
烟粉虱成虫在不同木薯品种上的空间分布型研究
2015-05-30 14:20段春芳宋记明沈绍斌严炜刘倩李月仙娄予强熊贤坤周迎春易怀锋张林辉刘光华
热带作物学报 2015年8期
段春芳 宋记明 沈绍斌 严炜 刘倩 李月仙 娄予强 熊贤坤 周迎春 易怀锋 张林辉 刘光华


摘 要 烟粉虱(Bemisia tabaci)是木薯上一种重要的害虫,为明确烟粉虱在木薯上的空间分布,及时采取有效的防控措施,在田间调查的基础上,采用聚集度指标、Iwao的M*-m回归法、Taylor幂法则、Lα-m回归模型和Z-V模型对烟粉粉虱成虫在不同木薯品种上的空间分布型进行了研究。结果表明木薯上的烟粉虱成虫在密度较低时呈现均匀分布,密度高时聚集分布,且其聚集是由环境因素造成的。在木薯叶层上的分布总体呈现数量上层>中层>下层,在SC11、GR4和SC124上烟粉虱成虫密度较大,SC8上密度最低。
关键词 木薯;烟粉虱;空间分布
中图分类号 S533;S435.6 文献标识码 A
Study on Spatial Distribution Patterns of Bemisia tabaci Adults in Different Cassava Species
DUAN Chunfang, SONG Jiming, SHEN Shaobin, YAN Wei, LIU Qian, LI Yuexian,
LOU Yuqiang, XIONG Xiankun, ZHOU Yingchun, YI Huaifeng, ZHANG Linhui, LIU Guanghua*
Tropical and Subtropical Cash Crops Research Institute, YAAS, Baoshan, Yuannan 678000,China
Abstract The whitefly, Bemisia tabaci, is a major pest of cassava, In order to make a cher underatanding of the spatial distribution of Bemisia tabaci on cassava and to carry out control methods effectively and timely, using the spatial distribution patterns of Bemisia tabaci adults in different cassava Species were analyzed by different aggregation indices, Taylorpower law, Iwaos M*-m regression analysis, Lans Lα-m model and Zhangs Z-V model.The results showed that the spatial distribution pattern of B. tabaci adults distributed evenly when its population density was low, but sggregated when its population density was high. The numbers of B. tabaci adults on the different leaf layers were significantly different, with the sequence of upper leaves>middle leaves>lower leaves, and B. tabaci adults mainly clustered on the leaf back of cassava. The density of B. tabaci adults on SC11, SC124 and GR4 were the most, SC8 was the least.
Key words Cassava;Bemisia tabaci;Spatial distribution pattern
doi 10.3969/j.issn.1000-2561.2015.08.016
