbyi)气孔外缘呈皱状隆起。(3)从气孔器类型来看,气孔器多为无规则型,无副卫细胞,由4~6个普通表皮细胞不规则地围绕着保卫细胞;少数种类气孔器为其他类型,例如二裂委陵菜(Potentilla bifurca)为环列型,由6个副卫细胞环状排列围绕保卫细胞(图版Ⅰ,21;图版Ⅱ,20);珠芽蓼(Polygonum viviparum)为不等型,保卫细胞周围有3个副卫细胞围绕(图版Ⅰ,22;图版Ⅱ,21),绿绒蒿(Meconopsis integrifolia)为辐射型(图版Ⅰ,25;图版Ⅱ,24)。
2.2 青藏高原草地双子叶植物叶片气孔数量特征的差异性分析
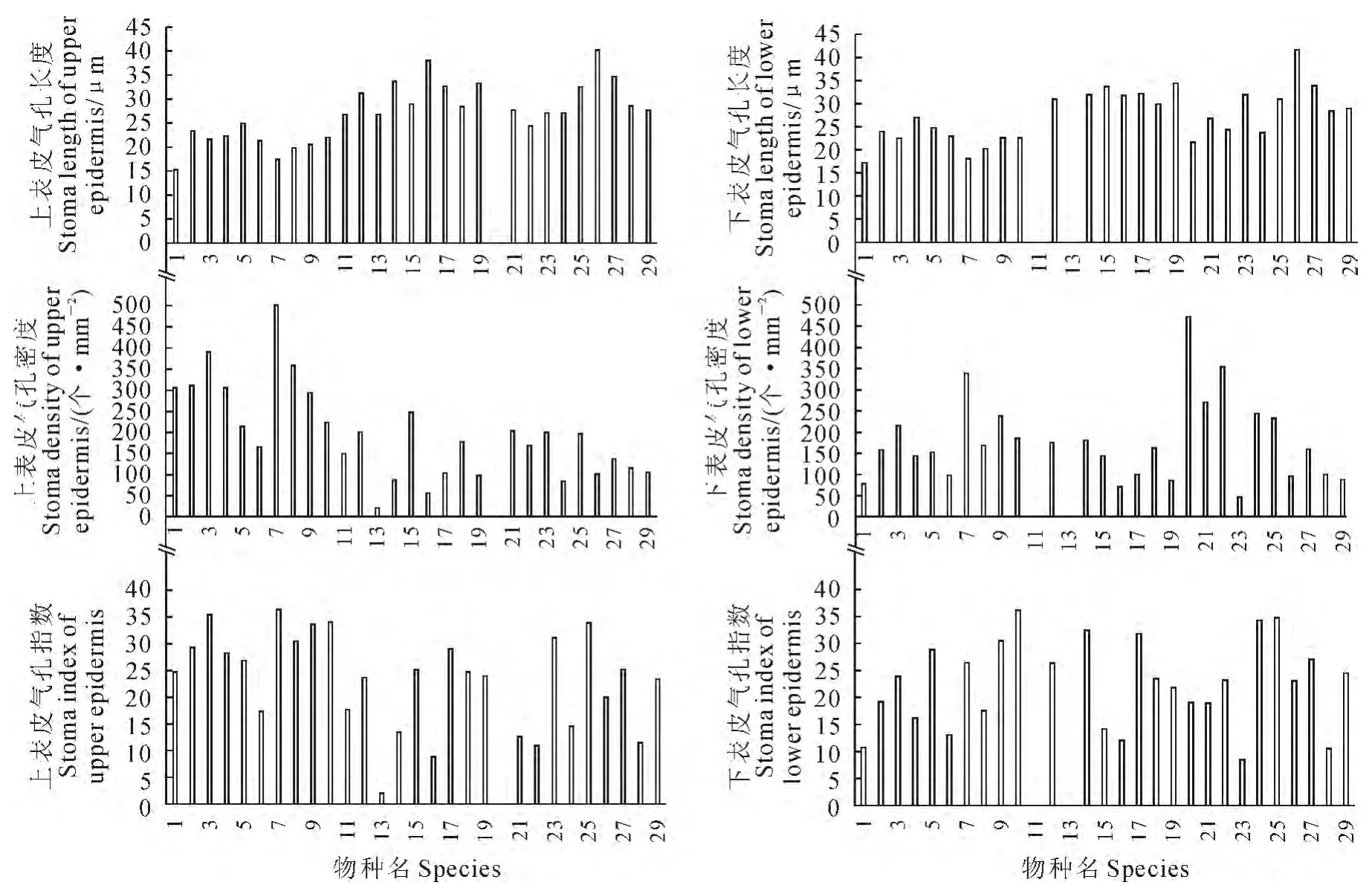
图1 青藏高原草地双子叶植物不同物种间的气孔数量特征比较1.黄花棘豆;2.兰花棘豆;3.小叶棘豆;4.毛瓣棘豆;5.急弯棘豆;6.镰形棘豆;7.二花棘豆;8.白棘豆;9.密丛棘豆;10.青海黄耆;11.银背凤毛菊;12.美丽凤毛菊;13.禾叶凤毛菊;14.雪莲;15.水母雪莲;16.凤毛菊;17.黄帚橐吾;18.青藏蒿;19.藏沙蒿;20.小叶金露梅;21.羽裂密枝委陵菜;22.二裂委陵菜;23.珠芽蓼;24.圆穗蓼;25.全缘叶马先蒿;26.绿绒蒿;27.麻花艽;28.狼毒;29.燥原芥。Fig.1 Stomatic quantative characteristics among various species of dicotyledonous plants in the Tibetan Plateau grasslands 1.Oxytropis ochrocephala;2.Oxytropis coerulea;3.Oxytropis microphylla;4.Oxytropis sericopetala;5.Oxytropis defexa;6.Oxytropis falcata;7.Oxytropis biflora;8.Oxytropis subfalcata;9.Oxytropis densa;10.Astragalus tanguticus;11.Saussurea nivea;12.Saussurea superba;13.Saussurea graminifolia;14.Saussurea globosa;15.Saussurea medusa;16.Saussurea glanduligera;17.Ligularia virgaurea;18.Artemisia duthreuil-de-rhinsii;19.Artemisia wellbyi;20.Potentilla parvifolia;21.Potentilla nivea;22.Potentilla bifurca;23.Polygonum viviparum;24.Polygonum macrophyllum;25.Pedicularis integrifolia;26.Meconopsis integrifolia;27.Gentiana straminea;28.Stellera chamaejasme;29.Ptilotrichum canescens.
2.2.1 青藏高原草地双子叶植物叶片气孔数量特征的比较 青藏高原29种草地双子叶植物叶表皮气孔数量特征的直观比较结果表明(图1):(1)从气孔长度分析:上表皮气孔长度在(15.26±2.85)μm~(40.27±3.06)μm 之间,下表皮气孔长度在(17.30±3.17)μm ~(41.85±2.68)μm 之间,气孔长度相对较小。对于上、下表皮均分布有气孔器的26种植物,其中16种(61.54%)下表皮的气孔长度大于上表皮的气孔长度,最大长度差为4.79μm。