
草鱼幼鱼游泳能力及游泳行为试验研究
2015-05-24 01:49:19吴一红白音包力皋穆祥鹏
中国水利水电科学研究院学报 2015年3期
龚 丽,吴一红,白音包力皋,穆祥鹏
(中国水利水电科学研究院 流域水循环模拟与调控国家重点实验室,北京 100038)
草鱼幼鱼游泳能力及游泳行为试验研究
龚 丽,吴一红,白音包力皋,穆祥鹏
(中国水利水电科学研究院 流域水循环模拟与调控国家重点实验室,北京 100038)
在28±1℃水温下,利用自制鱼类游泳行为试验装置,采用流速递增法,研究了体长5.0~15.0 cm草鱼幼鱼的游泳能力和游泳行为。结果表明:该体长范围内的草鱼幼鱼临界游速在0.68~1.0 m/s之间,临界游速基本随鱼类体长增加而线性增长;在整个试验过程中,根据鱼类对水流流速的游泳行为响应,鱼类的摆尾频率随着水流速度的增加而线性增加,整个游泳过程分为4个阶段,每个游泳阶段的最大流速依次为临界游速的45.6%、82.8%、95.9%和100%。本研究成果可为四大家鱼游泳能力进行有益补充,也可为鱼道流速设计提供游泳能力参数。
游泳能力;游泳行为;草鱼幼鱼;临界游泳速度;鱼道
1 研究背景
据第一次全国水利普查统计,我国已建水库大坝高达9.8万座,是世界上建造水库大坝最多的国家,另有过流能力不低于1 m3/s的闸、堰等水利设施27万多座[1]。这些水利工程在实现水资源开发利用的同时,也给河流生态环境带来一系列负面影响,特别是阻断了河道的纵向连通性,使河流水生生境片段化,隔断了鱼类洄游通道,阻碍了鱼类种质交流,使鱼类产卵、索饵等洄游难以进行,致使江河鱼类资源逐年衰退,这种情况不仅在我国相当严重,在亚洲其他一些国家以及欧洲、澳大利亚及美洲地区也都普遍存在[2]。鱼道作为一种生态工程措施,可以保持河流的纵向连通性,满足鱼类生长繁衍的洄游需要,已广泛应用于江河开发的生态保护[3]。鱼道设计的关键之一是使鱼道流速与过鱼对象的游泳能力相匹配,保证鱼类能够克服水流顺利上溯。不同鱼种、不同体长的鱼类游泳能力各不相同,开展鱼类游泳行为研究对于指导鱼道设计尤为重要。
