
斑马鱼uba6和uba7突变体的构建与鉴定
2015-05-11 13:56丁慧美黄红辉
中国比较医学杂志 2015年10期
丁慧美,阮 华,黄红辉
(淡水鱼类资源与生殖发育教育部重点实验室,三峡库区生态环境与生物资源省部共建国家重点实验室培育基地,西南大学生命科学学院,重庆 400715)
斑马鱼uba6和uba7突变体的构建与鉴定
丁慧美,阮 华,黄红辉
(淡水鱼类资源与生殖发育教育部重点实验室,三峡库区生态环境与生物资源省部共建国家重点实验室培育基地,西南大学生命科学学院,重庆 400715)
目的 构建斑马鱼uba6和uba7突变体。方法 运用TALEN技术在斑马鱼uba6和uba7基因编码区引进突变,并测序鉴定突变类型。结果 成功获得了两种突变类型的uba6和三种突变类型的uba7斑马鱼突变体。结论 成功构建了斑马鱼uba6和uba7突变体,为进一步研究斑马鱼uba6和uba7基因的功能提供了材料。
斑马鱼;uba6基因;uba7基因;TALEN
TALEN (transcription activator-like (TAL)effector nucleases)是一种新型的靶向基因编辑技术,现已应用于细胞、酵母、植物、斑马鱼及大、小鼠等各类研究对象。其原理是利用植物细菌Xanthomonas sp.TALE蛋白核酸结合域的氨基酸序列可对应识别结合位点的核酸,组装Xanthomonas sp.TALE的氨基酸序列模块识别特定DNA序列,并与核酸酶重组融合,这一人工构建的核酸酶可在特定位点识别(通过TALE氨基酸序列模块执行)并剪切(通过核酸酶执行)目的基因DNA序列,断裂的DNA在修复过程中会引进小片段插入或缺失,导致基因突变[7-8]。
斑马鱼是研究脊椎动物发育的理想模式动物,其基因组与人类高度同源。小鼠中UBA6和UBA7基因功能已有一定的研究[4,6],但在斑马鱼中这两个基因的功能尚不可知,运用反向遗传学手段在斑马鱼中建立突变体有助于研究其功能。本文应用TALEN技术对斑马鱼中uba6和uba7基因进行编辑,通过筛选获得可遗传的突变体,为后续进一步探究斑马鱼中uba6和uba7基因的功能提供了重要材料。
1 材料和方法
1.1 材料
1.1.1 实验动物
野生型斑马鱼品系AB Tüebingen品系。
1.1.2 质粒
另外,教师在实践中要引导学生自己摸索实验条件,想办法改进操作方法。例如,某小组探究“绿色蔬菜变黄过程中色素的变化”,预计提取、分离新鲜和放置变黄后的菠菜叶中的色素进行比较,但时值5月,菠菜放在冰箱,没变太黄前就腐烂了,教师要引导学生分析原因,摸索储存条件,或探寻其他的实验方法。某小组探究“红花檵木紫色叶片中的色素种类”设计了用清水纸层析分离色素的实验组。实验过程中发现,滤纸条吸水后变软变重,会贴壁或倒入水中。因此,不能像平时实验中那样把滤纸条靠在试管或小烧杯壁上,学生自主进行了改进:使用试管架和夹子把滤纸条固定并吊在液面上方,保证下端没入水中而不会整根滤纸条倒入水中。
TALEN载体为pCS2-FokⅠ,靶向uba6基因外显子5和uba7基因外显子2的TALEN质粒由北京唯尚立德生物科技有限公司合成。uba6靶位点:CTGGGCACCAACTTCT tcatcagagaggagg ATGTGAATA ATCAGA,其中左臂 TALE识别序列 CTGGGCA CCAACTTCT 16 bp,右臂TALE识别序列TCTGATT ATTCACAT 15 bp,间隔15 bp,突变体检测酶BseRI识别序列 GAGGAG。uba7靶位点:TGCCAAAAA CGTGATcctggctggagttagaACAGTTACAATTCAGG,其中左臂TALE识别序列TGCCAAAAA CGTGAT 15 bp,右臂TALE识别序列CCTGAATTGTAACTGT 16 bp,间隔16 bp,突变体检测酶Bpm I识别序列CTGGAG。
