
高丹草叶片PSⅡ光化学活性的抗旱优势
2015-06-24 13:09胡举伟张会慧张秀丽逄好胜孙广玉
草业科学 2015年3期
胡举伟,张会慧, 2,张秀丽,逄好胜,孙广玉
(1.东北林业大学生命科学学院, 黑龙江 哈尔滨 150040;2.黑龙江省科学院自然与生态研究所,黑龙江 哈尔滨 150040)
高丹草叶片PSⅡ光化学活性的抗旱优势
胡举伟1,张会慧1, 2,张秀丽1,逄好胜1,孙广玉1
(1.东北林业大学生命科学学院, 黑龙江 哈尔滨 150040;2.黑龙江省科学院自然与生态研究所,黑龙江 哈尔滨 150040)
本试验利用快速叶绿素荧光诱导动力学分析技术研究了高丹草(Sorghumbicolor×S.sudanense)和苏丹草(S.sudanense)叶片PSⅡ光化学活性对干旱的响应。结果表明,随着干旱的加剧,高丹草和苏丹草幼苗叶片PSⅡ的最大光化学效率(Fv/Fm)和效能指数(PIABS)明显降低,特别是PIABS的降低幅度明显大于Fv/Fm,即干旱下两种牧草幼苗叶片的PSⅡ光化学活性降低,但不同干旱程度下高丹草幼苗叶片的PSⅡ光化学活性明显高于苏丹草。分析原因可以发现:干旱下高丹草幼苗叶片OJIP曲线上0.15和0.3 ms时(L点和K点)的相对可变荧光VL和VK增加幅度明显小于苏丹草,即干旱条件下对高丹草幼苗叶片PSⅡ电子供体侧放氧复合体OCE活性和类囊体聚合状态的影响明显小于苏丹草,这对于电子传递链上的电子供应及电子传递的稳定性起到了重要的作用。另外,干旱下两种牧草幼苗叶片OJIP曲线上2和30 ms时(J点和I点)的相对可变荧光VJ和VI明显增加,即干旱导致PSⅡ光化学活性降低还与PSⅡ电子受体侧电子由QA向QB传递受阻以及PQ库接受电子能力的降低有关。但干旱下高丹草和苏丹草幼苗叶片仅VJ差异明显,而VI却无明显差异,这说明干旱下高丹草幼苗叶片电子传递体QB接受电子的能力明显高于苏丹草,从而使干旱下高丹草幼苗叶片PSⅡ受体侧电子传递能力高于苏丹草。
高丹草;杂交种;干旱;PSⅡ
杂种优势是生物界普遍存在的生物学现象,特别是在逆境条件下,如玉米(Zeamays)[1]、水稻(Oryzasativa)[2]、大豆(Glycinemax)[3]和红麻(Hibiscuscannabinus)[4]等杂交种均具有明显高于亲本的抗逆优势。高丹草(Sorghumbicolor×S.sudanense)为高粱(S.bicolor)与苏丹草(S.sudanense)的杂交种,不但具有苏丹草再生性强、营养价值高、适口性好等优点,而且具有高粱抗旱、抗倒伏的抗逆优势[5-6],每年可刈割2~3次,产草量高,在一定的干旱等胁迫条件下能维持正常生长[7],并且在广大农牧区已被较为广泛地种植[8-9],是干旱、贫瘠和盐碱等困难立地畜牧业生产中的重要牧草品种。植物体中90%~95%的干物质为光合作用的产物[10],由于植物光合作用的光反应阶段、暗反应阶段都会受到干旱影响,使植物的光化学活性降低,碳同化能力下降,最终表现为植物的生长受到影响[11],所以使植物的光合能力保持稳定,是植物适应干旱这种胁迫条件的决定因素[10-11]。与多数具有灌溉条件的农田相比,畜牧业地区缺少灌溉条件和设施,加之地域性气候因素的限制,干旱已经严重限制了我国干旱、半干旱地区的畜牧业发展[12]。因此,选育推广耐旱的牧草新品种在发展干旱、半干旱地区畜牧业生产中尤为重要。
