
鸡开产日龄和开产体质量的主基因+多基因混合遗传分析
2015-05-28 07:40:32王克华窦套存沈曼曼胡玉萍
江西农业大学学报 2015年3期
马 猛,王克华 ,曲 亮,窦套存,沈曼曼,郭 军,卢 建,胡玉萍
(1.江苏省家禽科学研究所,江苏 扬州 225003;2.扬州翔龙禽业发展有限公司,江苏 扬州 225200)
鸡的开产日龄和开产体质量是影响蛋鸡育种和生产的重要指标。开产日龄不仅影响蛋鸡开产后产蛋率上升的速度及产蛋高峰期的持续时间,而且对整个产蛋周期中的产蛋量也有一定的影响[1],开产日龄和开产体质量的过大或者过小对蛋鸡的蛋质量和产蛋量均具有较大的影响[2-8]。有研究表明经典的遗传学方法只能在总体上对基因的效应和作用方式进行估计,但是却不能区分不同基因在效应上的差别[9]。盖钧镒等[10]、Zhang 等[11]、Wang 等[12]提出的主基因+多基因混合遗传分析方法不仅能够鉴别主基因,而且对多基因也可以进行检测,且能够估计出相应的遗传参数。本研究利用主基因+多基因混合遗传模型来对绿壳蛋鸡黑羽纯系和白来航鸡资源群体的开产日龄和开产体质量进行分析,对蛋鸡开产日龄和开产体质量的遗传规律进行探讨,确定开产日龄和开产体质量的最适合模型,为蛋鸡的育种提供一定的理论指导。
1 材料与方法
1.1 试验材料
试验以黑羽绿壳蛋鸡纯系为亲本P1,以白来航鸡为亲本P2,其中P1共109只,P2共66只,F1(P1♂×P2♀、P2♂×P2♀)代 513 只,F2[(P1♂×P2♀)♂×(P1♂×P2♀)♀、(P2♂×P1♀)♂×(P2♂×P1♀)♀]代1 744只。试验动物均饲养于扬州翔龙禽业发展有限公司,饲养管理条件一致,试验过程中每只鸡均单笼饲养。
1.2 测定内容
试验中记录每个世代的每只母鸡产下第1枚蛋的时间,开产时间为鸡产下第1枚蛋时间减去出雏时间;同时测量鸡产下第1枚蛋时的体质量,即为开产体质量。
1.3 数据的处理
试验所有数据均采用Excel 2003进行统计,用SPSS17.0进行正反交群体间的t检验,用南京农业大学研发的SEA-G4F2软件包进行主基因+多基因混合遗传模型分析。