以往我国的鱼道水力学研究主要针对鱼道的水工构筑物流态进行模拟、分析和优化,缺乏对实际过鱼对象的喜好水流和游泳能力的研究,鱼道水力设计多以前人相关的鱼类游泳能力研究成果为依据。如国外主要过鱼对象一般为鲑科鱼类等个体较大,克流能力强的洄游性鱼类,而我国过鱼对象一般是珍贵鱼类和鲤科鱼类,其个体较小,克流能力小,对复杂流态的适应能力较差,且不同鱼种、不同体长的鱼类游泳能力差别很大,水温也是影响鱼类游泳能力的敏感因素。鱼道设计如果缺乏对实际环境条件下过鱼对象游泳能力的研究,就会缺乏针对性,难以达到较好的过鱼效果。对鱼道设计而言,鱼类临界游速是最重要的游泳能力评价指标[4-5]。鱼道设计需以过鱼对象的临界游速作为参考依据,当鱼道流速在临界游速以下时,鱼类可在较长时间内持续游泳而不会产生生理压力,这对于确定鱼道池室的最大流速和休息池间距都具有重要的参考价值。
四大家鱼是我国内陆水域主要的鱼类种群,有着重要的生态和经济价值,它们广泛分布于长江、珠江、黑龙江和黄河等流域水系。四大家鱼属半洄游性鱼类,具有典型的江湖洄游特性,其性成熟个体在江河流水中产卵,产卵后的亲鱼和幼鱼进入支流及通江湖泊中摄食育肥。因此,不仅成鱼具有产卵洄游需求,其幼鱼的摄食、育肥等生理行为也有洄游需求。随着近十几年来水电工程的大量兴建,四大家鱼种群出现了明显退化,据调查统计显示,由于葛洲坝水利枢纽兴建,水库前端的一些产卵场已很少有家鱼活动[6]。三峡大坝的修建,更加剧了四大家鱼数量的衰减,据2013年长江三峡工程生态与环境监测公报显示,2012年5—7月,三峡坝下监利江段家鱼鱼苗径流量为1.21亿尾,比上年减少71.7%,仅为蓄水前(1997—2002年)平均值的4.8%,苗汛过程不明显[7],大规模水电开发已对长江四大家鱼的资源量产生了严重影响,开展四大家鱼保护已刻不容缓。针对四大家鱼,特别是幼鱼开展鱼类游泳能力研究,为鱼道建设提供水流设计依据,是一项重要的研究内容。
目前,水产部门对四大家鱼的耗氧率、生理和养殖等方面已有很多研究成果,国内外有关草鱼行为方面的研究包括:温度对草鱼分布及行为的影响、溶氧浓度对草鱼幼鱼快速启动能力的影响、运动消耗对草鱼幼鱼游泳能力的影响、草鱼快速启动过程的加速—滑行游泳行为和草鱼幼鱼感应流速的比较等内容[8-11]。在众多研究中,虽有提到8~9.7 cm的草鱼幼鱼在18±1℃的条件下临界游速为7.11±0.6 BL/s[9],但鱼类的游泳能力与鱼种类、体长和过鱼季节水温存在着密不可分的关系。