(2)从气孔密度分析:不同物种间气孔密度差异较大,如禾叶凤毛菊(Saussurea graminifolia)的气孔密度最小,其上表皮的气孔密度仅为(20.92±3.34)个/mm2,下表皮的气孔密度为0;二花棘豆(Oxytropis biflora)上表皮的气孔密度最大,为(518.03±41.05)个/mm2;小叶金露梅(Potentilla parvifolia)下表皮的气孔密度最大,为(474.95±18.60)个/mm2。(3)从气孔指数分析:不同物种叶表皮气孔指数差异明显。如上表皮气孔指数最小的为禾叶凤毛菊(Saussurea graminifolia)(8.89±2.01),最大的 为 二 花 棘 豆(Oxytropis biflora)(36.61±6.33);下表皮气孔指数最大的为青海黄耆(Astragalus tanguticus)(36.30±6.16);最小的为珠芽蓼(Polygonum viviparum)(8.38±3.56)。
对不同属间叶表皮6 项气孔数量特征进行比较,可以看出其存在明显差异(图2)。如绿绒蒿属叶片上、下表皮的气孔长度均最大,分别为(40.27±3.06)μm 与(41.85±2.68)μm,棘豆属叶片的上、下表皮气孔长度均最小,分别为(19.97±4.63)μm 与(21.50±4.46)μm;棘豆属叶片上表皮气孔密度最大为(307.30±9.98)个/mm2,委陵菜属下表皮的气孔密度最大(416.62±13.82)个/mm2;黄芪属上、下表皮气孔指数均相对较大,分别为(34.34±5.15)与(35.13±6.06)。
实验植物隶属的9个科之间叶表皮气孔数量特征也存在明显差异(图3),如罂粟科上、下表皮的气孔长度均最大,分别为(40.27±3.06)μm 与(41.85±2.68)μm;蝶形花科上表皮的气孔密度最大,蔷薇科下表皮的气孔密度最大,玄参科植物叶片上下表皮的气孔指数均较大,分别为(34.97±2.70)与(24.91±3.90)。

图2 青藏高原草地双子叶植物不同属间气孔数量特征比较1.橐吾属;2.龙胆属;3.凤毛菊属;4.蓼属;5.棘豆属;6.委陵菜属;7.绿绒蒿属;8.狼毒属;9.马先蒿属;10.黄芪属;11.蒿属;12.燥原芥属Fig.2 Stomatic quantative characteristics among different genera of dicotyledonous plants in the Tibetan Plateau grasslands 1.Ligularia;2.Gentiana;3.Saussurea;4.Polygonum;5.Oxytropis;6.Potentilla;7.Meconopsis;8.Stellera;9.Pedicularis;10.Astragalus;11.Artemisia;12.Ptilotrichum

图3 青藏高原草地双子叶植物不同科间气孔数量特征比较1.菊科;2.龙胆科;3.蓼科;4.蝶形花科;5.蔷薇科;6罂粟科;7瑞香科;8玄参科;9十字花科。Fig.3 Stomatic quantative characteristics among different families of dicotyledonous plants in the Tibetan Plateau grasslands 1.Compositae;2.Gentianaceae;3.Polygonaceae;4.Papilionaceae;5.Rosaceae;6.Papaveraceae;7.Thymelaeaceae;8.Scrophulariaceae;9.Brassicaceae.
2.2.2 青藏高原草地双子叶植物气孔数量特征的差异性统计分析 对青藏高原29种草地双子叶植物的6个气孔数量特征进行方差分析(表2),结果表明:不同物种间在上下表皮气孔长度、上下表皮气孔密度、上下表皮气孔指数总体上差异极其显著(P<0.01);不同属、不同科间叶片上下表皮气孔长度、上下表皮气孔密度、上下表皮气孔指数均差异极其显著(P<0.01)(表3、表4);从统计学上进一步验证了2.2.1中不同植物叶片气孔数量特征之间的差异性。