大量的试验表明,杂交能明显提高植物的光合能力,特别是在逆境条件下,杂交种的光合能力明显高于亲本[13-14],高丹草的抗旱优势在农艺性状[15]和产量[16]等方面已经得到了证实,干旱胁迫下杂交高丹草的生长和光合机构功能明显优于亲本苏丹草,干旱胁迫下杂交高丹草叶片的电子传递速率也明显高于苏丹草[17]。为揭示干旱下杂交高丹草具有较高光合能力的内在原因,本试验利用快速叶绿素荧光动力学分析技术研究干旱下杂交高丹草与苏丹草叶片PSⅡ电子供体侧放氧复合体OEC的活性,电子传递受体侧QA向QB的传递以及PQ库容量和类囊体膜状态等方面的差异,以期在光化学活性方面揭示高丹草抗旱优势的生理基础,为丰富杂交高丹草的抗旱优势的机理研究,提供一定的数据支撑。
1 材料与方法
1.1 试验材料
试验以苏丹草、高丹草两种牧草为材料,于2011年3月在东北林业大学植物生理学实验室进行。将高丹草和苏丹草的种子播种到直径12 cm、高15 cm的营养钵中,每钵15粒种子,并使其均匀分布在营养钵中,播种后覆土约2 cm。营养钵底部有渗水口,混合的草炭土和蛭石比例1∶1(v/v)为培养基质。将营养钵放入光照培养箱,光照时温度为23~27 ℃,黑暗时温度为17~22 ℃,每24 h中照光12 h,黑暗12 h,相对湿度75%左右,设定光强为200 μmol·m-2·s-1,适时补充水分,并清除杂草,当两种牧草植株株高约10 cm时,每个营养钵保留8株幼苗(植株大小近似、株距合理),当两种牧草株高约40 cm时,停止浇水,开始自然干旱处理。分别于干旱处理1、4、7、10 d时进行高丹草和苏丹草幼苗叶片OJIP曲线的测定。
1.2 定参数和方法
OJIP曲线的测定:每个处理挑选植株大小相近的5株幼苗,每株选取苏丹草和高丹草幼苗从顶端数第3片完全展开叶片,暗适应0.5 h后,采用Mini调制式掌上叶绿素荧光仪(FluorPen FP 100 max,捷克)对苏丹草和高丹草叶片的OJIP曲线进行测定,共计5次重复,测定时荧光仪的照光位置避开叶片的主叶脉,并且选取叶基和叶尖的中间位置。利用5次重复的平均值绘制OJIP曲线,其中OJIP曲线上的O、J、I和P点分别为0、2、30和1 000 ms对应的时刻,相对荧光强度分别以Fo、FJ、FI和Fm表示,为分别分析L、K、J和I点(0.15、0.3、2和30 ms的对应时刻)的相对可变荧光VL、VK、VJ和VI变化,按照Zhang等[3]的方法分别进行O-K、O-J和O-P曲线标准化,即分别得到相对可变荧光VO-K、VO-J和VO-P的时间对应曲线[17],其计算公式分别为:VO-K=(Ft-Fo)/(FK-Fo)、VO-J=(Ft-Fo)/(FJ-Fo)、VO-P=(Ft-Fo)/(Fm-Fo),式中Ft表示各时间点的相对荧光强度,FK为K点即0.3 ms时的相对荧光强度。对测得的OJIP曲线进行JIP-test分析,分别得到PSⅡ的最大光化学效率(Fv/Fm)、效能指数(PIABS)。
1.3 数据处理方法
运用Excel 2007和SPSS 17.0软件对测定数据进行统计分析,采用单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)比较不同数据组间的差异,图中数据为平均值±标准差(SD)。
2 结果与分析
2.1 干旱对苏丹草和高丹草幼苗叶片OJIP曲线形态的影响