烟粉虱(Bemisia tabaci Gennadius)属同翅目粉虱科,是一种世界性害虫,在热带、亚热带及温带地区均有分布[1]。烟粉虱在我国一年发生11~15代,世代重叠严重,其寄主较广泛,可多达74科500余种[2-3]。烟粉虱在刺吸植物汁液的同时还传播植物病毒,另外分泌的蜜露可诱发煤烟病,在木薯上,烟粉虱还可传播非洲木薯花叶病毒,是一种重要的害虫[4]。昆虫空间格局反映了昆虫在寄住植物和非生物环境影响下,空间需求的内在生物学特性[5],包括种内种间关系、对环境的适应、行为特征、遗传特征等。研究昆虫的空间分布型,可以进一步认识种群在田间的分布特征[6],并对昆虫田间防治有着重要的意义[7]。目前,国内外对烟粉虱在不同寄主植物上的空间分布进行了一些研究,研究多见于棉花[8-9]、蔬菜[10-12]和花卉[13-14]类寄主植物。然而,关于木薯上烟粉虱的空间分布格局还未见报道。为此,本研究旨在探明烟粉虱在木薯上的空间分布结构,获得准确和系统的田间调查资料,从而为预测预报和防治提供科学依据。
1 材料与方法
1.1 调查地概况
该木薯地位于云南省农业科学院热带亚热带经济作物研究所木薯科技示范园,木薯面积为8 hm2,调查木薯品种有SC5、SC11、SC124、SC205、GR4和新选048。
1.2 方法
1.2.1 虫口密度调查方法 采用栅格式取样法[15],每个木薯品种各选5行,每行随机调查20株,每株分上、中、下3层,每层选4片叶,轻轻翻动叶片,分别记录每叶烟粉虱成虫数量,用于空间分布型计算。调查时间为2014年6月27~28日,调查时木薯平均株高:SC5:69.3 cm;SC11: 89.7 cm; SC124: 97.6 cm;SC205: 113.4 cm;GR4: 122.4 cm;新选048: 104.1 cm。
1.2.2 分析方法 用Excel 2003和DPS v 7.0处理田间调查的数据。
烟粉虱种群的聚集程度[16-17]。根据扩散系数C、Ca指数、聚集指数I、聚集度指标M*/M、Lα指标等来分析不同木薯品种上烟粉虱种群的空间分布格局。
Iwao回归法[18]。用Iwao的M*-m回归模型, 求出平均拥挤度与平均数的回归关系M*=α+βm。
Taylor幂法则[19]。用Taylor幂法则求出平均数与方差的回归关系lg(v)=lga+blg(m)。
兰星平[18]的Lα-m回归模型。Lα指标:La=m-m/υ+1,式中m为样本平均值,υ为样本方差。Lα-m回归模型:La=θ+ηm,式中m为样本平均值,θ、η为引入的参数。
张连翔[20]Z-V模型。Z指标:Z=V/m-1+V,V为方差,m为样本平均值。Z-V模型:Z=A+BV,式中v为样本方差。
聚集原因的分析。根据Blackith[21]提出种群聚集均数(λ)来分析昆虫分布的聚集原因:
λ=■× r
上式中m为平均虫口密度,K为负二项分布参数值,r为具有自由度等于2K的X2 0.5(卡方) 分布的函数值, 根据公式计算出不同木薯品种烟粉虱成虫的λ值。
1.2.3 数据处理 采用方差分析比较烟粉虱在不同品种木薯上不同层次上的分布情况和差异。
2 结果与分析
2.1 聚集度指标法测定
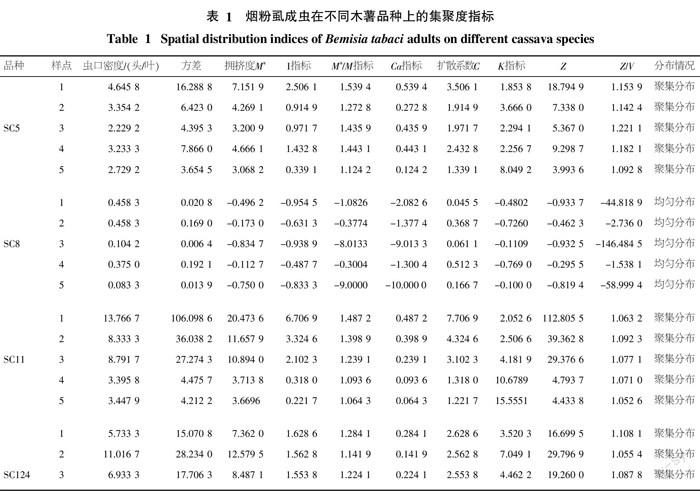
各个木薯品种田间聚集度各项指标如表1,SC8木薯品种上烟粉虱成虫的田间分布各项指标为C<1、I<0、K<0、Ca<0、M*/m<1、Z/V<1,为均匀分布,其余木薯品种上烟粉虱成虫的各项聚集度指标均达到C>1、I>0、K>0、Ca>0、M*/m>1, Z/V>1,符合种群聚集分布的检验标准。从各木薯品种上烟粉虱成虫的平均数来看,SC8上密度较低。这说明,烟粉虱成虫在不同虫口密度上呈现不同的分布特征,在低密度上呈现均匀分布,随着虫口密度的增加上升而聚集度增加,并趋向聚集分布状态。
2.2 回归模型分析
用Iwao M*-m回归法、Taylor幂法则、Lα-m回归模型和Z-V模型得到的不同木薯品种烟粉虱成虫回归方程见表2,从采用的4种回归模型结果来看,Iwao M*-m回归法、Taylor幂法则得到的结果一致,但在Taylor方法和Iwao方法的判断系统中却找不到a<0,b>1和α<0,β>1的组合型式,所以无法通过这2种回归模型判断该木薯地烟粉虱成虫的空间分布型,这进一步说明了Taylor方法和Iwao方法存在的局限性[20]。采用兰星平和张连翔等提出的模型得出,除SC8是均匀分布以外,烟粉虱成虫在其他木薯品种上均为聚集分布。这与聚集度指标测定结果相一致。从Lα-m回归模型和Z-V模型相关系数r值来看,此次木薯烟粉虱空间分布型研究以Lα-m回归模型为佳。