数据进行分析前对F1和F2代的数据进行正反交之间的比较,如果正反交之间差异显著,则需要消除正反交效应。
正交效应=正交群体平均值-群体平均值(1)
反交效应=反交群体平均值-群体平均值 (2)
用于分离分析的数据=原数据-正反交效应 (3)
1.4 遗传模型简介
利用P1、P2、F1和F23个世代进行分离分析的遗传模型共有五大类24个遗传模型,其具体对应的解释见表1。
1.5 统计与分析
应用盖钧镒等[10]提出的主基因+多基因混合遗传模型来进行分离分析,通过极大似然法和IECM算法对混合分布中的有关成分分布参数做出估计,然后对24个模型中AIC值(Akaike’s information criterion)进行比较,选择其中AIC值较小的相对应的模型进行适合性检验,然后确定出最适模型,根据最适模型的分析结果,估计其相应的一阶和二阶遗传参数。具体的最适模型的选择和遗传参数估计参照文献[10]。适合性检验共有5个统计量,即均匀性检验U21、U22和U23,Smirnov检验nW2和 Kolmigorov检验 Dn。

其中h2mg为主基因遗传率,h2pg为多基因遗传率。
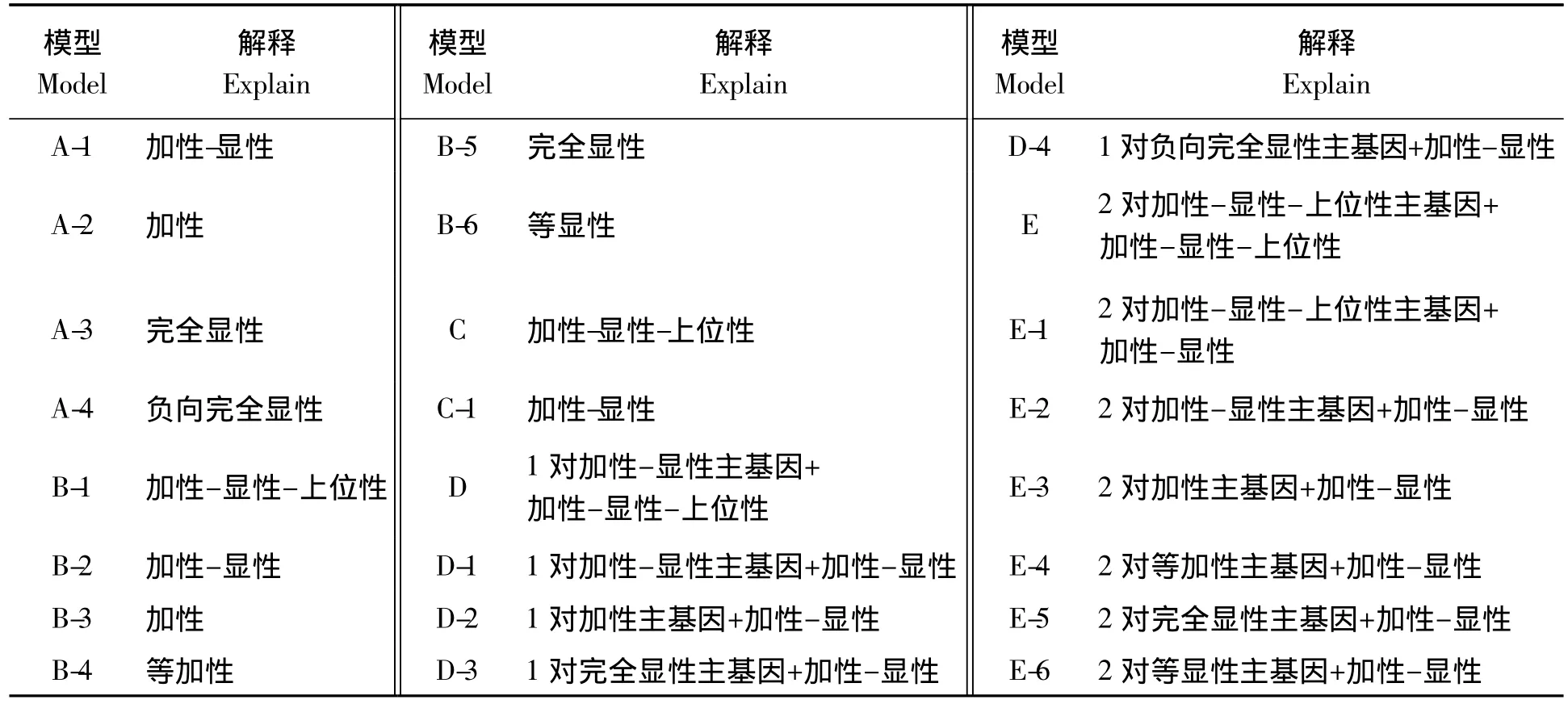
表1 各个遗传模型对应的解释Tab.1 Explain of each geneticalmodel
2 结果与分析
2.1 开产日龄和开产体质量的次数分布
开产日龄和开产体质量的次数分布见表2和表3,由表2,3知,2个亲本的开产日龄均要大于F1和F2代的开产日龄,亲本P1和P2开产日龄的均值差异不显著(P>0.05),F1和F2的开产日龄均值均要小于亲本的,说明F1和F2代的开产日龄具有超亲遗传。F2代的次数分布主要呈现单峰正态分布。开产体质量的2个亲本的开产体质量的均值分别为1 102 g和1 320 g,亲本P2的开产体质量明显大于亲本P1的开产体质量,F1和F2代的开产体质量分别为1 118 g和1 175 g,介于2个亲本之间。F2的次数分布主要呈现出单峰偏态分布。

表2 开产日龄的次数分布Tab.2 Frequency distributions for the age at first egg