不同干旱程度下苏丹草和高丹草幼苗叶片的OJIP曲线形态均发生了明显的变化,随着干旱天数的增加,O点的相对荧光强度(Fo)呈增加趋势,而P点的相对荧光强度(Fm)降低(图1)。但是不同干旱天数时高丹草各点的相对荧光强度变化幅度明显小于苏丹草,定量分析结果表明,干旱1 d时两种牧草叶片的Fo和Fm均无显著差异,但干旱10 d时,高丹草幼苗叶片的Fo低于苏丹草16.22%(P<0.05),而Fm高于苏丹草25.44%(P<0.05),均达显著差异水平。
2.2 干旱对苏丹草和高丹草幼苗叶片光系统Ⅱ光化学活性的影响
最大光化学效率(Fv/Fm)、效能指数(PIABS)可以表征光系统Ⅱ的光化学活性。随着干旱时间的延长,苏丹草和高丹草植株叶片的Fv/Fm、PIABS呈现明显降低的趋势,与Fv/Fm相比,PIABS降低幅度更大(图2)。除干旱1 d时苏丹草和高丹草幼苗叶片的Fv/Fm之间无显著差异外,干旱4~10 d时高丹草幼苗叶片的Fv/Fm均显著高于苏丹草。干旱1 d和4 d时高丹草幼苗叶片的PIABS分别高于苏丹草52.46%(P<0.05)和113.69%(P<0.05),但干旱7 d和10 d时高丹草幼苗叶片的PIABS与苏丹草却未达显著差异水平(P>0.05)。
2.3 干旱对苏丹草和高丹草幼苗叶片OJIP曲线上J点和I点相对可变荧光的影响
分别将不同干旱天数下苏丹草和高丹草幼苗叶片的OJIP曲线按O-P标准化后可以看出(图3),随着干旱时间的延长,苏丹草和高丹草幼苗J点(2 ms)和I点(30 ms)的相对可变荧光VJ和VI均呈明显的增加趋势,并且VJ的变化幅度大于VI,且高丹草VJ和VI的变化幅度明显小于苏丹草。除干旱7 d外,高丹草幼苗叶片的VJ均显著低于苏丹草,特别是在干旱10 d时,高丹草幼苗叶片的VJ低于苏丹草20.01%(P<0.05),而苏丹草和高丹草幼苗叶片的VI在不同干旱程度下差异均不显著(P>0.05)。
2.4 干旱对苏丹草和高丹草幼苗叶片OJIP曲线上K点相对可变荧光的影响
分别将干旱下苏丹草和高丹草幼苗叶片的OJIP曲线按O-J标准化后可以看出,不同干旱天数下苏丹草和高丹草叶片标准化O-J曲线在0.3 ms时,干旱10 d的相对可变荧光与干旱1 d相比增加最为明显,即K点的相对可变荧光VK随着干旱的加剧而增加,但不同干旱天数下高丹草叶片的VK增加幅度明显低于苏丹草(图4)。干旱1 d时苏丹草和高丹草幼苗叶片的VK差异不显著(图5),而干旱4、7和10 d时,高丹草幼苗叶片的VK分别低于苏丹草11.12%(P<0.05)、7.17%(P>0.05)和15.45%(P<0.05)。

图1 干旱对苏丹草和高丹草叶片OJIP曲线形态的影响Fig.1 OJIP tramsient in leaves of Sorghum sudanense and S. bicolor×S. sudanense seedlings under drought stress
注:图表中数据为平均值±标准差;不同小写字母表示不同处理之间差异显著(P<0.05)。下同。
Note: Data in the figures are present as mean±SD; Different lower case letters mean significant difference (P<0.05). The same below.

