
青藏高原植被净初级生产力对物候变化的响应
2015-06-07 11:09:03杨柏娟,王思远,常清,孙云晓,尹航,汪萧悦
地理与地理信息科学 2015年5期
杨 柏 娟,王 思 远,常 清,孙 云 晓,尹 航,汪 萧 悦
(1.中国科学院遥感与数字地球研究所,北京 100094;2.中国科学院大学,北京 100049;3.中国地质大学,北京 100083)
青藏高原植被净初级生产力对物候变化的响应
杨 柏 娟1,2,王 思 远1*,常 清1,2,孙 云 晓1,2,尹 航1,3,汪 萧 悦1,2
(1.中国科学院遥感与数字地球研究所,北京 100094;2.中国科学院大学,北京 100049;3.中国地质大学,北京 100083)
该文开展了全球变暖影响下青藏高原植被净初级生产力(Net Primary Productivity,NPP)对物候变化响应机制的研究。首先,基于MODIS MOD13Q1 NDVI 数据集,采用动态阈值法和CASA模型分别反演了2002-2012年青藏高原植被物候和植被 NPP。而后研究了青藏高原物候和NPP时空演变格局,分析了青藏高原不同生态单元、不同植被类型的NPP对物候变化的响应机制。结果表明:1)2002-2012年青藏高原高寒植被生长初期(Beginning of Glowing Season,BSG) 提前(0.034 d/a),生长结束期(Ending of Growing Season,EGS)推后(0.299 d/a),生长期(Length of Growing Season,LGS)延长(0.300 d/a),植被 NPP与此相应出现不同程度的增加(1.494 gC·m-2/a)。2)青藏高原植被NPP对植被物候变化的响应出现明显的区域分异。整体上青藏高原植被LGS的延长促进了NPP的增加,但由于不同地区主导气象因子的差异,两者之间的响应关系也存在明显差异。3)春季物候的提前对NPP所带来的影响明显大于生长结束期,而BGS主要靠影响春季NPP的累积进而影响全年植被NPP的总量。
青藏高原;物候;净初级生产力;时空演变;响应机制
0 引言
植被物候的变化能够直观地反映全球气候变化,并引起植物生产力、结构组成及土壤-植物-大气系统水热碳交换的变化,进而影响气候系统,加剧气候变化,因而受到广泛关注。植被净初级生产力(NPP)是指绿色植物在单位面积、单位时间内由光合作用所产生的有机物总量中扣除自养呼吸后的剩余部分,不仅直接反映了植被群落在自然环境条件下的生产能力以及陆地生态系统的质量状况,也是判定生态系统碳收支和调节生态过程的主要因子,在全球变化以及碳循环中扮演着重要的角色。随着全球气候变暖,植被物候发生了显著的改变,并且这一影响在高纬度和高山陆地生态系统中尤为突出和重要[1]。目前大量针对气候变化对植被物候影响的研究表明:在过去的几十年,北半球植物生长期呈逐渐延长趋势,各地春季物候普遍提前[2,3],并且得到了地面观测资料的验证。近年来也有少量学者开展物候变化如何影响生态系统进程的研究,理论上认为气候变暖会延长植被生长期(Length of Growing Season,LGS), 进而增加植被的生产力[4,5]。然而,实际情况并非如此,野外观测表明植被物候变化对NPP的影响十分复杂,这加剧了定量评价植被生态系统碳收支的困难。如:Lieth等[6,7]通过模型研究发现落叶林随着LGS延长一天,净生态生产力增加5.2 gC·m-2和5.9 gC·m-2。然而,Dunn等[8]的研究指出,没有明确的证据表明这两者之间存在显著的关系。Richardson[9]通过通量站点数据和遥感数据提取落叶阔叶林和针叶林的物候和净生态生产力,结果表明不同植被不同区域植被净生态生产力对物候变化在时间和空间上的响应都不同。