表3 开产体质量的次数分布Tab.3 Frequency distributions for the body weight at first egg
2.2 最适模型的确定及适合性检验
开产日龄和开产体质量的各个模型及其所对应的AIC值见表4。由表4可知,开产日龄的所有模型中模型D、E和E-1的AIC值分别为20 915.84、20 865.8和20 954.48,选择这3个模型作为期望模型,进一步进行适合性检验后发现,模型D的适合性检验统计量有9个值达到了显著水平,模型E有7个统计量达到了显著性水平,模型E-1有13个统计量达到了显著性水平,因此,根据最适模型的选择标准,选择统计量显著性个数较少的模型E作为最适模型,且模型E的AIC值最小,因此选择模型E为开产日龄的最适模型,即2对加性-显性-上位性主基因+加性-显性-上位性多基因混合遗传模型。开产体质量的24个模型中AIC值较小的模型为B-1、E和E-1,其对应的AIC值分别为34 717.05、34 710.72和34 707.34,因此选择模型B-1、E和E-1作为期望模型进行下一步的适合性检验,检验结果为模型B-1的检验统计量有2个达到了显著性水平,模型E和E-1各有1个检验统计量达到了显著性水平,但是模型E-1的AIC值最小,因此选择模型E-1作为开产体质量的最适模型,即2对加性-显性-上位性主基因+加性-显性多基因混合遗传模型。开产日龄和开产体质量的最适模型的适合性检验见表5。
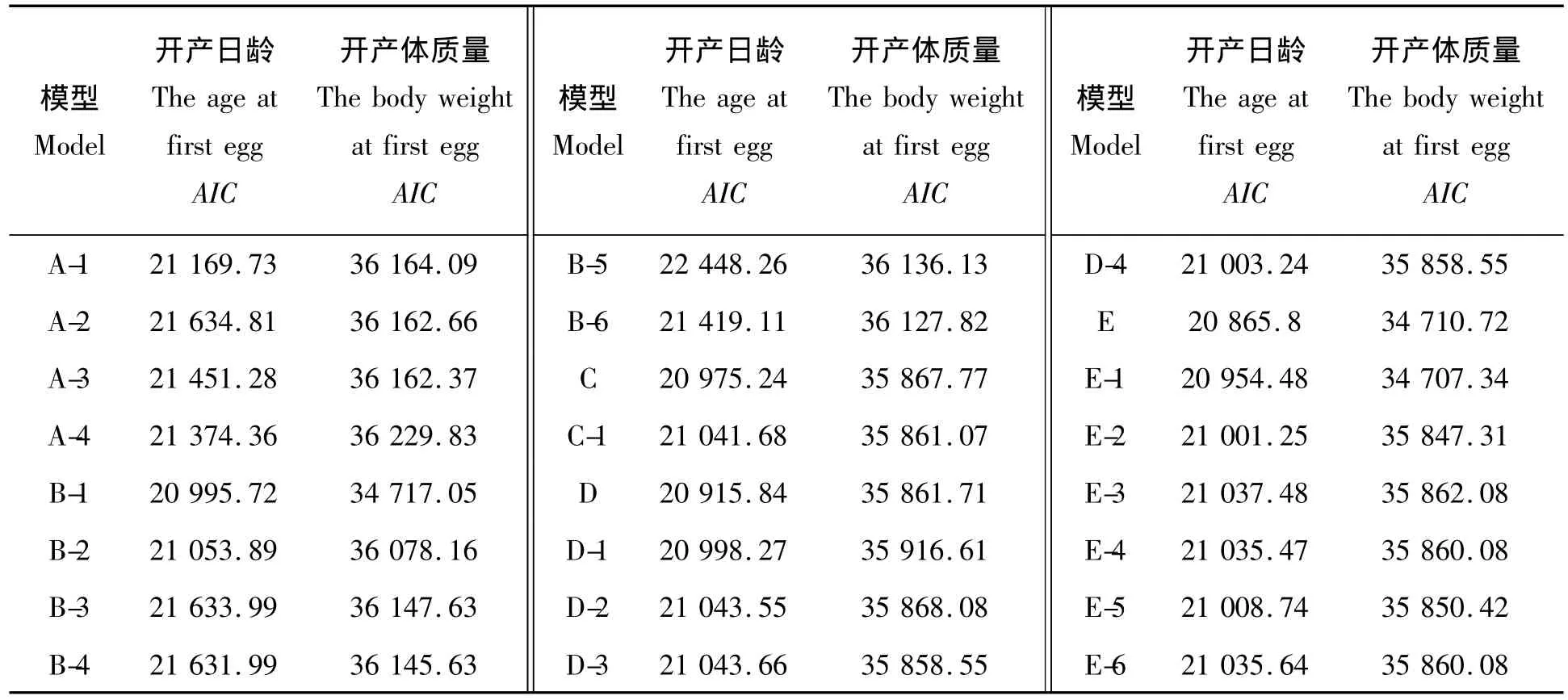
表4 开产日龄和开产体质量各个模型所对应的AIC值Tab.4 The Akaike’s information criterion(AIC)values in each model for the age and body weight at first egg

表5 开产日龄和开产体质量最适模型的适合性检验Tab.5 Tests for goodness-of-fit of bestmodel for the age and body weight at first egg