2.3 聚集均数λ分析
在SC8上的Blackkith种群聚集数λ<2,其聚集是由于某些环境如气候、栽培条件、植株生育状况等所引起的,其余木薯品种均λ>2,说明烟粉虱成虫在木薯上聚集是由烟粉虱本身的聚集行为与环境条件综合影响的结果。
2.4 烟粉虱成虫在木薯不同层次上的分布
烟粉虱成虫在不同木薯品种上部和中部叶片上的数量最大,木薯下部叶片上数量较少,往下烟粉虱成虫数量减少。SC5上中部与下部、SC11上部与下部、SC124上中部与下部有显著差异(表3)。这说明烟粉虱成虫主要集中在木薯上中部嫩叶片上,这是因为烟粉虱喜欢在幼嫩叶片上取食和产卵造成的。
2.5 烟粉虱成虫在不同木薯品种上的分布
不同木薯上烟粉虱成虫虫口密度如表4。从表中看出,SC8与SC11、SC124、GR4差异显著,其中,SC8号木薯上虫口密度最低,仅为0.30头/叶,SC11、GR4和SC124木薯品种虫口密度最大,达7.55头/叶、7.55头/叶和7.43头/叶。调查时发现,GR4木薯上虫口密度最大可达到125头/叶,SC11木薯上虫口密度最大可达到103头/叶,其次是SC124木薯上虫口密度最大可达到57头/叶。
3 讨论与结论
研究表明,烟粉虱成虫在木薯上种群密度较低时,呈均匀分布,随着烟粉虱成虫密度增加而呈聚集分布。这与烟粉虱在棉田[8]、番茄[21]上的空间格局研究结果相似。烟粉虱成虫在木薯植株叶片上部叶片数量最大,这与烟粉虱在茄子、西瓜、甜瓜、黄瓜[10]和棉花[8]上的分布规律基本相似。从木薯品种上烟粉虱虫口密度来看,SC11、GR4和SC124 3个木薯品种虫口密度最大,SC8木薯烟粉虱密度最低。该研究只分析了木薯烟粉虱成虫的空间分布格局,烟粉虱的发生规律以及烟粉虱卵、若虫的时空动态还需要进一步研究,再者,木薯烟粉虱在整个木薯种植区的发生危害还需调查研究,这样更好的为木薯烟粉虱的发生危害积累一定的数据依据,并为木薯烟粉虱的危害数据填补空白。根据本文的研究结果,在木薯烟粉虱的防治过程中应该以树冠中上部为防治重点。
在昆虫种群空间格局研究中,回归模型分析法被认为是较新较好的方法[20],其中,应用最广泛的主要是Iwao的M*-m回归分析和Taylor幂法则[16],但在一些实际应用中,Iwao模型和Taylor模型出现某些不灵验的现象[22-26],本研究中采用改进过的Lα-m模型和Z-V模型,较Iwao模型和Taylor模型能很好的对烟粉虱成虫空间格局做出分析。
参考文献
[1]Boykin L M, Shatters R G Jr, Rosell R C, et al. Global relationships of Bemisia tabaci(Hemiptera: Aleyrodidae)revealed using Bayesian analysis of mitochondrial COI DNA sequences[J]. Molecular Phylogenetics and Evolution,2007, 44(3): 1 306-1 319.
[2]Stansly P A, Naranjo S E. Introduction, XV-XVIII[M]. //PA Stansly and SE Naranjo(eds), Bemisia: Bionomics and management of a global pest. Springer, New York,2010:540.
[3]Li S J,Xue X,Ahmed M Z,et al. Host plants and natural enemies of Bemisia tabaci(Hemiptera: Aleyrodidae)in China[J]. Insect Science, 2011, 18(1):101-120.
[4]黄贵修, 李开绵. 中国木薯主要病虫草害识别与防治[M]. 北京: 中国农业科学技术出版社, 2012.