据胡茂林等人2007—2008年调查发现,草鱼幼鱼进入鄱阳湖口的时间主要集中在7月中旬至8月底,湖口平均水温约为28~30℃左右,体长以5.00~14.99 cm为主[12]。以往的研究尚无法反映草鱼幼鱼在索饵洄游时期的实际游泳能力。因此,本文以5~15 cm的草鱼幼鱼为研究对象,在温度为28±1℃的条件下,利用自制的鱼类游泳行为的试验装置对草鱼幼鱼游泳能力和游泳行为进行较为系统的研究。
2 鱼类游泳能力试验方法
2.1试验用鱼试验用鱼由北京市某渔场提供,采集时间为2014年7月中旬,体长为5~15 cm,主要有4种体长范围:6.20±0.45 cm、7.50±0.35 cm、9.80±0.80 cm、12.30±0.80 cm,体重为1.84~19.92 g,鱼体长和体重的关系如图1所示。试验鱼的肥满度为0.97±0.10,说明4种体长范围的草鱼幼鱼生长和健康良好。
将试验鱼放置于2.0×1.5×1.0 m的矩形水池中暂养2周,暂养水为曝气5 d的自来水,水温为28± 1℃,溶解氧浓度维持在6.50 mg/L以上,氨氮浓度在0.01 mg/L以下,光照为室内自然光,试验前2 d停止喂食。
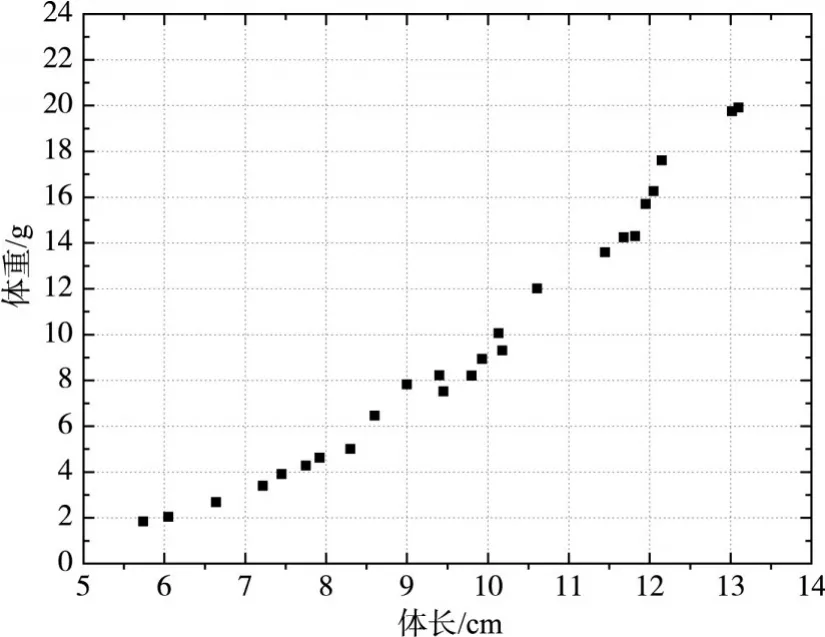
图1 试验草鱼样本的体长与体重的关系
2.2试验装置试验装置为一密闭循环水槽,如图2所示。水槽截面为25 cm×25 cm,试验观测段长80 cm。由电机带动螺旋桨产生循环水流,水流通过试验观测段前端的管簇稳流装置后,可在鱼类游泳区获得均匀、稳定的流场。通过变频控制器来调节电机转速,以调控水槽流速,采用毕托管对试验段的断面流速进行标定,确定变频电机频率与水槽流速之间的关系曲线,试验水槽的试验流速范围为0~1.3 m/s。在装置上方和正侧面各安置一个摄像机,记录鱼类游泳的全过程,以分析鱼类的游泳能力和游泳行为。
2.3试验设计与方法本试验水温为28±1℃,与实际情况基本一致。试验鱼样本共25尾,其中预试验样本3尾,正式试验22尾。
本试验中,临界游泳速度的试验方法采用递增流速法。首先进行3次预试验,对试验鱼的临界游速进行摸底,每次选取1尾鱼放入试验段进行预实验,试验前使试验鱼在1 BL/s的流速下适应1 h(BL为试验鱼体长),然后每2 min增加0.4 BL/s[13],直至鱼疲劳,记录此时的流速,并将3次预试验所得到的流速平均值作为临界游速的预估值,供正式试验参考。试验鱼疲劳判断标准为:试验鱼被水流冲至试验段下游钢丝网上无法游动的时间超出20 s。