2.3 开产日龄和开产体质量的遗传参数估计
由表6可知,在开产日龄的一阶遗传参数的估计中,2对主基因的加性效应之间和显性效应之间差别很小,分别为6.25和6.25、-6.25和-6.20。AA 的加性效应和 BB 的加性效应相等,均为6.25,Bb 的显性效应和加显效应也相等,估计值均为-6.20;二阶遗传参数的估计中,总体方差为125.54,主基因的方差为31.46,多基因的方差较小,几乎为0,主基因的遗传率25.06%。在开产体质量的一阶遗传参数中,2对主基因 AA 和 BB的加性效应分别为593.23和585.51,Aa和 Bb的显性效应为-14.91和70.44,加加效应和显显效应分别为12.84和-70.95,加显效应和显加效应分别为-499.85和-581.89,多基因的加性效应和显性效应分别为-1 287.39和-64.23。在二阶遗传参数中,总体的方差为31 787.27,主基因的方差为20 114.99,主基因的遗传率为63.28%,2个性状的主基因遗传率均远远大于多基因的遗传率,说明开产日龄和开产体质量主要是由主基因控制的。
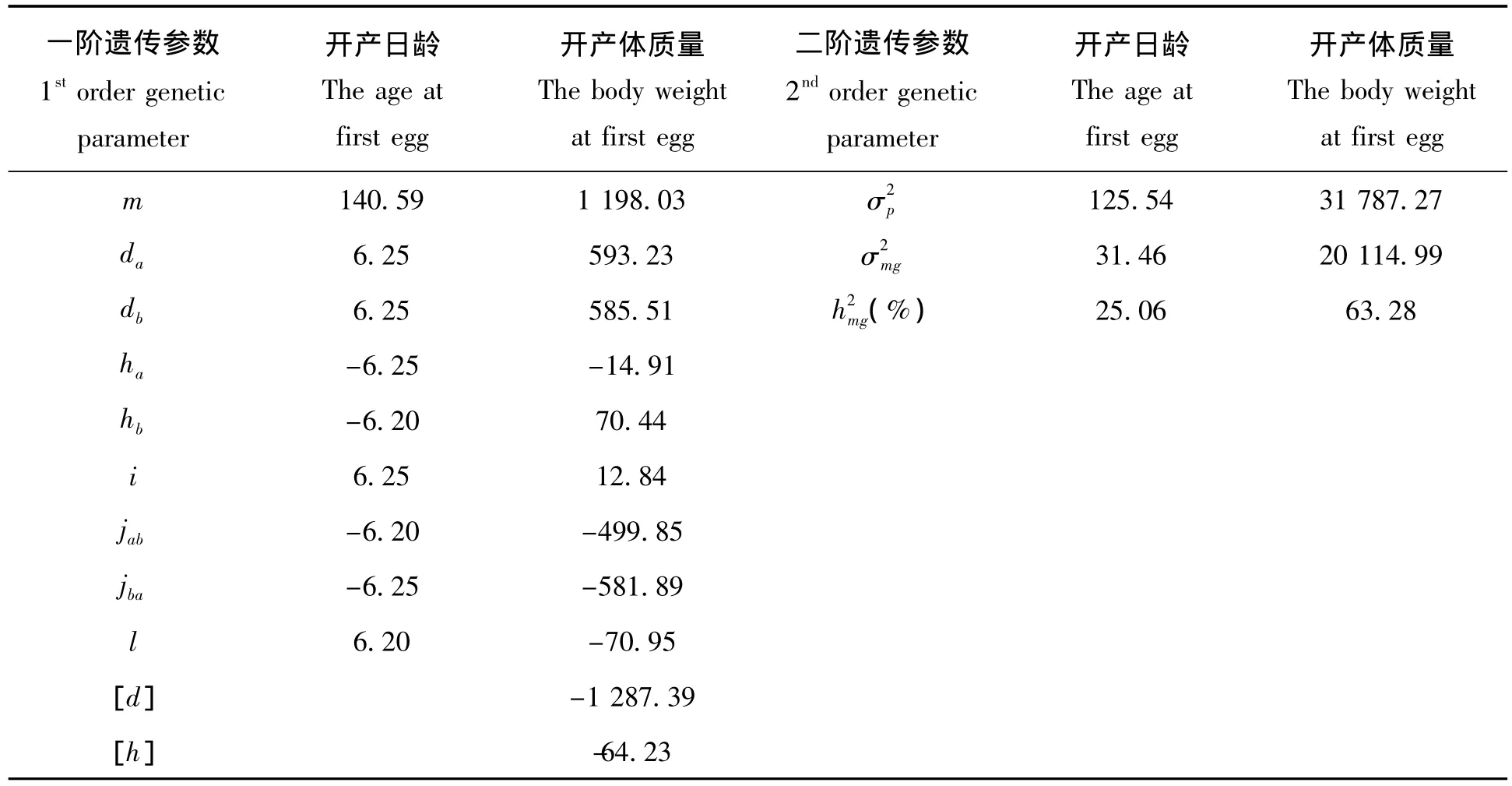
表6 开产日龄和开产体质量的遗传参数估计Tab.6 Estimates of genetic parameters for the age and body weight at first egg
3 讨论与结论
目前,关于主基因+多基因的应用在植物数量性状的遗传研究上报道的较多[13-15],且有研究表明用QTL定位检测出来的植物数量性状的主基因的数量与用主基因+多基因混合遗传模型分析出来的主基因对数是一样的[16]。但是目前将该模型应用于动物数量性状遗传的研究报道较少,王克华等[17]用仙居鸡和隐性白羽鸡为素材对鸡12周龄体质量进行主基因+多基因混合遗传模型分析,认为鸡12周龄体质量的最适模型为E-6模型。曲亮等[18-19]对鸡的40周龄蛋质量、冠长、冠高和冠厚进行主基因+多基因的遗传模型分析,认为40周龄蛋质量、冠长、冠高和冠厚的最适模型均为E模型。