Falge等[10]利用遥感叶面积指数提取物候发现LGS每延长一天,净生态生产力减少0.84 gC·m-2。Piao等[11]利用陆地生物过程模型和遥感反演植被指数研究了北半球生态系统对秋季变暖的响应,发现秋季物候推迟导致光合作用累积量减少。综上所述,目前国内外关于植被生产力对物候响应关系的研究结果,由于数据源、研究区域的不同而存在较大差异,因此进一步开展基于卫星遥感的大区域植被物候变化与植被净初级生产力响应关系的研究具有重要意义。
青藏高原特殊的地理位置、海拔高度及地貌条件,形成了从热带到寒带、从湿润到干旱等多种气候和生态系统类型,是研究陆地生态演变的理想场所[12]。过去几十年的研究表明,全球变暖使青藏高原植被群落、物种组成和物种多样性发生了较大变化[13]。自20 世纪80 年代至今,高寒植被物候整体表现出返青期提前、枯黄期推迟、生长期延长的趋势[14],然而这种变化对植被净初级生产力的影响目前研究尚少[15],仅有的研究又存在结论不一致或矛盾的地方。因此,利用卫星遥感反演的长时间序列植被物候与NPP数据,研究青藏高原植被NPP与植被物候的变化以及不同地理单元NPP对物候变化的响应关系,显得尤为必要。
1 数据与方法
1.1 数据源及预处理
本文物候提取采用的研究数据是由NASA网站下载的2002年1月1日 至2012年12月31日MODIS MOD13Q1 NDVI 数据集,具有250 m的空间分辨率,每隔16 d一幅。该数据集由MODIS 地面反射率产品经过几何精校正、辐射矫正、大气校正预处理,采用最大值合成法(MVC)在像元水平上计算合成,进一步消除了大气、云、太阳高度角等因素的影响。同时利用MRT(MODIS Reprojection Tools)进行影像拼接和投影转换,以WGS-1984为基准面将影像重投影为Albers等面积圆锥投影。
本文用于计算NPP的相关气象数据包括月太阳辐射、月平均温度、月降水量、月平均相对湿度和月日照时数,由中国气象科学数据共享服务网提供。在对典型植被类别进行分析时,采用中科院地理科学与资源研究所提供的1∶100万青藏高原地物类型图。该数据产品受到云、汽溶胶及水汽等影响,为减少噪声,需对NDVI 时间序列数据做进一步处理。本研究在充分考虑遥感数据集及青藏高原典型植被NDVI特点的基础上,采用云、雪像元反距离加权空间插值与Chen等[16,17]提出的S-G滤波算法相结合的处理方法,对青藏高原2002-2012年NDVI时间序列数据进行逐像元重建。为了消除非植被像元的影响,在重建前去除了原始影像中全年NDVI最大值小于0.2的像元,该滤波方法有效地消除了突变噪声对NDVI数据集的影响,并很好地保留了曲线的真实信息。
1.2 动态阈值法提取植被物候
目前具有代表性的植被物候遥感反演模型有:动态阈值法、最大变化斜率法及logistic曲线拟合法。考虑到所用遥感数据的时间尺度,并经过日均温模型对3种物候提取方法结果的验证,本文最终采取动态阈值法对植被物候进行提取(图1)。
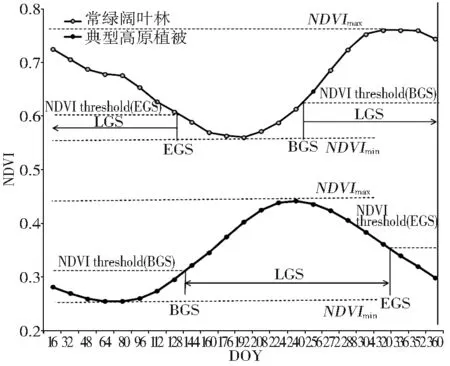
图1 动态阈值法提取青藏高原植被物候
Fig.1 Dynamic threshold phenology extraction algorithm
首先统计青藏高原主要植被类型的NDVI曲线以分析植被的生长特征,根据统计结果和相关研究的参考值设动态阈值[18,19]:
NDVIt=(NDVImax-NDVImin)×20%
(1)
其中:NDVImax为全年植被NDVI最大值,NDVImin为NDVI上升或下降阶段的最小值,分别用于确定生长初期与结束期的阈值。