正式试验时,将单尾试验鱼放入试验段中,使其在1 BL/s的流速下适应1 h以消除转移过程对鱼体的胁迫。测试开始后,每隔5 min增加0.5 BL/s的水流速度至60%的预估临界游泳速度,然后每隔20 min增加15%的预估游速[11],同时观察记录鱼的游泳行为,直至试验鱼疲劳无法继续游动,此时结束试验,记录此时水流速度和游泳时间,测试完成后,记录试验鱼的体长和体重。
临界游速按公式(1)计算[14]:

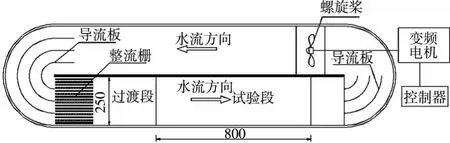
图2 鱼类游泳能力试验装置(单位:mm)
式中:Ucrit为临界游泳速度(m/s);U为试验鱼疲劳时对应的最大水流速度(m/s);ΔU为流速,即增速15%的临界游速预估值(m/s);t为该时段鱼疲劳时所用的时间(min);Δt为时间间隔20 min。
相对临界游速指用鱼类体长来表征鱼类个体的临界游泳速度,其计算公式如下:

式中:U′crit为相对临界游泳速度(m/s),BL为鱼类体长(cm)。
3 结果与分析
3.1草鱼幼鱼的临界游泳速度通过试验得到4类体长范围的草鱼幼鱼临界游泳速度分别为0.70± 0.02 m/s、0.79±0.03 m/s、0.88±0.1 m/s、0.95±0.05 m/s,对应的相对临界游泳速度分别为10.83±0.51BL/s、10.36±0.65 BL/s、8.88±0.50 BL/s、7.71±0.82 BL/s,如表1所示。通过分析可知,临界游泳速度随着试验鱼体长的增加而增加,相对临界游泳速度随着试验鱼体长的增加而降低,基本都呈线性关系,如图3和图4所示。
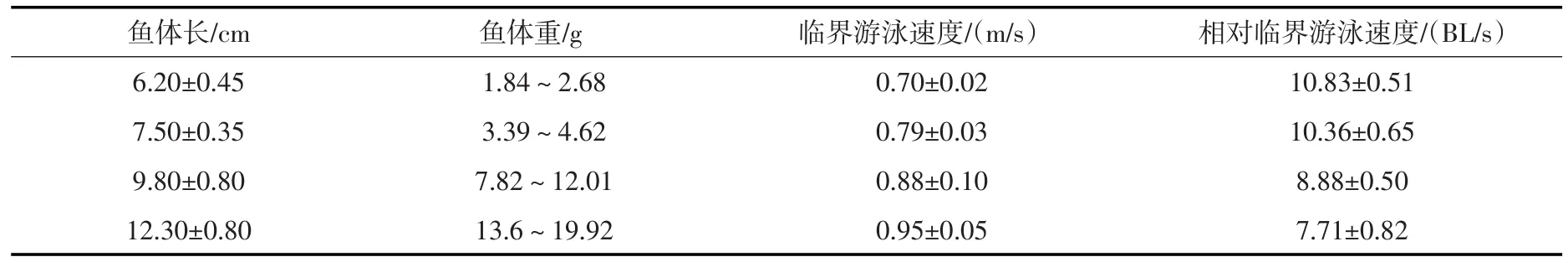
表1 草鱼幼鱼的临界游泳速度
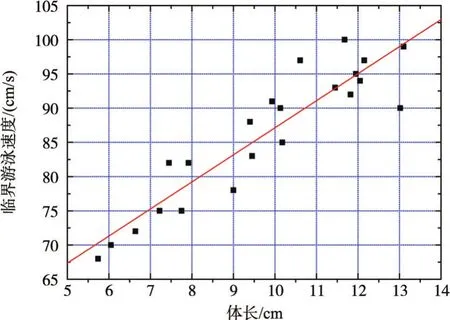
图3 草鱼幼鱼临界游泳速度与体长的关系
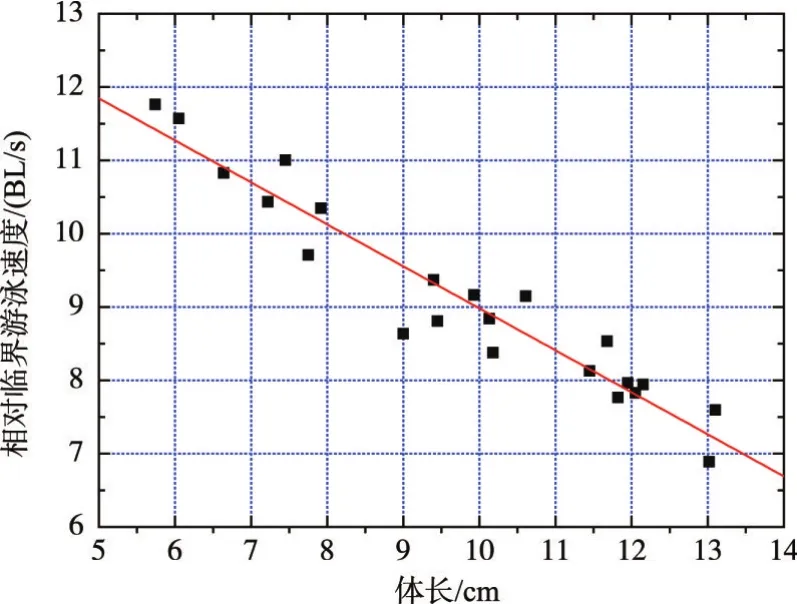
图4 草鱼幼鱼相对临界游泳速度与体长的关系
临界游泳速度与草鱼幼鱼体长的线性拟合关系式为:

相对临界游泳速度与草鱼幼鱼体长的线性拟合关系式为:
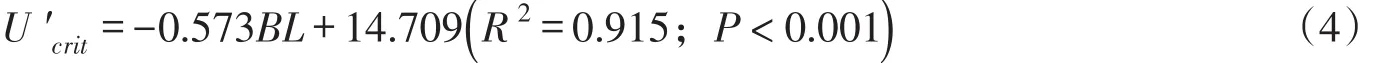