1.1.3 引物
uba6基因外显子5扩增引物:TTGCTGATATAG GTGACTTGGAG(正向),CACATTAACCTTTCAC CGGAAG(反向);uba7基因外显子2扩增引物:GGTAATTATTTTGGCACCAACAGC(正向),GGAGT TACAAATGCCTATGTGCC(反向)。
1.1.4 主要试剂
本文实验所用引物由上海生工生物技术公司以及上海Invitrogen公司合成。限制性内切酶购自NEB公司;AxyPrepTM plasimid Miniprep Kit和AxyPrepTM PCRCleanup Kit购自 Axygen公司;T4 DNA连接酶购自Roche公司;体外转录试剂购自Ambion公司。
1.2 方法
1.2.1 胚胎收集
在成年斑马鱼交配前一天将斑马鱼置于有适量养殖系统水的交配盒中,用挡板将雌雄斑马鱼分开。次日早晨,将挡板拔出,斑马鱼产卵,30 min后可收集,放置于28℃恒温培养,在胚胎12 hpf(hour post-fertilization)时加入0.03%PTU抑制色素形成。
1.2.2 全胚胎原位杂交
探针合成与全胚胎原位杂交实验依据已有报导进行[9]。本实验中所用uba6和uba7RNA反义探针浓度为1 ug/mL。
1.2.3 斑马鱼uba6和uba7突变体的构建
将体外转录的编码识别TALE左右臂序列蛋白和核酸酶融合蛋白的mRNA等量混合显微注射入一细胞期斑马鱼胚胎,注射浓度为500 pg/nL。在1.5 dpf(day post-fertilization)收集胚胎至EP管中,每管8个胚胎,加入80 uL 50 mmol/L NaOH溶液,95℃加热裂解10 min,冷却后加入 8 uL 1mol/L Tris8.0混合均匀,即为提取的基因组DNA。以此基因组DNA为模板进行PCR反应,扩增产物用特定的酶进行酶切检测。如果对照组胚胎的PCR产物能够全部被切开,而注射过mRNA的实验组胚胎的PCR产物中有未切开的PCR条带,则说明靶点发生了突变,TALEN质粒有效。将注射后的斑马鱼胚胎饲养至成鱼,为F0代。F0代与野生型交配产卵,收集胚胎,重复上述步骤提取基因组DNA、PCR扩增及酶切检测,确定此F0是否在生殖细胞水平携带突变,如携带突变,将F0与野生型生产的胚胎饲养至成鱼,为F1代。拔取F1成鱼鳞片,将鱼鳞片放入PCR管中,加入10 uL 50 mmol/L NaOH溶液,95℃加热裂解 10 min,冷却后加入 8 uL 1mol/L Tris8.0混合均匀,获得基因组DNA。以此基因组DNA为模板进行PCR反应,扩增产物纯化后送样测序,通过读取套峰获得斑马鱼F1突变类型。
2 结果
2.1 斑马鱼uba6基因的时空表达模式
检测基因时空表达模式有助于研究基因的功能。我们运用全胚胎原位杂交的方法检测了斑马鱼中uba6基因早期的表达模式。结果显示uba6呈现全身性表达,在1~3 dpf时于肠中有一定程度的富集,此富集随发育的进行趋于明显(彩插3图1)。
2.2 斑马鱼uba7基因的时空表达模式
类似于uba6基因,我们也检测了uba7基因的时空表达模式。结果显示,uba7基因在1~3 dpf的斑马鱼胚胎中表达,表达模式为全身性,在肠中稍有富集(彩插3图2)。