图2 干旱对苏丹草和高丹草叶片Fv/Fm 和PIABS的影响Fig.2 Fv/Fm and PIABS in leaves of Sorghum sudanense and S. bicolor×S. sudanense seedlings under drought stress
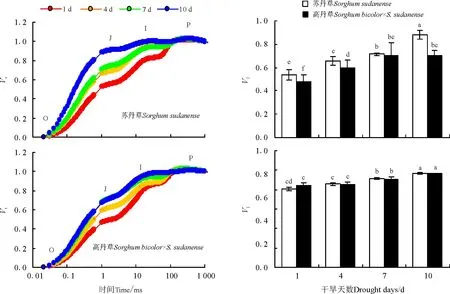
图3 干旱胁迫对苏丹草和高丹草叶片OJIP曲线上J点和I点相对可变荧光的影响Fig.3 VJ and VI in leaves of Sorghum sudanense and S. bicolor×S. sudanense seedlings under drought stresss

图4 干旱对苏丹草和高丹草叶片OJIP曲线上K点和L点相对可变荧光的影响Fig.4 VK and VL in leaves of Sorghum sudanense and S. bicolor×S. sudanense seedlings under drought stress

图5 干旱对苏丹草和高丹草叶片OJIP曲线上K点和L点相对可变荧光的影响Fig.5 VK and VL in leaves of Sorghum sudanense and S. bicolor×S. sudanense seedlings under drought stress
2.5 干旱对苏丹草和高丹草幼苗叶片OJIP曲线上L点相对可变荧光的影响
进一步将干旱条件下苏丹草和高丹草幼苗叶片的OJIP曲线按O-K标准化后可以看出(图4),苏丹草和高丹草幼苗标准化O-K曲线上0.15 ms时(L点)的相对可变荧光VL随着干旱程度的加剧明显增加,不同干旱程度下高丹草幼苗叶片的VL增加幅度明显小于苏丹草。虽然在干旱1 d时高丹草幼苗叶片的VL稍高于苏丹草,但干旱4、7和10 d时,高丹草幼苗叶片的VL分别低于苏丹草7.55%(P<0.05)、4.63%(P>0.05)和10.17%(P<0.05)。
3 讨论与结论
土壤干旱下植物根系吸水困难,导致叶片含水量降低而使光合机构遭到破坏,而较多的研究表明,逆境条件下光合机构的损伤部位多发生在PSⅡ,因此,保证相对较高的PSⅡ光化学活性是反映植物具有较强抗旱性的关键。快速叶绿素荧光诱导动力学曲线(OJIP)分析JIP-test方法的应用对于研究逆境下植物光合作用,特别是原初反应中的损伤部位提供了便捷的工具[18]。本试验利用快速叶绿素荧光诱导动力学技术研究了杂交高丹草和苏丹草幼苗叶片PSⅡ功能对干旱的响应,结果发现,干旱明显改变了高丹草和苏丹草幼苗叶片的OJIP曲线形态,其中Fo随着干旱时间的延长而增加,而Fm则明显降低,即干旱下高丹草和苏丹草幼苗叶片的OJIP曲线变得相对平缓,但不同干旱天数下高丹草幼苗叶片OJIP曲线变化幅度均明显小于苏丹草。另外,干旱还导致高丹草和苏丹草幼苗叶片的Fv/Fm和PIABS的明显降低,Fv/Fm和PIABS均是反映植物PSⅡ光化学活性的重要指标,特别是PIABS敏感性明显大于Fv/Fm[19],因此干旱胁迫导致了高丹草和苏丹草幼苗叶片PSⅡ反应中心光化学活性的降低,甚至导致光抑制的发生。但本试验中不同干旱程度下高丹草幼苗叶片的Fv/Fm和PIABS均明显高于苏丹草,说明干旱条件下高丹草幼苗叶片可以保证相对较高的PSⅡ光化学活性,以保证植株叶片光系统Ⅱ原初光化学反应维持正常。