本研究利用主基因+多基因混合遗传模型对鸡开产日龄和开产体质量进行分析,结果表明开产日龄的最适模型为模型E,即2对加性-显性-上位性主基因+加性-显性-上位性多基因混合遗传模型,开产体质量的最适模型为E-1,即2对加性-显性-上位性主基因+加性-显性多基因混合遗传模型。开产日龄和开产体质量的2对主基因的加性效应均为正值,说明开产日龄和开产体质量越大的个体对后代的影响较大。开产日龄的2对主基因的加性效应值之和与显性效应值之和相加基本可以抵消,但是开产体质量的2对主基因和多基因的加性效应之和远远大于显性效应之和,说明开产体质量主要受加性效应的互作影响,其次才是显性效应的互作。开产日龄和开产体质量的主基因遗传率分别为25.06%和63.28%,这些主基因的发现以及加性、显性、上位性等遗传效应的明确,对理解鸡开产日龄和开产体质量的遗传特征,指导鸡的育种实践具有重要意义。研究表明,鸡开产日龄的遗传力为0.15~0.54,开产体质量遗传力为0.18~0.56[20-26],与前人的研究结果相比,本研究的开产日龄的遗传力处于前人研究的范围内,开产体质量的遗传力要高于前人的研究,其原因可能是不同鸡种和不同算法之间的差异造成的。
虽然主基因+多基因混合遗传模型应用很广泛,但是还是有一定的局限,目前只能够分析1~3对主基因的遗传,而对于3对以上的主基因则不能够进行判别,但是其能够计算出各个性状的一阶和二阶遗传参数,这对数量性状的遗传效应研究,与主基因紧密连锁的分子标记的发掘和分子标记辅助选择都能够奠定一定的理论基础。
[1]李俊营,詹凯,李绍全,等.淮南麻黄鸡生长曲线拟合及开产性状相关性分析[J].畜牧与兽医,2012,44(8):46-48.
[2]周海龙,朱庆.丝羽乌骨鸡早期产蛋性能的月产蛋模型拟合分析[J].中国家禽,2004,26(2):9-10.
[3]潘剑平.蛋鸡开产前期体增重比率与产蛋性能的关系[J].山东家禽,1998(3):8-9.
[4]顾亚玲,龙翔,杨天平.AA肉种鸡体重与产蛋量相关性研究[J].内蒙古畜牧科学,2003,24(2):11-12.
[5]张永英王保安,海兰灰育成蛋鸡不同体重对生产性能的影响[J].兽药与饲料添加剂,2001,6(4):14.