2.3 斑马鱼中uba6和uba7突变体建立
2.3.1 TALEN质粒活性检测
为检测TALEN靶点设计是否工作,必须检测质粒的活性。将以质粒为模板体外转录的mRNA注射至一细胞期的斑马鱼胚胎中,提取注射后胚胎的基因组DNA,PCR扩增包含靶点的DNA片段,酶切检测此片段能否被切开。结果如图3所示,对照组野生型胚胎的PCR产物能够完全被切开;而实验组即注射了mRNA的斑马鱼胚胎中部分PCR产物被切开,部分PCR产物未被切开,说明该靶点发生了突变,且效率较高。
2.3.2 F0突变体筛选
将上述实验中注射mRNA的斑马鱼饲养至成鱼,为F0代。F0是野生型和突变型细胞的嵌合体,为检测突变是否产生在生殖细胞水平,将F0代与野生型交配产生F1子代,提取F1胚胎的基因组DNA,采取2.3.1中相同的策略,即PCR扩增含靶位点的基因组DNA并酶切检测。结果如图3所示,对照组野生型的PCR产物能够完全被切开;而实验组PCR产物未能被全部切开,这说明部分F1子代含有靶位点突变,F0代含有可遗传的突变,将F1代斑马鱼胚胎饲养至成鱼。

图3 TALEN质粒活性检测及F0筛选Note:asterisk,PCR product;arrowhead,enzyme digest products.Fig.3 Activity tests of TALEN plasmids and F0 screen
2.3.3 F1突变体筛选
F1代斑马鱼中含有野生型和杂合突变体,为鉴定出杂合突变体,拔取少许F1成鱼鳞片,提取基因组DNA,扩增含有靶位点的基因组DNA片段,测序检测靶位点序列,测序结果显示F1代含有多种突变类型,表1和表2分别列举了F1代中筛选获得的移码突变类型,其中红色标记部分是TALEN设计敲除的靶位点序列,这些移码突变都可以产生早熟终止密码子。

表1 TALEN uba6 F1突变类型Tab.1 TALEN uba6 F1 mutation type
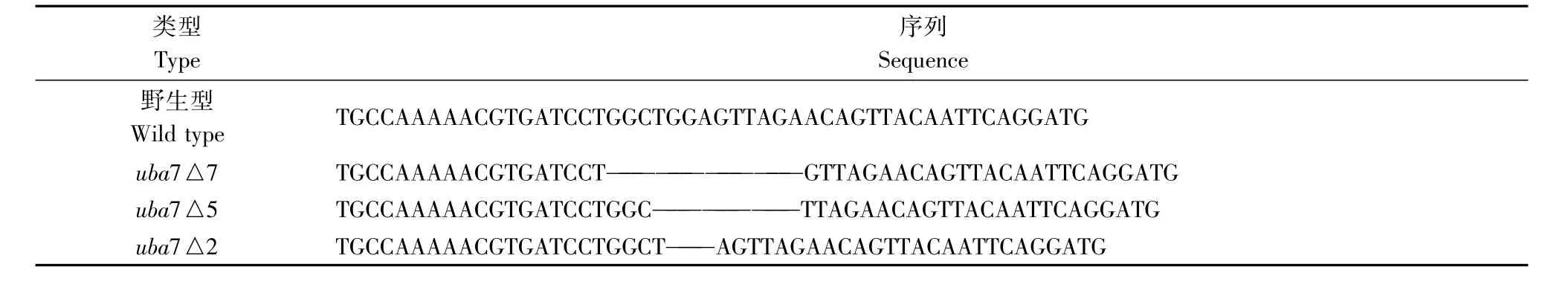
表2 TALEN uba7 F1突变类型Tab.2 TALEN uba7 F1 mutation type
3 讨论
小鼠中的研究表明,UBA6基因敲除会导致突变体胚胎在E10.5死亡[4],而UBA7基因的敲除虽然不影响小鼠的存活与生殖,但是会导致ISG15蛋白修饰的缺失[6]。斑马鱼中这两个基因的功能未有报道,我们构建的uba6和uba7突变体将是在斑马鱼中研究其功能的出发材料。基因的时空表达模式结果显示,两个基因在胚胎期都有表达,这提示着两个基因有可能调控斑马鱼的胚胎发育。尽管我们获得的突变体中基因突变会导致移码,但是它们有可能不会导致胚胎出现发育缺陷表型,可能的原因有以下几种(1)斑马鱼中有其他基因(例如可能存在的复制基因、功能相似的基因等)补偿了突变基因的功能;(2)斑马鱼uba6和uba7基因有可能存在多种异构体,突变并没有导致所以异构体移码突变;(3)两者不是斑马鱼胚胎发育必需的基因。
运用TALEN技术构建突变体,可以获得多种突变类型,通常都是小片段DNA的插入或缺失。我们的实验中也获得了多种突变类型,只保留了引起移码的突变,这些突变都导致了早熟终止密码子的出现,从理论上分析这些突变可以导致基因功能丧失,但是还是需要其他分子生物学方法的验证。
[1] Hershko A,Ciechanover A.The ubiquitin system[J].Annu Rev Biochem.1998,67:425-479.