2.3 青藏高原草地双子叶植物气孔数量特征的相关性分析
2.3.1 气孔数量参数之间的相关性分析 对29种草地双子叶植物的上、下表皮气孔数量特征进行Pearson相关分析,结果见表5。从表中可看出上下表皮的6 个气孔数量特征之间呈显著相关(P<0.01)。对于上表皮而言,气孔长度与气孔密度、气孔指数呈显著负相关(P<0.01,r=-0.601,r=-0.160);对于下表皮而言,气孔长度与气孔密度呈显著负相关(P<0.01,r=-0.314),但与气孔指数呈显著正相关(P<0.01,r=0.158);对于上下表皮,气孔密度与气孔指数均呈显著正相关(P <0.01,r=0.640,r=0.437)。
2.3.2 气孔数量特征与植物种类的相关性分析 对青藏高原草地双子叶植物的气孔数量特征与植物种类之间进行Pearson 相关分析,结果见表6。从表中可看出叶片上表皮的气孔长度、气孔密度与不同科、属、种间显著相关(P<0.01),气孔指数与不同科、属、种间相关不显著(P>0.05);而下表皮除气孔指数与物种间相关不显著(P>0.05)外,其他指标与科、属、种间相关均极其显著(P<0.01 或0.01<P<0.05)。这在一定程度上说明,就上、下表皮相比而言,下表皮气孔特征作为植物分类依据相对比较可靠。另外,就气孔数量特征而言,气孔长度与气孔密度与不同科、属、种间统计学上差异显著,因此可作为较好的分类特征参数。

表2 青藏高原草地双子叶植物不同物种间气孔数量特征的单因素方差分析Table 2 One-way ANOVA of stomatic quantative characteristics among different species of dicotyledonous plants in the Tibetan Plateau grasslands

表3 青藏高原草地双子叶植物不同属间气孔数量特征的单因素方差分析Table 3 One-way ANOVA of stomatic quantative characteristics among different genera of dicotyledonous plants in the Tibetan Plateau grasslands

表4 青藏高原草地双子叶植物不同科间气孔数量特征的单因素方差分析Table 4 One-way ANOVA of stomatic quantative characteristics among different families of dicotyledonous plants in the Tibetan Plateau grasslands

表5 青藏高原草地双子叶植物气孔数量特征的相关性分析Table 5 The Pearson correlation analysis of stomatic quantative characteristics of dicotyledonous plants in the Tibetan Plateau grasslands

表6 青藏高原草地双子叶植物气孔特征参数与植物类群间相关性分析Table 6 The Pearson correlation analysis between stomatic quantative characteristics and groups of plants of dicotyledonous plants in the Tibetan Plateau grasslands
3 讨 论
气孔是植物表皮的特殊结构,一般由成对保卫细胞以及保卫细胞之间的孔隙组成[14],它是植物与外界环境进行CO2和H2O 等气体交换的重要通道,其对环境变化具有较强的敏感性[15]。在高山植物中,草本植物居多数[4,16],这些草本植物长期生活在气温低,昼夜温差大、气压低和太阳辐射强等不利于环境因子中,气孔的形态与数量特征对环境响应与适应,进化出了特有的气孔特征及生态适应机制。