[6]赵振华,黎寿丰,黄华云,等.邵伯鸡母系8周龄,20周龄体重对部分产蛋性状的影响[J].华北农学报,2011,25(12):174-176.
[7]张德祥,王谦,张细权,等.土鸡开产日龄双向选择效果分析[J].中国家禽,2012,34(24):15-17.
[8]赵振华,黄华云,张静,等.邵伯鸡母系开产日龄对早期产蛋性状的影响[A]//杨宁,李辉主编.中国家禽科学研究进展——第十四次全国家禽科学学术讨论会论文集[C].哈尔滨:中国农业科学技术出版社,2009:612-614.
[9]赵建军.白菜类作物重要农艺性状的遗传分析[D].北京:中国农业科学院,2005.
[10]盖钧镒,章元明,王建康.植物数量性状遗传体系[M].北京:科学出版社,2003:169-219.
[11]Zhang Y M,Gai JY.The EIM algorithm in the joint segregation analysis ofquantitative traits[J].Genetical Research,2003,81(2):157-163.
[12]Wang J,Fodlied DW,Cooper M,etal.Power of the joint segregation analysismethod for testingmixedmajor-gene and polygene inheritancemodels of quantitative traits[J].Theoretical and Applied Genetics,2001,103(5):804-816.
[13]陈学军,方荣,周坤华,等.辣椒果实性状主基因+多基因遗传分析[J].西北植物学报,2012,32(2):246-251.
[14]陈凤真.西葫芦产量性状主基因-多基因混合遗传分析[J].华北农学报,2011,26(3):79-85.
[15]包和平,毕成龙,李颖,等.爆裂玉米百粒重性状的主基因+多基因混合遗传分析[J].华北农学报,2011,26(3):199-203.
[16]王春娥,盖钧镒,傅三雄,等.大豆豆腐和豆乳得率的遗传分析与 QTL定位[J].中国农业科学,2008,41(5):1274-1282.
[17]王克华,章元明,曲亮,等.鸡12周龄体重的主基因+多基因混合遗传分析[J].江西农业大学学报,2011,33(1):123-127.
[18]曲亮,王克华,窦套存,等.鸡第二性征的主基因+多基因混合遗传分析[C].扬州:第十六次全国动物遗传育种学术讨论会论文集,2011:508.
[19]曲亮,马猛,王克华,等.鸡40周龄蛋重主基因-多基因混合遗传模型分析[J].中国家禽,2014,36(7):10-12.
[20]黎寿丰,陈宽维,丁余荣,等.“邵伯草鸡”母系 (S2系)主要产蛋性状的遗传参数估测[J].中国家禽,2004,26(17):13-14.
[21]王克华,窦套存,曲亮,等.如皋黄鸡产蛋性能与遗传参数分析[J].中国畜牧杂志,2010(9):12-14.
[22]宋素芳,康相涛,赖葆春,等.豫州褐壳蛋鸡Ⅰ系主要经济性状的遗传分析[J].河南农业大学学报,2003,37(3):278-282.
[23]宋素芳,康相涛,李明,等.豫州褐壳蛋鸡Ⅱ系部分数量性状的遗传分析[J].中国家禽,2004,8(1):110-113.
[24]宋素芳,康相涛,黄修奇,等.豫州褐壳蛋鸡Ⅲ系部分数量性状遗传参数的估测[J].西北农林科技大学学报:自然科学版,2003,31(1):121-123.
[25]皮劲松,杜金平.金水乌鸡部分产蛋性状遗传参数分析[J].中国家禽,1999,21(2):6-7.
[26]曾华,王谦,温美燕,等.优质肉鸡早熟性状的遗传分析[A]//杨宁.中国家禽业——机遇与挑战——第十三次全国家禽学术讨论会论文集[C].郑州:中国农业科学技术出版社,2007.
猜你喜欢
今日农业(2022年2期)2022-11-16 12:29:47
今日农业(2021年7期)2021-11-27 13:44:48
养猪(2020年1期)2020-02-19 04:18:04
今日农业(2019年11期)2019-08-15 00:56:32
国外畜牧学·猪与禽(2018年11期)2018-05-14 11:14:22
消费导刊(2017年24期)2018-01-31 01:29:31
辽宁大学学报(哲学社会科学版)(2017年3期)2017-06-21 21:16:59
少儿科学周刊·少年版(2015年11期)2015-12-17 23:47:17
少儿科学周刊·儿童版(2015年11期)2015-12-17 03:53:46
农村农业农民·B版(2015年9期)2015-10-16 16:10:38