式中:Ucrit为临界游泳速度(m/s);U′crit为相对临界游泳速度(m/s);BL为试验鱼体长(cm)。
3.2草鱼幼鱼的游泳行为在自然界中,鱼类的游泳行为是一种状态较不稳定的运动,一般是阶段性的持续式游泳运动、暂停及偶发性的爆发游泳运动相互穿插发生[5]。在不同水流速度下,不仅鱼类的摆尾频率会发生变化,其游泳行为也不相同。鱼类在水体中一般呈现出4种不同的游泳状态,主要有逆流向前、逆流静止、逆流后退及顺流而下。
通过试验录像分析,对草鱼幼鱼在不同流速下的尾鳍摆动频率做了统计,从而得到试验鱼的摆尾频率与游泳速度的关系,如图5所示。草鱼幼鱼的游泳速度随着摆尾频率的增加而增加,基本呈线性关系,拟合关系式如式(4)所示:
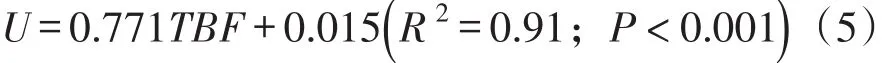
式中:U为游泳速度(m/s);TBF为试验鱼的摆尾频率(Hz)。
在本次试验中,通过对草鱼幼鱼游泳行为进行观察,发现4种体长的鱼类在整个试验过程中,随着试验流速的增大,其游泳行为大致可分为4个阶段:
第一阶段:在水体流速较小时,试验鱼在游泳区能自在的游动,试验鱼能够感应到水流方向而处于逆流状态,水流并未对鱼类游泳产生胁迫。此时,试验鱼的最大游速在0.30~0.45 m/s以下,约为试验鱼临界游速的45.6±2.3%,鱼类游泳基本上保持逆流状态,逆流静止所占时间比例为30%~57%左右,逆流向前保持在21%~42%左右;逆流后退占20%~30%,3种状态分布相对较为均匀,顺流而下所占比例为1%~3%。
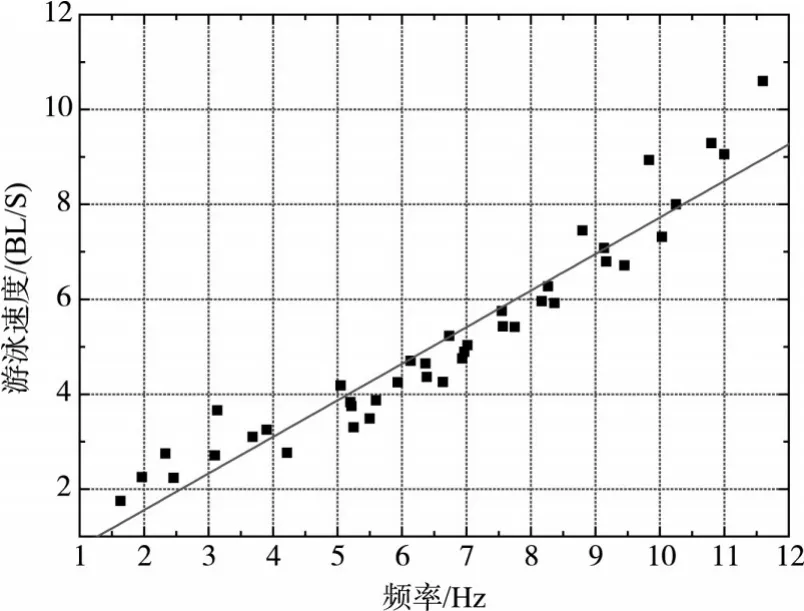
图5 草鱼幼鱼摆尾频率与游泳速度的关系
第二阶段:当水体流速达到一定值时,可明显观察到试验鱼因流速变大,摆尾频率增加,且鱼体肌肉处于紧张状态,此时,水流对鱼类的游泳产生了一定的阻碍作用。此时,鱼类最大游速在0.55~0.84 m/s以下,约为试验鱼临界游速的82.8±3.2%,主要以逆流静止状态为主,所占比例保持在40%~69%,较前一阶段有所增加,逆流向前占15%~33%,逆流后退占16%~34%。