植物在胁迫环境中光系统Ⅱ反应中心光化学活性下降主要体现为PSⅡ电子传递速率的降低,而有研究认为,电子传递速率的降低主要与PSⅡ电子供体侧电子供应能力的降低和PSⅡ电子受体侧电子传递的受阻有关[20]。为分析高丹草在干旱下具有较高光化学活性的原因,本试验分析了干旱对高丹草和苏丹草幼苗叶片PSⅡ电子供体侧和电子受体侧电子产生和传递的影响,结果发现,高丹草和苏丹草幼苗叶片O-J标准化曲线上K点的相对可变荧光VK随着干旱的天数的增加而增加,VK的增加是特异反映叶绿体内PSⅡ电子供体侧放氧复合体OEC受损的标志[18],说明干旱导致高丹草和苏丹草幼苗叶片OEC活性的降低,使光合原初反应中水的裂解受到抑制,进而导致其光合电子传递链上的电子供应能力受限。但不同干旱程度下高丹草幼苗叶片的VK均明显低于苏丹草,说明高丹草幼苗叶片OEC活性对干旱的耐受性高于苏丹草,这就保证了干旱下高丹草幼苗叶片光合电子传递链上的电子供应。OJIP曲线上J点荧光变化反映了PSⅡ电子受体侧QA向QB的电子传递特征,而I点荧光变化则反映了PQ库的异质性[21],本试验中干旱导致了高丹草和苏丹草幼苗叶片J点和I点的相对可变荧光VJ和VI明显增加,说明干旱导致PSⅡ电子传递受阻的原因与PSⅡ受体侧QA向QB传递受阻以及PQ库接受电子能力降低有关。高丹草和苏丹草相比,干旱过程中高丹草幼苗叶片的VJ明显低于苏丹草,但二者叶片的VI在不同干旱程度下却均没有显著差异,说明干旱下高丹草幼苗电子传递速率高于苏丹草的原因主要是高丹草PSⅡ电子受体侧电子由QA向QB传递过程没有受到明显抑制。
逆境下植物叶片电子供体侧OEC活性的降低会导致放氧过程中水裂解功能异常和叶绿体内H2O2的产生[22],另外,电子传递受体侧电子传递受阻时过剩电子也会与叶绿体内游离的O2结合产生超氧阴离子[23],叶绿体内超氧阴离子和H2O2等活性氧的爆发会使叶绿体膜质过氧化加剧,导致叶绿体内的类囊体膜的破坏甚至类囊体的解离和PSⅡ结构的破坏[24],而标准化O-K曲线上L点相对可变荧光VL的上升则是类囊体解离的特异性标志[25],本试验中干旱下高丹草和苏丹草幼苗叶片的VL均呈明显的增加趋势,结合图3、图4和图5的结果,说明干旱可能导致高丹草和苏丹草幼苗类囊体解离的原因与电子传递链上电子传递受阻有关。但是干旱4~10 d高丹草幼苗叶片的VL增加幅度均明显小于苏丹草,说明干旱下高丹草幼苗叶片PSⅡ结构在干旱下具有较苏丹草更强的稳定性,这在维持其PSⅡ的光化学活性,减轻其光抑制程度方面发挥了重要作用。
综上所述,干旱胁迫下高丹草和苏丹草幼苗叶片PSⅡ光化学活性降低的原因与干旱导致PSⅡ电子供体侧放氧复合体OEC活性的降低以及PSⅡ电子受体侧电子传递受阻以及光合机构的破坏有关。而不同干旱程度下高丹草幼苗叶片OEC的活性以及电子QA向QB传递速度均明显高于苏丹草,但QB向PQ传递过程二者之间却无明显差异,即高丹草幼苗叶片QB接受电子的能力明显高于苏丹草。较高的OEC的活性以及电子传递速率有效减轻了高丹草幼苗叶片的类囊体的解离程度,这对维持其PSⅡ结构和功能发挥了重要的作用。
[1] 袁力行,傅骏骅,刘新芝,彭泽斌,张世煌,李新海,李连城.利用分子标记预测玉米杂种优势的研究[J].中国农业科学,2000,33(6):6-12.
[2] 曾千春,周开达,朱祯,罗琼.中国水稻杂种优势利用现状[J].中国水稻科学,2000,14(4):243-246.
[3] Zhang X K,Zhou Q H,Cao J H,Yu B J.Differential Cl-/salt tolerance and NaCl-induced alternations of tissue and cellular ion fluxes inGlycinemax,Glycinesojaand their hybrid seedlings[J].Journal of Agronomy and Crop Science,2011,197(5):1-11.
[4] 张加强,潘凤英,窦俊焕,陈鹏,周琼,周瑞阳.红麻杂交种的根系生长对盐胁迫的抗性优势表现[J].中国农业大学学报,2011,16(5):6-12.
[5] 张晓艳,刘锋,王风云,张吉旺,叶梅,董树亭.施氮对杂交苏丹草植株硝态氮累积及产量的影响[J].草地学报,2009,17(3):327-332.
[6] 于卓,刘永伟,赵晓杰,马艳红,李小雷,赵娜.高粱11A与3种苏丹草杂交种F1的农艺特性及细胞遗传学研究[J].草业学报,2006,15(1):90-96.