假设在遥感数据时间间隔内,植被NDVI呈线性增长,采用线性插值的方法根据得到的阈值确定植物返青期和枯黄期,植物生长期即为枯黄期减去返青期。青藏高原常绿阔叶林NDVI曲线由于所处地区季节性特征不明显,与其他植被存在很大差异,进入夏季后降雨量骤增,持续的阴雨天气使得光合有效辐射明显下降,导致NDVI曲线上半年下降,下半年上升,生长期定为全年天数加上EGS与BGS的差值。鉴于常绿阔叶林的特殊性,后续讨论整体植被物候和NPP时将其排除。
1.3 CASA模型估算青藏高原植被NPP
本文基于CASA模型对植被NPP进行了估算,该模型充分考虑了植被本身的生理生态特性以及生长的环境条件,综合利用遥感数据、气象数据及植被覆盖类型数据实现植被NPP估算。CASA模型中NPP通过植被吸收光合有效辐射(APAR)和实际光能利用率(ε)计算得到[20,21]。
NPP(x,t)=APAR(x,t)×ε(x,t)
(2)
APAR(x,t)=SOL(x,t)×fPAR(x,t)×0.5
(3)
式中:APAR(x,t)是t月份x像元处植被吸收的光合有效辐射,单位为MJ/(m2·month);ε(x,t)是t月份x像元处的实际光能利用率,单位为gC/MJ;SOL(x,t)是t月份x像元处的太阳总辐射量,单位为MJ/(m2·month); fPAR(x,t)是t月份x像元处植被对入射光合有效辐射的吸收比例,无量纲;0.5表示植被所能利用的太阳有效辐射(波长为0.38~0.71mm)占太阳总辐射的比例。
植被对入射光合有效辐射的吸收比例fPAR由植被类型和植被覆盖度决定,研究表明fPAR与NDVI值之间存在良好的线性关系:
(4)
(5)
式中:SR是比值植被指数,据经验取SRmin为1.08,SRmax根据植被类型的不同,取值为4.14~6.17。
本文选择与国内外相关研究成果及地面实测资料进行NPP精度验证。经过对比,本文各植被类型NPP估算与周才平等[22,23]研究成果近似。此外与纳木错站点实测生物量数据(2007-2010年5-9月共17个实测值)对比进一步验证了NPP估算精度。两者线性拟合得到估算NPP与实测值间的关系为Y=0.832X+3.647,R2=0.672,通过了P<0.001的显著性检验。
2 结果分析
2.1 青藏高原植被物候与NPP时空演变分析
青藏高原中东部地区BGS有较大程度的提前,提前区域占51.61%,提前速率在-1~0d/a内所占比例最大(24.37%),西南地区BGS则显示出推后趋势。EGS提前区域集中在青藏高原中部,大部分地区显示出推后趋势,推后区域占青藏高原的57.53%,在青藏高原西北部推后程度最大(>1d/a)。55.31%的区域LGS显示出延长的趋势,而青藏高原中部和西南部分区域则显示出缩短趋势。上述结果与近年来国内外发表的同类研究保持一致[24-26]。
青藏高原地区年累计NPP由西向东呈现从减少到增加的变化趋势,增加趋势最多的区域集中在青藏高原中东部地区,占66.26%,增加趋势强烈的地区面积占20.25%,增加速率在0~2gC·m-2/a间的面积比例达34.49%。而在青藏高原南部NPP呈强烈减少趋势。整体上在2002—2012年间青藏高原NPP有明显的增加,各变化区间的统计数据见表1。

表1 青藏高原植被物候、NPP不同趋势变化区间所占百分比
通过计算2002-2012年青藏高原整体植被物候平均值及年均NPP,分析两者年际变化趋势(图2)。2002-2012年,青藏高原整体植被返青期开始于第111-122天,呈现提前趋势(0.034 d/a);枯黄期开始于第297-308天,呈现推迟趋势(0.299 d/a);生长期为183-193天,呈现延长趋势(0.300 d/a),但物候年际变化并不显著。平均年NPP在219.802~242.543 gC·m-2之间,增长趋势显著(1.494 gC·m-2/a,P=0.007)。