[5] Taylor L R. Aggregation, variance and the mean[J]. Nature, 1961, 189: 732-735.
[6] 李培兴, 李 云, 张春海,等. 桃蚜在桃园的种群消长动态和空间分布型研究[J]. 山东农业科学, 2009, 4: 83-85.
[7] 黄立飞,杨 朗,姜建军. 桑园主要害虫的空间分布型研究[J]. 中国农学通报, 2011, 27(31): 280-283.
[8] 周福才, 任顺祥, 杜予州, 等. 棉田烟粉虱种群的空间格局[J]. 应用生态学报, 2006, 17(7): 1 239-1 244.
[9] 杨益众, 吴 娟, 余月书,等. 棉株上烟粉虱若虫种群的垂直分布与统计方法研究[J]. 昆虫知识, 2004, 41(4): 331-332.
[10] 沈斌斌, 任顺祥, Musa, P. D等. 烟粉虱成虫空间分布格局型的研究[J]. 昆虫知识, 2005, 42(5): 544-546.
[11] 牛新利,巫厚长,邹运鼎,等. 黄瓜上烟粉虱成虫种群空间分布格局及时序动态[J]. 中国农学通报, 2007, 23(7): 457-461.
[12] 汪恩国, 陈林松, 蒋尚军,等. 番茄地烟粉虱空间格局参数特征及其应用[J]. 植物保护, 2007, 33(6): 113-116.
[13] 黄 俊, 张 娟, 李明江, 等. B型烟粉虱成虫在一品红上垂直分布规律[J]. 浙江农业学报, 2011, 23(4): 748-753.
[14] 姚士桐,郑永利. 大棚非洲菊烟粉虱成虫空间分布模型及抽样技术研究[J]. 浙江农业学报, 2007, 19(3): 233-236.
[15] 赵志模. 地质统计学理论与方法及其在生态学中的应用[M]. 重庆: 西南师范大学出版社, 2006.
[16] 徐汝梅. 昆虫种群生态学[M]. 北京: 北京师范大学出版社, 1987: 7-33.
[17] 兰星平. Lα-m模型在检验昆虫种群空间分布型与抽样调查中的应用[J]. 林业科学研究, 1996, 9(1): 41-46.
[18] Iwao S. Application of the M*-M method to analysis of spatial pattern by changing the Quadra size[J]. Res Popul Eco, 1972, 14(1): 97-128.
[19] Taylor L R. Agg regation, va riance and them ean[J]. Nature, 1961, 189: 7 325.
[20] 张连翔, 吕尚彬, 温豁然,等. 种群空间格局研究的Z-V模型及其抽样设计方法[J]. 西北林学院学报, 1997, 12(1): 75-79.
[21] 汪恩国, 陈林松, 蒋尚军,等. 番茄地烟粉虱空间格局参数特征及其应用[J]. 植物保护, 2007, 33(6): 113-116.
[22] 徐汝梅, 刘来福,丁岩钦. 改进的Iwao m*-m模型[J]. 生态学报, 1984, 4(2): 111-118.
[23] 张振田,张联中. 楸螟在苗田的空间分布型及其应用的研究[J]. 昆虫知识, 1995, 32(1): 32-34.
[24] 兰星平, 魏义贤,朱秀娥. 松毒蛾空间分布型的研究[J]. 贵州林业科技, 1992, 20(1): 8-19.
[25] 张庆贺, 刘篆芳,初 冬. 落叶松八齿小蠹雄虫空间分布型及群居危害机理[J]. 东北林业大学学报, 1989, 17(6): 51-57.
[26] 敬甫松. 大螟在玉米田的空间格局及其抽样研究[J]. 昆虫知识, 1987, 24(2): 71-78.
猜你喜欢
古今农业(2021年1期)2021-07-20
世界热带农业信息(2019年7期)2019-08-30
世界热带农业信息(2018年11期)2018-01-17
广东第二课堂·小学(2017年5期)2017-05-27
安徽农学通报(2016年24期)2017-01-12
绿色科技(2016年20期)2016-12-27
中国民族民间医药·上半月(2016年10期)2016-11-19
科技视界(2016年18期)2016-11-03
科技视界(2016年13期)2016-06-13
世界热带农业信息(2014年8期)2014-09-23