[7] 赵娜,于卓,马艳红,李小雷,周亚星.高丹草幼苗抗旱和耐盐性的品种间差异[J].中国草地学报,2007,29(3):39-44.
[8] Chung J M,Lee B C,Kim J S.Fine-scale structure among genetic individuals of clone-forming monotypic genusEchinosophorakoreensis(Fabaceae)[J].Annals of Botany,2006,98(1):165-173.
[9] 詹秋文,林平.高粱-苏丹草杂交种研究与利用前景[J].草业学报,2001,10(2):56-61.
[10] Kramer D M,Johnson G,Kiirats O,Edwards G E.New fluorescence parameters for the determination of QAredox state and excitation energy fluxes [J].Photosynthesis Research,2004,79(2):209-218.
[11] Hendrickson L,Förster B,Pogson B J.A simple chlorophyll fluorescence parameter that correlates with the rate coefficient of photoinactivation of photosystemⅡ[J].Photosynthesis Research,2005,84(1-3):43-49.
[12] 赵山志,夏明,吴新宏,侯武英,刘雅学,阿拉木斯,武斌.干旱、半干旱草原退化、沙化区的重要选择——设施草业生产[J].农业工程学报,2003,19(1):184-186.
[13] 李霞,丁在松,李连禄,王美云,赵明.玉米光合性能的杂种优势[J].应用生态学报,2007,18(5):1049-1054.
[14] 王荣富,张云华,钱立生,于江龙.超级杂交稻两优培九及其亲本的光氧化特性[J].应用生态学报,2003,14(8):1309-1312.
[15] 于卓,山田敏彦.高丹草品种主要农艺性状的比较研究[J].中国草地学报,2006,28(6):1-6.
[16] 李源,谢楠,赵海明,刘贵波.不同高丹草品种对干旱胁迫的响应及抗旱性评价[J].草地学报,2010,18(6):891-896.
[17] Zhang L T,Gao H Y,Zhang Z S,Xue Z C,Meng Q W.Multiple effects of inhibition of mitochondrial alternative oxidase pathway on photosynthetic apparatus in Rumex K-1 leaves[J].Biologia Plantarum,2012,56(2):365-368.
[18] Strasser R J,Srivastava A,Govindjee.Polyphasic chlorophyll a fluorescence transient in plants and cyanobacteria[J].Photochemistry and Photobiology,1995,61(1):32-42.
[19] 张会慧,张秀丽,王娟,田野,李鑫,胡举伟,孙广玉.利用快相叶绿素荧光参数综合评价3种丁香的耐盐性[J].南京林业大学学报(自然科学版),2013,37(5):13-19.
[20] 张会慧,张秀丽,李鑫,丁俊男,朱文旭,齐飞,张婷,田野,孙广玉.NaCl和Na2CO3胁迫对桑树幼苗生长和光合特性的影响[J].应用生态学报,2012,23(3):625-631.
[21] Strasser R J.Donor side capacity of photosystem Ⅱ probed by chlorophyll a fluorescence transients[J].Photosynthesis Research,1997,52(2):147-155.
[22] Henmi T,Miyao M,Yamamoto Y.Release and reactive-oxygen-mediated damage of the oxygen-evolving complex subunits of PSⅡ during photoinhibition[J].Plant and Cell Physiology,2004,45(2):243-250.
[23] 张会慧,田褀,刘关君,胡彦波,吴翔宇,田野,李鑫,孙广玉.转2-CysPrx基因烟草抗氧化酶和PSⅡ电子传递对盐和光胁迫的响应[J].作物学报,2013,39(11):2023-2029.
[24] 魏晓东,陈国祥,施大伟,刘丹,唐加红,李霞.干旱胁迫对银杏叶片光合系统Ⅱ荧光特性的影响[J].生态学报,2012,32(23):7492-7500.
[25] 叶露幻,沈唯军,郑宝刚,宋涛,陈国祥,吕川根.两优培九剑叶衰老过程中光合膜功能及蛋白复合物的变化[J].作物学报,2013,39(11):2030-2038.