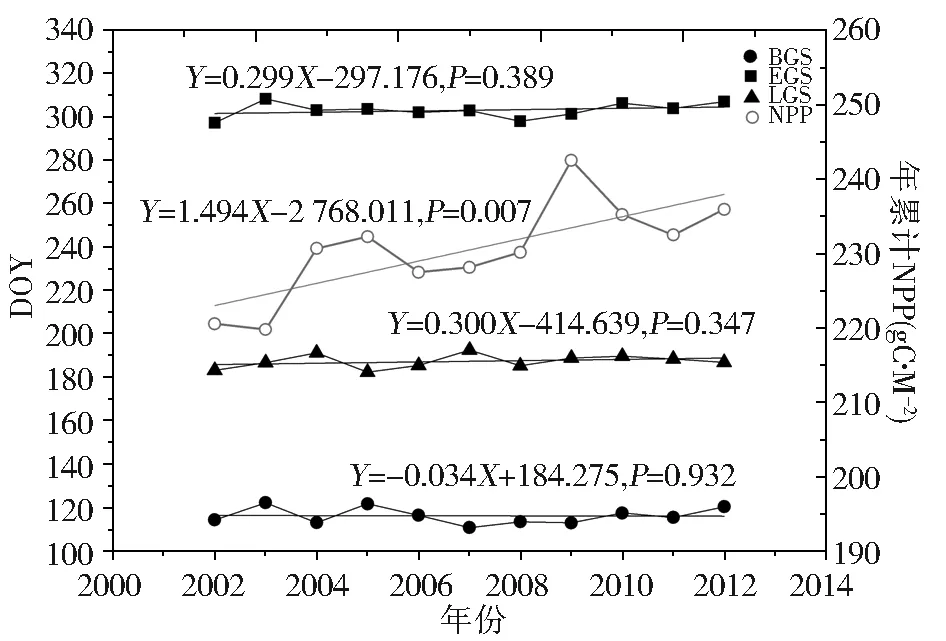
图2 青藏高原整体植被物候、年累计NPP年际变化
Fig.2 Interannual variability of overall vegetation phenology and NPP on Tibet Plateau
分析不同植被类型(针阔混交林、常绿阔叶林、灌丛、高寒草原、高寒草甸/苔原及高山稀疏植被)的物候及NPP的年际变化特征可知:针阔混交林BGS微弱提前(0.013 d/a),EGS推后(0.465 d/a)、LGS延长(0.177 d/a),NPP呈减少趋势(0.876 gC·m-2/a),波动幅度较大。其年NPP与LGS经相关性统计分析,显示出强烈的正相关性(r=0.729,P=0.011)。常绿阔叶林返青期晚,有提前趋势(0.180 d/a),EGS也有所提前(1.001 d/a),生长期缩短(0.456 d/a)。常绿阔叶林NPP呈现减少趋势,但并不显著(0.492 gC·m-2/a),波动幅度在所有植被类型中最大。灌丛BGS提前(0.086 d/a),EGS推后(0.275 d/a),LGS延长(0.301 d/a),NPP呈现增加趋势0.085 gC·m-2/a)。高寒草原BGS推后(0.170 d/a),EGS推后(0.100 d/a),LGS延长(0.260 d/a),NPP显著增加(0.896 gC·m-2/a ,P=0.001)。高寒草甸和苔原BGS提前(0.086 d/a),EGS推后(0.275 d/a),LGS延长(0.301 d/a),NPP显著增加 (3.459 gC·m-2/a,P=0.006)。高山稀疏植被BGS有所推后(0.103 d/a),但由于EGS的推迟(0.354 d/a),LGS仍然延长(0.235 d/a),NPP也呈现显著增加的趋势(0.982 gC·m-2/a,P=0.004)。
植被物候及NPP的年际变化因植被类型不同而不同。针阔混交林、常绿阔叶林和灌丛主要分布于青藏高原东南部,这3种植被类型的NPP年累积量高,年际波动大;高寒草原、高寒草甸和苔原及高山稀疏植被位于青藏高原中部和西部,虽然物候变化不尽相同,但NPP累积量都有显著的增加。鉴于如此明显的区域分异,下文将对不同地理单元植被物候与NPP的响应关系展开分析。
2.2 NPP对生长季长度变化的响应分析
由于植被生长季长度直接影响植被NPP的累积,本文着重分析NPP与植被生长季长度变化的响应关系,并将响应关系分为4类:A:LGS缩短,NPP随之减少;B:LGS延长,但NPP减少;C:LGS 缩短,NPP反而增加;D:LGS延长,NPP随之增加(图3)。
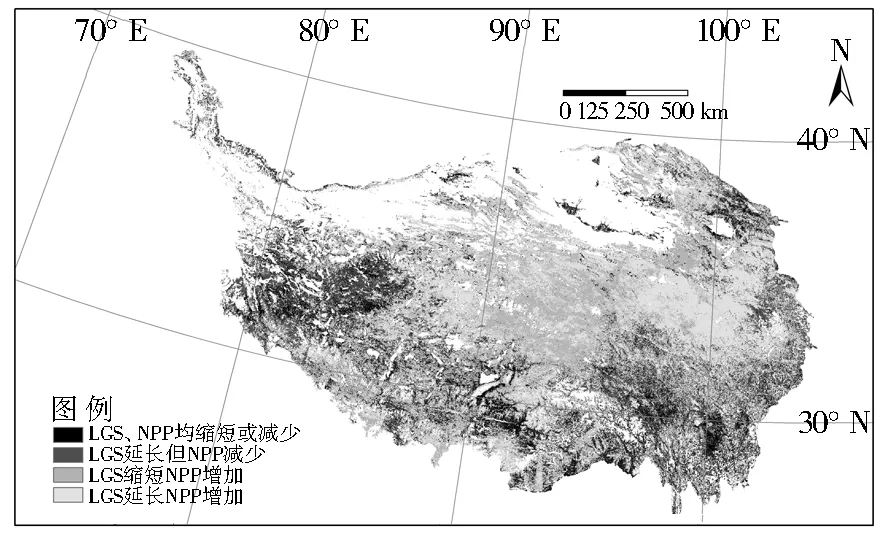
图3 青藏高原NPP与LGS响应的空间差异
Fig.3 Spatial variabilty of the relationship between NPP and LGS