[2] Pelzer C,Kassner I,Matentzoglu K,etal.UBE1L2,a novel E1 enzyme specific for ubiquitin[J].J Biol Chem.2007,282 (32):23010-23014.
[3] Jin J,Li X,Gygi SP,et al.Dual E1 activation systems for ubiquitin differentially regulate E2 enzyme charging [J].Nature.2007,447(7148):1135-1138.
[4] Chiu YH,Sun Q,Chen ZJ.E1-L2 activates both ubiquitin and FAT10[J].Mol Cell.2007,27(6):1014-1023.
[5] Yuan W,Krug RM.Influenza B virus NS1 protein inhibits conjugation of the interferon(IFN)-induced ubiquitin-like ISG15 protein[J].EMBO J.2001,20(3):362-371.
[6] Kim KI,Yan M,Malakhova O,et al.Ube1L and protein ISGylation are not essential for alpha/beta interferon signaling[J].Mol Cell Biol.2006,26(2):472-479.
[7] Miller JC,Tan S,Qiao G,et al.A TALE nuclease architecture for efficientgenome editing[J].Nat Biotechnol.2011,29:143–148.
[8] ZhongweiQiu,Meizhen Liu, Zhaohua Chen, et al. Highefficiency and heritable gene targeting in mouse by transcription activator-like effector nucleases[J].Nucleic Acids Res.2013,41(11):e120.
[9] Huang H,Ruan H,Aw M Y,et al.Mypt1-mediated spatial positioning of Bmp2-producing cells is essential for liver organogenesis[J].Development.2008,135(19):3209 -3218.
Construction and identification of zebrafish uba6 and uba7 mutants by TALEN
DING Hui-mei,RUAN Hua,HUANG Hong-hui
(Key Laboratory of Freshwater Fish Reproduction and Development,Ministry of Education,State Key Laboratory Breeding Base of Eco-Environments and Bio-Resources of the Three Gorges Reservoir Region,School of Life Sciences,Southwest University,Beibei,Chongqing 400715,China)
Objective To construct zebrafish uba6 and uba7mutants.Method In-delswere introduced into coding regions of zebrafish uba6 and uba7 gene by TALEN technology,andmutation typeswere identified by sequencing.Results We obtained two uba6 and three uba7mutantswith different small fragment deletions resulting in reading frame shifts.Conclusion These uba6 and uba7mutants provided startingmaterials for the study of zebrafish uba6 and uba7 gene functions.
Zebrafish;uba6;uba7;TALEN
R-332
A
1671-7856(2015﹞10-0007-04
10.3969.j.issn.1671.7856.2015.010.002
国家自然科学基金面上项目(31071286);西南大学中央高校基本科研业务费(XDJK2014A013;2362014xk02)。
丁慧美(1990-),女,硕士,研究方向:发育生物学E-mail:15895840307@163.com。
黄红辉(1974-),男,博士,教授,研究方向:发育生物学E-mail:honghuih@126.com;阮华(1974-),女,博士,教授,研究方向:发育生物学,E-mail:ruanhua23@126.com。
﹞2015-08-18
研究报告
猜你喜欢
亚热带农业研究(2022年1期)2022-08-08
四川农业大学学报(2021年6期)2022-01-05
小天使·二年级语数英综合(2021年8期)2021-08-16
农业科技通讯(2021年1期)2021-03-06
家畜生态学报(2020年6期)2020-07-02
中国农业科技导报(2020年3期)2020-03-15
中国食品学报(2019年12期)2019-01-13
中成药(2017年6期)2017-06-13
第二课堂(小学版)(2015年2期)2015-05-22
中国药业(2014年21期)2014-05-26