3.1 气孔形态特征的适应
有研究表明高山植物与旱生植物类似,气孔多分布在叶 片 下 表 皮[4,16,17],有 利 于 减 少 水 分 蒸 发。而施海燕等[18]对青藏高原东缘两个不同居群的星状雪兔子(Saussurea stella)叶结构进行观察,发现上下表皮均有气孔分布。本研究对29种双子叶草地植物气孔观察结果表明,大多数(26种)叶片上下表皮均分布有气孔器,从规模尺度上验证了青藏高原双子叶草地植物为双面气孔分布类型。由于气孔不仅是水分散失的途径,同时也是气体流通的通道,因而在CO2进入和水分散失之间起平衡调节作用。青藏高原适合植物生长发育的时间只有2~4 个月[16],为了保证短时间积累足够的营养物质,求得生存与繁衍,植物在长期进化过程一定会保留有利于保持旺盛光合、呼吸代谢活力的气孔特征。青藏高原草地双子叶植物气孔的双面分布模式可以保证叶片两面同时进行光合作用,增加光合作用表面积,弥补高山环境大气中CO2的不足,促进植物与外界环境的气体交换,提高CO2的同化率,从而有效避免高原环境中高海拔、低CO2分压造成的生理胁迫。青藏高原草地双子叶植物气孔总数量的1/3到2/3(因植物种类而异)随机分散到叶片下表皮,也可在一定程度上起到限制水分蒸发的作用,有利于植物较好保持水分,避免水分过度蒸发而造成生理干旱。这种“位置改变”策略在保证植物获得最大限度CO2同化量的同时,阻止植株水分降低到使组织受害的水平,是高山草地双子叶植物对高寒极端环境长期适应的结果。另外,青藏高原草地双子叶植物气孔器多呈无规则型,没有副卫细胞限制,虽然缺少了副卫细胞对保卫细胞的协同作用,但这样气孔运动时阻力较小,气孔开闭更灵活,同时能量耗散也减少,更有利于光能的有效利用,这对植物在高寒极端环境中降低蒸腾、增强储水以及提高光合效率是有利的。
3.2 气孔数量特征的适应
植物气孔长度、气孔密度和气孔指数等参数的大小均能反映植物的生物学和生态学特性。青藏高原草地双子叶植物气孔数量特征统计结果表明,大多数物种叶片的气孔密度显著高于已报道的拟南芥(Arabidopsis thaliana)叶片[19],反映出叶表皮气孔数量上的相对优势。Woodward[20]对比200年前的草本植物与现在的植物,发现气孔密度和气孔指数与CO2浓度呈负相关关系。青藏高原海拔高、空气中CO2稀薄,植物叶片气孔密度高,从侧面验证了这一点。高的气孔密度有利于增强单位面积叶片的气体交换能力,这样当部分气孔生理活动受到抑制时,其余功能正常的气孔能做出适度调节,在功能上及时补偿,使叶片的基本生理过程得以维持。贺金生等研究表明,气孔数目与密度的增加,可提高叶片与外界环境的气体交换能力,增强叶片对CO2的摄入,以提高光合作用速率,是植物对高山环境中低CO2和O2分压的适应性[21]。单位面积具有较多数量的气孔,可以保证在低CO2和O2分压条件下,以有限的水分损耗,换取尽可能大的CO2同化量,维持光合作用正常进行,保证植物完成生命周期。对于一些植物种类如禾叶凤毛菊与银背凤毛菊,仅上表皮具有气孔,且气孔密度较低,仅为同属植物美丽凤毛菊上表皮气孔密度的10%与70%,它们可能通过气孔开度增加、气孔导度增大、CO2补偿点降低,或者形成“无光合午休”等机制提高CO2固定效率,对叶表皮气孔密度的减小进行生态补偿,从而保证植物在低温和低CO2分压下顺利进行光合作用。但高的气孔密度在增加CO2吸收的同时,必然会增强水分蒸发,而这在高海拔、低温、大风环境中更易造成植物的“生理干旱”。青藏高原草地双子叶植物上表皮气孔的平均长度为26.20μm,下表皮气孔的平均长度为25.56μm,气孔长径相对较小。小的气孔除了可减少水分流失外,还具有灵活的调节功能,有利于快速关闭,可在一定程度上减少水分散失、降低污染物的侵入机会,对高气孔密度可能造成的“生理干旱”通过进行代偿,这可能是植物对高原寒冷强辐射环境作出的响应和适应性进化。
3.3 气孔特征作为分类依据