情形A主要位于青藏高原南部的灌丛区,地处高原温带湿润/半湿润地区,水热条件好,年降水量达700 mm以上,植被生产力水平常年较为稳定,该区2002-2012年降水量相比其他地区减少最多。生长期缩短(BGS推迟1.400 d/a,EGS提前0.378 d/a)的趋势下,灌丛NPP也显著减少,减少速率为4.359 gC·m-2/a(P<0.01)。B情形主要位于青藏高原西部——高原温带干旱地区和高原亚寒带半干旱地区羌塘高原湖盆高寒草原,高寒草原在返青期提前和生长结束期推后的双重作用下(BGS提前0.942 d/a,EGS推后1.296 d/a)整个物候期有了较大程度的延长。标准化之后温度、降水与NPP的复相关系数分别为-0.079和0.780,降水量为影响该区域植被生长的主导因子。降水量下降,温度升高又促进了水分蒸发,尽管生长期有所延长,它对植被生产力累积的促进作用因植被自身消耗增大而被抵消, NPP减少速率为-0.870 gC·m-2/a(P<0.01)。C情形主要位于青藏高原中部,主要植被类型为高寒草甸和苔原。该区年降水量基本维持在500 mm左右,标准化之后温度、降水与NPP的复相关系数分别为0.673和0.269,气温对该区域植被生产力水平的影响占主导地位,并且近10年该区域温度增加最为显著,而降水量略微下降,植被生产力提高。李英年等[27]通过模拟实验也发现,生长期气温的升高导致矮蒿草草甸植物发育速度加快提前成熟,生长季变短。而提前结束的生长期避免了秋季增温所增加的自身消耗,LGS缩短一天,NPP增加1.271 gC·m-2(P<0.01)。D情形主要位于青藏高原东部——高原温带半干旱草原区和亚寒带半干旱高寒草甸草原地区,主要植被类型为高寒草甸/苔原及高寒草原,且所占面积比例最大(38.27%)。D情形所处区域年均温和年降水量均高于C情形所处区域,标准化之后温度、降水与NPP的相关系数分别为0.582和0.543,植被生产力对温度变化的敏感程度相对于C情形下降,对降水敏感度上升。LGS延长,降水增加,从而NPP显著增加,增加速率为5.942 gC·m-2/a(P<0.01)。
在不同温度、降水区间NPP随LGS有不同的变化规律。以2℃为间隔划分年均温区间,总体上温度越高NPP值越大,当年均温在-6℃以下时,年NPP累积量基本在100 gC·m-2以下,并且NPP与LGS显著负相关(r=-0.810,P<0.05),LGS越长,NPP累积反而越少。年均温在-6~2℃时,NPP随着LGS的延长先增大,后减小,转折点随着温度升高而向后推移;年均温在2℃以上时,NPP随着LGS的延长先有略微的减小,随后急剧增大。年降水量在800 mm以下时,NPP总体上随着降水量增加而增加;年降水量超过800 mm时,过高的降水反而抑制了NPP的累积;年降水量在300 mm以内时,NPP随LGS变化幅度小;年降水量在300~400 mm时,NPP随LGS的延长先增大后减小;年降水量在400~600 mm时,LGS越长NPP累积越多;年降水量超过600 mm,以LGS 165 d为转折点,NPP先减少后增加。水热条件好的区域,LGS越长则NPP累积量越多。
2.3 植物生长初期、生长结束期对NPP的影响
植被生长季的开始与结束同时影响着整个LGS,进而影响植被的生产力。理论上,春季物候主要影响上半年生产力,而秋季物候则对下半年植被生产力影响较大。为深入了解上述关系,本文分析了青藏高原整体及各植被类型BGS、EGS分别对春季(3-5月)、秋季(9-11月)和全年NPP的影响。
首先计算BGS、EGS与全年NPP之间的相关性。57.4%的地区全年NPP与BGS间呈现负相关关系,其中对BGS提前敏感的区域集中在青藏高原东部,负相关性显著。55.78%的地区全年NPP与EGS之间呈现负相关关系,主要集中在青藏高原中部地区的高寒草甸和苔原。这与上文该地区LGS由于EGS提前而缩短、NPP反而增加的结论相一致。全年NPP与EGS正相关响应较为强烈的区域在青藏高原高寒草甸和苔原的东南部分。