第三阶段:当水体流速继续增大到一定阶段,试验鱼出现逆流后退的状态增多,并会有偶发性的逆流冲刺行为,随着流速的增大,这种反复逆流后退和冲刺的行为变得频繁。此时,鱼类的最大游速为0.64~0.97 m/s,约为试验鱼临界游速的95.9±1.8%,游泳状态仍以逆流静止为主,所占比例为27%~56%,逆流向前为5%~42%,逆流后退为16%~39%;
第四阶段:当水体流速增加到足够大时,试验鱼的尾鳍触及试验区下游拦网不再向前游动,此时流速已接近试验鱼的临界游速,试验鱼已接近疲劳状态。在该阶段,逆流静止和逆流后退所占比例相当,分别为27%~50%和32%~50%,逆流向前为16%~42%。
4种体长的草鱼幼鱼在上述4个阶段的最大游速和游泳状态所占时间比例如表2和图6所示。在这4个游泳阶段,顺流而下只出现在了第一阶段,其余三个阶段均只出现了逆流游泳的状态。随着流速的增加,逆流静止状态呈现先增加后减小的趋势,逆流前进所占时间比例变化不明显,而逆流后退则随着流速的增加有增加的趋势,尤其接近临界游速的第四阶段,逆流后退所占时间比例最高达50%。综合分析4个阶段的游泳行为,以逆流静止状态所占比例最高,在整个试验过程中约占50%左右。
4 结论
在28±1℃水温下,草鱼幼鱼(体长5~15 cm,1.84~19.92 g)的临界游速为0.84±0.16 m/s,且随鱼类体长增加而线性增长,其关系式为:Ucrit=0.04BL+0.476;在整个试验过程中,随着流速的增加,鱼类不断调整其游泳行为来适应水流环境,草鱼幼鱼逆流静止所占的时间比例呈先增后减的趋势,逆流后退则随着流速的增加有增加的趋势,尤其当流速接近临界游速时,逆流后退所占时间比例高达50%;根据试验鱼对水流流速的行为反应,其摆尾频率随着水流速度增加而增加,整个试验阶段可分成4个阶段,每个阶段的最大游速依次约为鱼类临界游速的45.6%、82.8%和95.9%和100%。本研究成果是对四大家鱼游泳能力和游泳行为研究的有益补充,可为确定鱼道池室的最大流速和休息池间距提供参考,对保护和恢复草鱼种群具有重要意义。
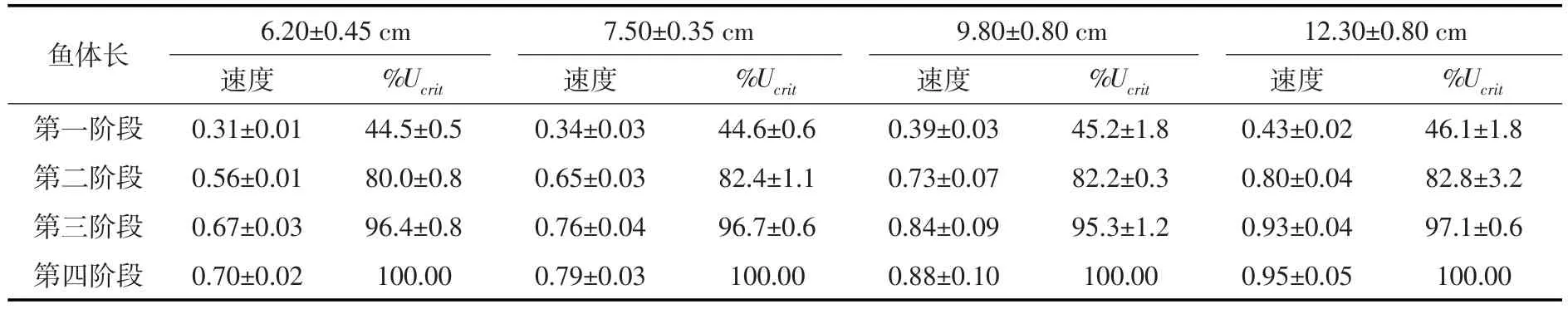
表2 草鱼幼鱼各游泳阶段的最大流速
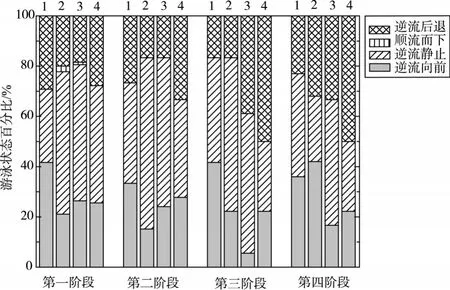
图6 4种体长的试验鱼在4个阶段中各游泳状态所占时间比例
[1] 中华人民共和国水利部,中华人民共和国国家统计局.第一次全国水利普查公报[M].北京:中国水利水电出版社,2012.