气孔是植物在自然选择中保留下来的重要器官,具有较大的稳定性,可以用来研究植物的亲缘关系、演化过程以及地理起源。Barthlott[22]指出气孔指数及气孔大小都具有分类学意义。姚兆华等[23]认为气孔大小、气孔密度和气孔指数均可以作为分类依据,尤其是气孔大小种间差异明显,分类作用更为明显。就青藏高原草地双子叶植物而言,仅就气孔的形态与气孔器类型来看,气孔多为椭圆形或宽椭圆形,气孔复体多为无规则型,不同植物种类之间区分度很弱,分类价值较小。但从气孔数量特性分析,则对植物分类具有较重要的参考价值。实验研究的青藏高原优势代表物种,植物叶片上、下表皮气孔长度、气孔密度与气孔指数与植物类群之间普遍存在明显相关关系,且在不同科、属以及种间差异明显,尤其是气孔长度与气孔密度指标在不同类群之间的差异比较稳定,虽然在一定程度上承载了高寒地带植物系统演化中气候环境的修饰痕迹,但仍然保持种内植物叶片独特的特征,体现了物种遗传上的相对稳定性,因此可以作为青藏高原双子叶草地植物分类的辅助依据。
[1] YANG Y H(杨元合),PIAO SH L(朴世龙).Variations in grassland vegetation cover in relation to climatic factors on the Tibetan Plateau[J].Chinese Journal of Plant Ecology(植物生态学报),2006,30(1):1-8.
[2] LI W H(李文华),ZHOU X M(周兴民).Ecosystems of Qinghai-Xizang(Tibetan)Plateau and Approaches for Their Sustainable Managmen[M].Guangzhou:Guangdong Science and Technology Press(广东科技出版社),1998:183-270.
[3] LU C F(卢存福),JIAN L CH(简令成),KUANG T Y(匡廷云).Freezing hardiness in alpine plants[J].Chinese Bulletin of Botany(植物学报),1998,15(3):17-22(in Chinese).
[4] ZHOU G T(周广泰),LIU F Q(刘凤琴),GUO SH X(郭书贤),et al.A study of characteristics of the anatomical structure of alpine plants[J].Journal of Qinghai Normal University(Natural Science Edition)(青海师范大学学报·自然科学版),1992,(4):45-60(in Chinese).
[5] BOSABALIDIS A M,KOFIDIS G.Comparative effects of drought stress on leaf anatomy of two olive cultivars[J].Plant Science,2002,163(2):375-379.
[6] HU J Y胡建莹GUO K郭 柯DONG M董 鸣.Variation of leaf structure of two dominant species in alpine grassland and the relationship between leaf structure and ecological factors[J].Chinese Journal of Plant Ecology(植物生态学报),2008,32(2):370-378(in Chinese).
[7] HE J S,WANG Z H,WANG X P,et al.A test of the generality of leaf trait relationships on the Tibetan Plateau[J].New Phytologist,2006,170(4):835-848.