BGS、EGS与NPP之间统计相关特性如表2。整体看,青藏高原地区植被生长初期与春季NPP有着显著的负相关关系(P=0.003),而EGS对秋季NPP和全年NPP的影响都没有BGS对春季NPP和全年NPP的影响大。就各植被类型看,针阔混交林BGS对全年NPP有着较大的影响,相关系数为-0.672,显著性达到0.023。常绿阔叶林由于上文提到的BGS/EGS与其他植被类型相反,BGS与秋季NPP显著相关,相关系数为-0.807。灌丛、高寒草原、高寒草甸和苔原及高山稀疏植被BGS都与春季NPP显示出显著的负相关性。除了高寒草原,剩余植被类型的EGS对NPP的影响则没有达到显著相关水平。
表2 植被生长初期、生长结束期对NPP的影响
Table 2 The effects of BGS and EGS on NPP
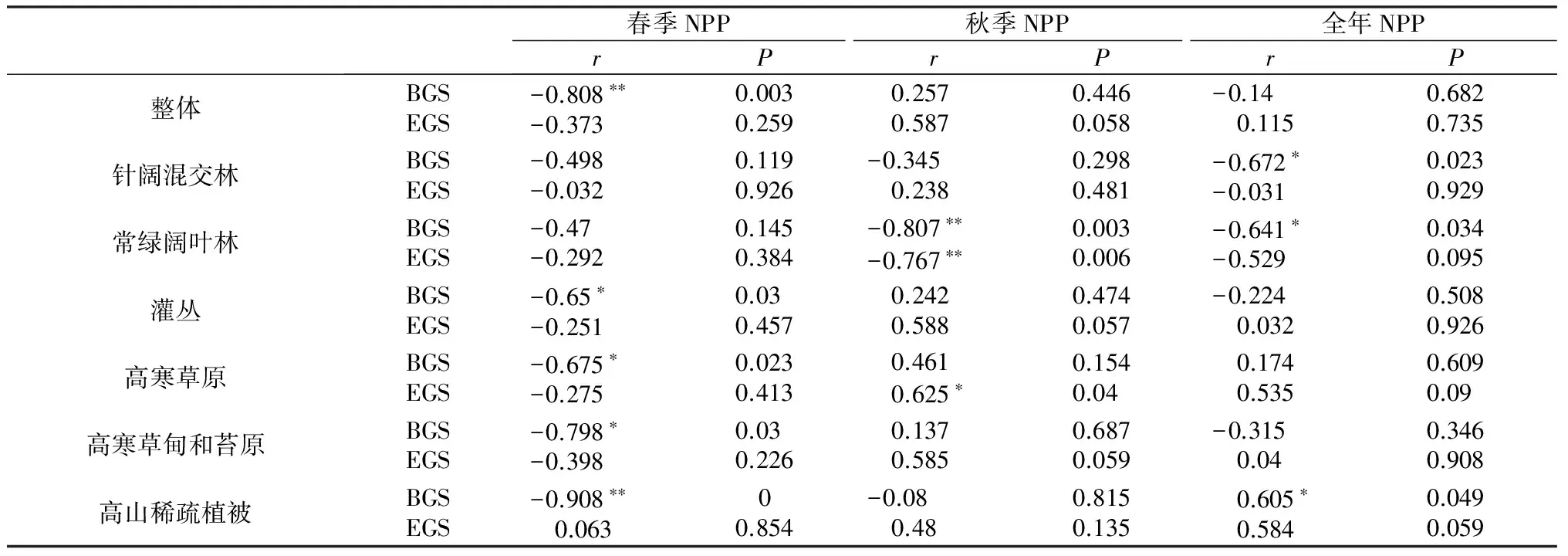
春季NPP秋季NPP全年NPPrPrPrP整体针阔混交林常绿阔叶林灌丛高寒草原高寒草甸和苔原高山稀疏植被BGSEGS-0.808∗∗-0.3730.0030.259 0.257 0.5870.4460.058-0.14 0.1150.6820.735BGSEGS-0.498-0.0320.1190.926-0.345 0.2380.2980.481-0.672∗-0.0310.0230.929BGSEGS-0.47-0.2920.1450.384-0.807∗∗-0.767∗∗0.0030.006-0.641∗-0.5290.0340.095BGSEGS-0.65∗-0.2510.03 0.457 0.242 0.5880.4740.057-0.224 0.0320.5080.926BGSEGS-0.675∗-0.2750.0230.413 0.461 0.625∗0.1540.04 0.174 0.5350.6090.09 BGSEGS-0.798∗-0.3980.03 0.226 0.137 0.5850.6870.059-0.315 0.040.3460.908BGSEGS-0.908∗∗ 0.06300.854-0.08 0.480.8150.135 0.605∗ 0.5840.0490.059
注:*P<0.05;**P<0.01。
3 结论与展望
青藏高原物候和NPP整体上随着水热梯度变化由西北向东南呈现出一定的变化特征。总体看,2002-2012年间青藏高原植被物候BGS提前、EGS推后、LGS延长,而NPP呈显著增长趋势。LGS的延长对NPP的增加有一定促进作用,但由于各植被类型的差异和不同生态地理单元特征,NPP对LGS变化的响应不同,即使相同的植被类型,在不同的气候环境下NPP与LGS的响应也表现出明显的区域分异。如在青藏高原东南部水热条件良好的区域,随LGS缩短NPP减少4.359 gC·m-2/a;但在年降水量小于500 mm的区域,在气候暖干化过程中,尽管LGS延长,NPP减少-0.870 gC·m-2/a,且降水为影响NPP的主导因子[23]。 春季NPP对BGS的变化有明显的响应,而植被NPP对EGS变化的敏感度没有BGS大[28,29]。返青期的提前加之气温的升高加速了植被生长发育,从而提高了上半年的生产力。
研究结果表明,NPP年际变化与物候变化间的关系较为复杂,这与植被类型、生态地理单元及气候特征的不同有很大关系。本文在研究NPP对物候变化的响应机制时仅考虑了温度、降水因子以及部分植被要素、地理单元的影响,不同地形特征、积雪冻土环境以及人类活动等对物候与NPP响应机制的作用有待进一步分析。基于不同的地理单元、气象水文特征、地形地貌特征甚至人类活动影响下开展全面的植被物候与植被生产力之间的响应关系研究将对青藏高原生态系统变化和预测有着重大意义。
[1] 陈建国,杨扬,孙航.高山植物对全球气候变暖的响应研究进展[J].应用与环境生物学报,2011,03:435-446.