[2] 董哲仁,孙东亚,等.生态水利工程原理与技术[M].北京:中国水利水电出版社,2007.
[3] 陈凯麒,常仲农,曹晓红,等.我国鱼道的建设现状与展望[J].水利学报,2012,43(2):182-188.
[4] 石小涛,陈求稳,黄应平,等.鱼类通过鱼道内水流速度障碍能力的评估方法[J].生态学报,2011,31(22):6967-6972.
[5] 郑金秀,韩德举,胡望斌,等.与鱼道设计相关的鱼类游泳行为研究[J].水生态学杂志,2010,31(5):104-110.
[6] 刘乐和.长江葛洲坝水利枢纽兴建后对中、上游主要经济鱼类影响的综合评价[J].淡水渔业,1991,(3):3-7.
[7] 长江三峡工程生态与环境监测公报[Z].2013.
[8] 张安杰,曹振东,付世建.溶氧水平对草鱼幼鱼快速启动能力的影响[J].生态学杂志,2013,32(4)927-931.
[9] 房敏,蔡露,等.运动消耗对草鱼幼鱼游泳能力的影响[J].长江流域资源与环境,2014,23(6):816-820.
[10] 路波,刘伟,等.草鱼快速启动过程的加速-滑行游泳行为[J].水产学报,2014,38(6):829-834.
[11] 鲜雪梅,曹振东,付世建.4种幼鱼临界游泳速度和运动耐受时间的比较[J].重庆师范大学学报:自然科学版,2010,27(4):16-20.
[12] 胡茂林.鄱阳湖湖口水位、水环境特征分析及其对鱼类群落与洄游的影响[D].江西:南昌大学,2009.
[13] 石小涛,陈求稳,等.胭脂鱼幼鱼的临界游泳速度[J].水生生物学报,2012,36(1):133-136.
[14] Plaut I.Critical swimming speed:its ecological elevance[J].Comparative Biochemistry and Physiology,2001,131(A):41-50.
Experimental study on swimming capability and swimming behavior of juvenile grass carp
GONG Li,WU Yihong,BAI Yinbaoligao,MU Xiangpeng
(State Key Laboratory of Simulation and Regulation of Water Cycle in River Basin,China Institute of Water Resources and Hydropower Research,BeiJing 100038,China)
The swimming capability and swimming behavior of the juvenile grass carp(bl=5.0~15.0 cm)were tested using experiment apparatus by increasing velocity method at 28±1℃.The results show that the critical swimming speed of juvenile grass carp ranges from 0.68 to 1.0 m/s.There is a significant linear cor⁃relation between the critical swimming speed and fish body length,and the fitting function is described as Ucrit=0.04BL+0.476.According to the response of fish swimming behavior to the flow velocity,fish need to constantly adjust their behavior to adapt to the environment with the increasing of flow velocity.Their tail beat frequency(TBF)are roughly proportional to rising,and the entire testing process can be divided into four stages.The maximum swimming speed of each stage is about 45.6%,82.8%,95.9%and 100%of the critical swimming speed,respectively.The research results complement the swimming capability gaps of the four major chinese carps(FMCC),and provide an important reference for the hydraulic design of fish⁃way.
swimming capability;swimming behavior;juvenile grass carp;critical swimming speed;fish⁃way
:Adoi:10.13244/j.cnki.jiwhr.20150010
11672-3031(2015)03-0211-06
(责任编辑:韩 昆)
2015-01-04
中国水利水电科学研究院重点科研专项(水基本科研1443)
龚丽(1984-),女,湖南益阳人,博士生,主要从事水力学及河流动力学工作。E-mail:gongli1011@163.com
猜你喜欢
天天爱科学(2022年9期)2022-09-15 01:12:28
科普童话·百科探秘(2022年8期)2022-05-30 10:48:04
昆明医科大学学报(2022年1期)2022-02-28 07:46:32
基层中医药(2021年11期)2021-03-26 18:18:41
小学生作文(低年级适用)(2019年4期)2019-04-29 09:14:14
散文诗(2017年18期)2018-01-31 02:43:49
动物营养学报(2015年10期)2015-12-01 03:30:54
动物营养学报(2015年10期)2015-12-01 02:26:22
应用海洋学学报(2014年1期)2014-11-22 07:17:50
天然产物研究与开发(2014年8期)2014-04-27 14:16:29