[8] LI F L(李芳兰),BAO W K(包维楷).Responses of the morphological and anatomical structure of the plant leaf to environmental change[J].Chinese Bulletin of Botany(植物学通报),2005,22(S):118-127(in Chinese).
[9] MOTT K A,GIBSON A C,OLEARY J W.The adaptive significance of amphistomatic leaves[J].Plant Cell and Environment,1982,5(6):445-460.
[10] LI Q F(李全发),WANG B J(王宝娟),AN L H(安丽华),et al.Leaf anatomical characteristics of the plants of grasslands in the Tibetan Plateau[J].Acta Ecologica Sinica(生态学报),2013,33(7):2 062-2 070(in Chinese).
[11] MA J J(马建静),JI CH J(吉成均),HAN M(韩 梅),et al.Comparative analyses of leaf anatomy of di-cotyledonous species in Tibetan and Inner Mongolian grasslands[J].Scientia Sinica Vitae(中国科学·生命科学),2012,42(2):158-172(in Chinese).
[12] ROCAS G,SCARANO F R,BARROS C F.Leaf anatomical variation in Alchornea triplinervia(Spreng)Mull.Arg.(Euphorbiaceae)under distinct light and soil water regimes[J].Botanical Journal of the Linnean Society,2001,136(2):231-238.
[13] MENDES M M,GAZARINI L C,RODRIGUES M L.Acclimation of Myrtus communis to contrasting Mediterranean light environments effects on structure and chemical composition of foliage and water relations[J].Environmental and Experimental Botany,2001,45(2):165-178.
[14] ZHENG Y L(郑玉龙),JIANG CH L(姜春玲),FENG Y L(冯玉龙).Stomatal generation in plants[J].Plant Physiology Journal(植物生理学通讯),2005,41(6):847-850(in Chinese).
[15] HETHERINGTON A M,WOODWARD F I.The role of stomata in sensing an driving environmental change[J].Nature,2003,424(6 951):901-908.
[16] WANG W Y(王为义).An investigation on specific structural characteristics of alpine plants on Qinghai-Xizang Plateau[J].Acta Biologica Plateau Sinica(高原生物学集刊),1985,(4):19-34(in Chinese).
[17] WU X M(吴学明).A study on anatomical characteristics of leaves and stems of five alpine plants used for Tibetan medicine in Qinghai-Tibetan Plateau[J].Acta Botanica Boreali-Occidentalia Sinica(西北植物学报),1996,16(1):56-60(in Chinese).
[18] SHI H Y(施海燕),WANG Y F(王一峰),GAO H Y(高宏岩).Adaptation of anatomical structures of the leaves of Saussurea stella Maxim to the alpine environment[J].Guihaia(广西植物),2010,30(5):621-625(in Chinese).
[19] INAN G,ZHANG Q,LI P H,et al,Saltcress:A halophyte and cryophyte arabidopsis relative model system and its applicability to molecular genetic analyses of growth and development of extremophiles[J].Plant Physiology,2004,135(3):1 718-1 737.
[20] WOODWARD F I.Stomatal numbers are sensitive to increases in CO2from pre-industrial levels[J].Nature,1987,327(6 123):617-618.
[21] HE J SH(贺金生),CHEN W L(陈伟烈),WANG X L(王勋陵).Morphological and anatomical features of QuercusSection Suber and its adaptation to the ecological environment[J].Chinese Journal of Plant Ecology(植物生态学报),1994,18(3):219-227(in Chinese).
[22] BARTHLOTT W.The Taxonomic Importance of the Leaf Surface[M]//HEYWOOD V H,MOORE D M.Current Concept in Plant Taxonomy.London:Academic Press,1984:67-94.
[23] YAO ZH H(姚兆华),HAO L ZH(郝丽珍),WANG P(王 萍),et al.Study on the stomata characters of leaves of the Puionium Gaertn[J].Bulletin of Botanical Research(植物研究),2007,27(2):199-203.