[2] 丁明军,张镱锂,孙晓敏,等.近10年青藏高原高寒草地物候时空变化特征分析[J].科学通报,2012,33:3185-3194.
[3] BADECK F W,BONDEAU A,BOTTCHER K,et a1.Responses of spring phenology to climate change[J].New Phytologist,2004,162:295-309.
[4] HU J,MOORE D J P,BURNS S P,et a1.Longer growing seasons lead to less carbon sequestration by a subalpine forest [J].Global Change Biology,2010,16:771-783.
[5] DRAGONI D,SCHMIDW H P,CRAIG A.Evidence of increased net ecosystem productivity associated with a longer vegetated season in a deciduous forest in south-central Indiana,USA[J].Global Change Biology,2011,17:886-897.
[6] LIETH H.Modeling primary productivity of the world[J].Primary Productivity of the Biosphere,1975,14:237-263
[7] BALDOCCHI D D,WILSON K B.Modeling CO2and water vapor exchange of a temperature broadleaved forest across hourly to decadal time scales[J].Ecol.Modell.,2001,142:155-184.
[8] DUNN A L,BARFORD C C,WOFSY S C,et al.A long-term record of carbon exchange in a boreal black spruce forest:Means,responses to interannual variability,and decadal trends[J].Global Change Biol.,2007,13:577-590.
[9] RICHARDSON A D.Influence of spring and autumn phonological transitions forest ecosystem productivity[J].Phenology and Forest Productivity,2010,365:3227-3246.
[10] FALGE E,BALDOCCHI D,TENHUNEN J,et al.Seasonality of ecosystem respiration and gross primary production as derived from FLUXNET measurements[J].Agricultural and Forest Meteorology,DOI:10.1016/S0168-1923(02)00102-8.
[11] PIAO S L.Net carbon dioxide losses of northern ecosystems in response to autumn warming[J].Nature,2008,451:49-52.
[12] 莫申国,张百平,程维明,等.青藏高原的主要环境效应[J].地理科学进展,2004,23(2):88-96.
[13] XU M H,XUE X.A review of effects of climate warming on terrestrial plant-soil ecosystem[J].Chinese Bulletin of Life Sciences,2012,24:492-500.
[14] 宋春桥,游松财,柯灵红,等.藏北高原植被物候时空动态变化的遥感监测研究[J].植物生态学报,2011,35(8):853-863.
[15] 李英年,王启基.气候变暖对高寒草甸气候生产潜力的影响[J].草地学报,2000,8(1):23-29.
[16] CHEN J J,NSSON P,TAMURA M,et al.A simple method for reconstructing a high-quality NDVI time-series data set based on the Savitzky-Golay filter[J].Remote Sensing of Environment,2004,91(3):332-344.
[17] BIAN J,LI A,SONG M,et al.Reconstruction of NDVI time-series datasets of MODIS based on Savitzky-Golay filter[J].Journal of Remote Sensing,2010,14(4):22-27.
[18] JÖNSSON P,EKLUNDH L.TIMESAT—a program for analyzing time-series of satellite sensor data[J].Computers & Geosciences,2004,30(8):833-845.
[19] YU H,LUEDELING E,XU J.Winter and spring warming result in delayed spring phenology on the Tibetan Plateau[J].Proceedings of the National Academy of Sciences of the United States of America,2010,107(51):22151-22156.
[20] FIELD C B,RANDERSON J T,MALMSTRM C M,et al.Global net primary production:Combining ecology and remote sensing[J].Remote Sensing of Environment,1995,51(1):74-88
[21] YANG H,MU S,LI J,et al.Effects of ecological restoration projects on land use and land cover change and its influences on territorial NPP in Xinjiang,China[J].CATENA,2014,115:85-95.