(责任编辑 武艳培)
The heterosis of photosystem Ⅱ functions inSorghumbicolor×S.sudanenseseedlings under drought stress
HU Ju-wei1, ZHANG Hui-hui1,2, ZHANG Xiu-li1, PANG Hao-sheng1, SUN Guang-yu1
(1.College of Life Science, Northeast Forest University, Harbin 150040, China; 2.Institute of Natural Resources, Heilongjiang Academy of Sciences, Harbin 150040, China)
The response of photosystem Ⅱ activity in leaves ofSorghumsudanenseandS.bicolor×S.sudanenseto drought stress was investigated by the fast chlorophyll fluorescence induction dynamics analysis. The results showed that the maximum photochemical efficiency (Fv/Fm) and the performance of PSⅡ activity (PIABS) in these two species decreased with the increasing intensity of drought stress, especially the PIABSreduction was significantly greater thanFv/Fm. Although the PSⅡ photochemical activity in leaves of both of these two species seedlings decreased under drought stress, the PSⅡ photochemical activity inS.bicolor×S.sudanenseseedling leaves was significantly higher (P<0.05) than that inS.sudanenseseedling. The possible reasons for these difference included that the rates of relative variable fluorescence increase at 0.15 and 0.3 ms on the OJIP curve ofS.bicolor×S.sudanenseseedlings were significantly less than that ofS.sudanenseseedlings under drought stress and that the impacts of drought on the complex activities of electron donor side OCE oxygen and thylakoid aggregation state inS.bicolor×S.sudanenseseedlings was significantly less than those inS.sudanenseseedlings which was important for the supply, transport and stability of electron in electron transport chain. In addition, the relative variable fluorescence ofVJandVIat 2 and 30 ms on the OJIP curve (J point and I point) of these two species forage seedlings significantly increased under drought stress suggested that the decrease of PSⅡ photochemical activity was correlated with the block of QAto QBtransport in PSⅡ electron acceptor side and the decrease of the PQ library capability of accepting electron. However, there was significant difference betweenVJand no significant difference between VIofS.bicolor×S.sudanenseandS.sudanenseleaves, indicating that the electron transfer body QBability to accept electrons and the ability of transmitting electron of PSⅡ donor side ofS.bicolor×S.sudanenseseedlings were higher than those inS.sudanenseunder drought stress.
S.bicolor×S.sudanense; hybrid; drought; photosystemⅡ
SUN Guang-yu E-mail:sungy@vip.sina.com
10.11829j.issn.1001-0629.2014-0249
2014-05-16 接受日期:2014-09-04
中央高校基本科研业务费专项资金项目(2572015AA27);哈尔滨市科技创新人才研究专项(2013RFXXJ063);黑龙江省自然科学基金重点项目(ZD2011-05)
胡举伟(1988-),男,山东临沂人,在读博士生,主要从事植物生理生态学研究。E-mail:hujuwei1988@aliyun.cn
孙广玉(1963-),男,黑龙江巴彦人,教授,博导,博士,主要从事植物生理生态学研究。 E-mail:sungy@vip.sina.com
Q945.1
A
1001-0629(2015)03-0392-08
胡举伟,张会慧,张秀丽,逄好胜,孙广玉.高丹草叶片PSⅡ光化学活性的抗旱优势[J].草业科学,2015,32(3):392-399.
HU Ju-wei,ZHANG Hui-hui,ZHANG Xiu-li,PANG Hao-sheng,SUN Guang-yu.The heterosis of photosystem Ⅱ functions inSorghumbicolor×S.sudanenseseedlings under drought stress[J].Pratacultural Science,2015,32(3):392-399.
猜你喜欢
今日农业(2021年10期)2021-11-27
现代盐化工(2021年3期)2021-08-27
趣味(语文)(2019年10期)2020-01-14
食品与生物技术学报(2020年4期)2020-01-06
环球时报(2019-04-12)2019-04-12
环球人物(2018年7期)2018-04-23
新课程·下旬(2015年10期)2015-10-21
中国火炬(2015年12期)2015-07-31
中国火炬(2013年3期)2013-07-24
中国火炬(2011年8期)2011-07-25