图版Ⅰ 青藏高原草地双子叶植物叶片上表皮的气孔特征1.黄花棘豆;2.兰花棘豆;3.小叶棘豆;4.毛瓣棘豆;5.急弯棘豆;6.镰形棘豆;7.二花棘豆;8.白棘豆;9.密丛棘豆;10.青海黄耆;11.银背凤毛菊;12.美丽凤毛菊;13.禾叶凤毛菊;14.雪莲;15.水母雪莲;16.凤毛菊;17.黄帚橐吾;18.青藏蒿;19.藏沙蒿;20.羽裂密枝委陵菜;21.二裂委陵菜;22.珠芽蓼;23.圆穗蓼;24.全缘叶马先蒿;25.绿绒蒿;26.麻花艽;27.狼毒;28.燥原芥。PlateⅠ Stomatic characteristics of upper epidermis of dicotyledonous plants in the Tibetan Plateau grasslands Fig.1.Oxytropis ochrocephala;Fig.2.Oxytropis coerulea;Fig.3.Oxytropis microphylla;Fig.4.Oxytropis sericopetala;Fig.5.Oxytropis defexa;Fig.6.Oxytropis falcata;Fig.7.Oxytropis biflora;Fig.8.Oxytropis subfalcata;Fig.9.Oxytropis densa;Fig.10.Astragalus tanguticus;Fig.11.Saussurea nivea;Fig.12.Saussurea superba;Fig.13.Saussurea graminifolia;Fig.14.Saussurea globosa;Fig.15.Saussurea medusa;Fig.16.Saussurea glanduligera;Fig.17.Ligularia virgaurea;Fig.18.Artemisia duthreuil-de-rhinsii;Fig.19.Artemisia wellbyi;Fig.20.Potentilla nivia;Fig.21.Potentilla bifurca;Fig.22.Polygonum viviparum;Fig.23.Polygonum macrophyllum;Fig.24.Pedicularis integrifolia;Fig.25.Meconopsis integrifolia;Fig.26.Gentiana straminea;Fig.27.Stellera chamaejasme;Fig.28.Ptilotrichum canescens.

图版Ⅱ 青藏高原草地双子叶植物叶片的下表皮气孔特征1.黄花棘豆;2.兰花棘豆;3.小叶棘豆;4.毛瓣棘豆;5.急弯棘豆;6.镰形棘豆;7.二花棘豆;8.白棘豆;9.密丛棘豆;10.青海黄耆;11.美丽凤毛菊;12.雪莲;13.水母雪莲;14.凤毛菊;15.黄帚橐吾;16.青藏蒿;17.藏沙蒿;18.小叶金露梅;19.羽裂密枝委陵菜;20.二裂委陵菜;21.珠芽蓼;22.圆穗蓼;23.全缘叶马先蒿;24.绿绒蒿;25.麻花艽;26.狼毒;27.燥原芥;28.黄花棘豆(表皮毛)。PlateⅡ Stomatic characteristics of lower epidermis of dicotyledonous plants in the Tibetan Plateau grasslandsFig.1.Oxytropis ochrocephala;Fig.2.Oxytropis coerulea;Fig.3.Oxytropis microphylla;Fig.4.Oxytropis sericopetala;Fig.5.Oxytropis defexa;Fig.6.Oxytropis falcata;Fig.7.Oxytropis biflora;Fig.8.Oxytropis subfalcata;Fig.9.Oxytropis densa;Fig.10.Astragalus tanguticus;Fig.11.Saussurea superba;Fig.12.Saussurea globosa;Fig.13.Saussurea medusa;Fig.14.Saussurea glanduligera;Fig.15.Ligularia virgaurea;Fig.16.Artemisia duthreuil-de-rhinsii;Fig.17.Artemisia wellbyi;Fig.18.Potentilla parvifolia;Fig.19.Potentilla nivia;Fig.20.Potentilla bifurca;Fig.21.Polygonum viviparum;Fig.22.Polygonum macrophyllum;Fig.23.Pedicularis integrifolia;Fig.24.Meconopsis integrifolia;Fig.25.Gentiana straminea;Fig.26.Stellera chamaejasme;Fig.27.Ptilotrichum canescens;Fig.28.Oxytropis ochrocephala(epidermal hair).
猜你喜欢
今日农业(2022年3期)2022-11-16 13:13:50
干旱地区农业研究(2022年1期)2022-01-28 04:51:30
装备制造技术(2019年12期)2019-12-25 03:06:52
甘肃林业(2019年4期)2019-10-08 07:19:52
科技创新与品牌(2019年12期)2019-02-06 04:03:29
中成药(2018年3期)2018-05-07 13:34:31
中国铸造装备与技术(2015年5期)2015-12-10 10:23:33
天然产物研究与开发(2014年3期)2014-04-27 14:15:34
生物进化(2014年2期)2014-04-16 04:36:27
中医研究(2013年3期)2013-03-11 20:26:39