[22] 周才平,欧阳华,王勤学,等.青藏高原主要生态系统净初级生产力的估算[J].地理学报,2004,1:74-79.
[23] 陈卓奇,邵全琴,刘纪远,等.基于MODIS的青藏高原植被净初级生产力研究[J].中国科学(地球科学),2012,3:402-410.
[24] SHEN M G,ZHANG G X,CONG N,et al.Increasing altitudinal gradient of spring vegetation phenology during the last decade on the Qinghai-Tibetan Plateau[J].Agricultural and Forest Meteorology,2014,189-190:71-80.
[25] DING M,ZHANG Y,SUN X,et al.Spatiotemporal variation in alpine grassland phenology in the Qinghai-Tibetan Plateau from 1999 to 2009[J].Chinese Science Bulletin,2013,58(3):396-405.
[26] 游松财,宋春桥,柯灵红,等.基于 MODIS 植被指数的藏北高原植被物候空间分布特征[J].生态学杂志,2011,30(7):1513-1520.
[27] 李英年,赵亮,赵新全,等.5年模拟增温后矮蒿草草甸群落结构及生产量的变化[J].草地学报,2004,3:236-239.
[28] NIEMAND C,KÖSTNER B,PRASSE H,et al.Relating tree phenology with annual carbon fluxes at Tharandt forest[J].Meteorologische Zeitschrift,2005,142:197-202.
[29] RICHARDSON A D,HOLLINGER D Y,DAIL D B,et al.Influence of spring phenology on seasonal and annual carbon balance in two contrasting New England forests[J].Tree Physiology,2009,29:321-331.
Response of NPP to Phenology Changes in the Tibet Plateau
YANG Bai-juan1,2,WANG Si-yuan1,CHANG Qing1,2,SUN Yun-xiao1,2,YIN Hang1,3,WANG Xiao-yue1,2
(1.InstituteofRemoteSensingandDigitalEarth,ChineseAcademyofSciences,Beijing100094; 2.UniversityofChineseAcademyofSciences,Beijing100049;3.ChinaUniversityofGeosciences,Beijing100083,China)
It is already an indisputable fact that the phenology of high-latitude areas has changed to a different extent following global warming.The research of this phenomenon is important for us to predict the response of terrestrial ecosystem to global change.However,the knowledge about the effects of phenology changes on biomass especially on vegetation net primary productivity (NPP) is hardly known.This paper took the Tibet Plateau (TP) as study area for its unique natural geographical environments and aimed to study the response mechanism between phenology and NPP.Firstly,phenology and NPP were detected respectively with dynamic threshold value method and CASA model based on MODIS NDVI dataset and meteorological data from 2002 to 2012 in the Tibet Plateau.Then,the paper described the spatio-temporal trends of phenology and NPP,and analyzed the impacts of phenology changes on NPP of different areas and vegetable types.The research also discussed the temperature and precipitation in different areas to better explain the response mechanism diversities.Conclusions can be drawn as follows:1) The beginning of growing season (BGS) of overall vegetations in Tibet Plateau has advanced during 2002 to 2012 at the rate of 0.034 d/a;the end of growing season(EGS) has delayed 0.299 d/a;the length of growing season(LGS) extended 0.300 d/a,and the annual NPP has increased 1.494 gC·m-2·a-1significantly.2) The extending of LGS can contribute to the increasing of NPP overall the Tibet Plateau.However this correlation is not notable and has regional differences because of the changing dominant meteorological factors.3) The advance of BGS has greater impact on NPP than the delay of EGS,and the effect of BGS mainly lies in the stimulation to the spring NPP.
Tibet Plateau;phenology;NPP;spatio-temporal evolution;response mechanism
2015-02-09;
2015-03-22
国家自然科学基金项目“青藏高原植被物候变化及其生物量响应机制研究”(D010702)
杨柏娟(1990-),女,硕士研究生,研究方向为植被物候与全球变化。*通讯作者E-mail:wangsy@radi.ac.cn
10.3969/j.issn.1672-0504.2015.05.023
X87;P463
A
1672-0504(2015)05-0115-06
猜你喜欢
今日农业(2022年3期)2022-11-16 13:13:50
热带作物学报(2022年6期)2022-07-08 06:45:10
甘肃林业(2019年4期)2019-10-08 07:19:52
西南农业学报(2017年5期)2017-06-23 08:14:35
红土地(2016年3期)2017-01-15 13:45:17
河北林业科技(2016年5期)2016-11-08 03:12:55
西南农业学报(2016年5期)2016-05-17 05:42:22
海峡姐妹(2015年9期)2015-02-27 15:13:07
生物进化(2014年2期)2014-04-16 04:36:27
杭州科技(2014年3期)2014-02-27 15:26:46
