
白花泡桐二倍体及其同源四倍体的AFLP 和MSAP分析
2015-05-10 02:07:44翟晓巧张晓申赵振利范国强
河南农业大学学报 2015年2期
翟晓巧,张晓申,赵振利,范国强
(1.河南省林业科学研究院,河南 郑州450008;2.河南农业大学泡桐研究所,河南郑州450002)
自然界中大约70%的被子植物在进化史中经历过1次或1次以上多倍化事件[1]。植物发生多倍体过程中,一方面在染色体结构和基因表达模式发生了变化[2-6],另一方面在DNA甲基化水平上也发生了变化[7~17]。研究表明,多倍体植物具有器官和生物量明显增大的特征和较强适应环境条件的能力[18,19]。泡桐(Paulownia spp.)是中国重要的速生用材和平原绿化树种之一,因其独特的生物学和生态学特性,能与农作物间作形成独特的生态复合系统,在防风固沙、改善生态环境、保障农作物稳产高产和提高农民生活水平等方面发挥着重要作用,但现有品种不能完全满足国民经济发展的需求。近年来,科技工作者创制的多个四倍体泡桐新种质[20-25],一方面丰富了泡桐的种质资源,另一方面也为筛选泡桐新品种提供了材料支撑。目前,虽然开展过四倍体泡桐叶片结构、光合作用和木材特性以及耐干旱胁迫等方面的研究工作[26,27],但迄今国内外还未见四倍体泡桐遗传特性相关文章的报道。因此,本研究利用AFLP和MSAP分子标记技术,研究了二倍体白花泡桐及其同源四倍体的遗传差异及DNA甲基化的变化,以期为阐明四倍体泡桐优良特性的分子机理奠定基础。
1 材料与方法
1.1 试验材料
材料为河南农业大学泡桐研究所林木生物技术实验室经体细胞胚胎发生途径获得、培养30 d的白花泡桐(Paulownia fortunei)二倍体及其同源四倍体组织培养苗。剪取适量生长状况良好、大小一致幼苗长约0.5 cm的顶芽用液氮速冻后放于-86℃冰箱内备用。组培苗培养条件为温度(25±2)℃、光照强度 130 μmol·m-2·s-1、光照时间 16 h·d-1。
1.2 试验方法
1.2.1 泡桐DNA的提取 白花泡桐二倍体及其同源四倍体DNA的提取参照张延召等的[28]方法。
1.2.2 泡桐AFLP和MSAP分析 白花泡桐二倍体及其同源四倍体DNA的AFLP和MSAP扩增所需96对引物组合的碱基序列和扩增过程序及其产物的电泳分别参照曹喜兵等的[29,30]方法。
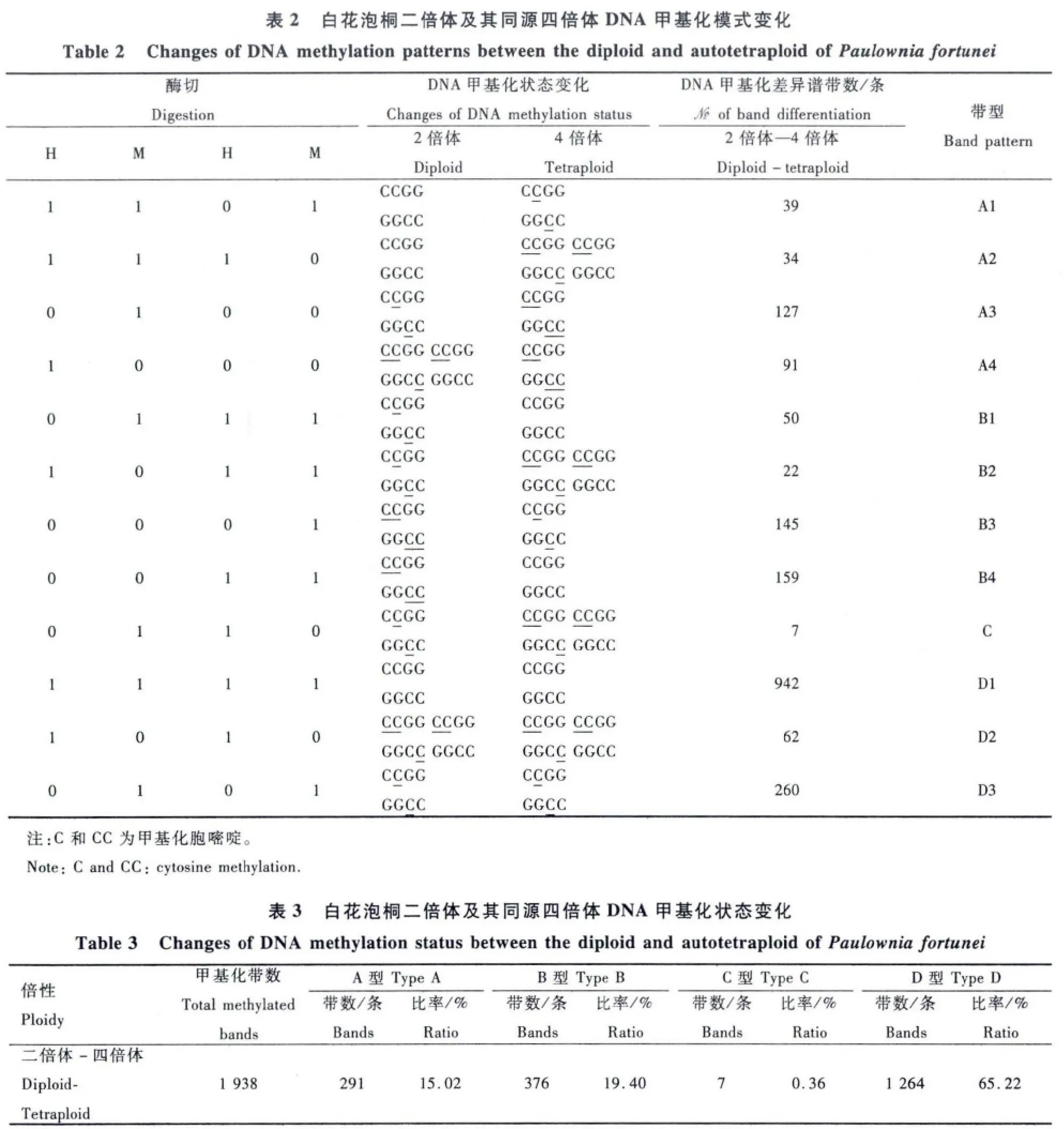
1.2.3 电泳凝胶的谱带分析 AFLP和MSAP扩增产物电泳结束、凝胶显影后,先在UMAX Power-Look 2100XL型扫描仪扫描成像,再分别统计分析不同MMS浓度处理样品间AFLP和MSAP凝胶谱带的变化。对MSAP凝胶谱带来说,H和M分别为EcoRⅠ/HpaⅡ和EcoRⅠ/MspⅠ双酶切产物电泳谱带。每1条谱带代表1个酶切位点。将谱带有或无分别记作1或0。将每个DNA样品的H和M扩增谱带可划分为4种[种类I(H,M=1,1),无甲基化发生;种类Ⅱ(H,M=1,0),单链 DNA外甲基化;种类Ⅲ(H,M=0,1),双链 DNA内甲基化;种类Ⅳ(H,M=0,0),双链DNA外甲基化]。DNA甲基化类型分为多态性和单态性类型。DNA多态性类型包括:A型(甲基化)、B型(去甲基化)和C型(不定)型。其中,A型中的A1和A2代表DNA重新甲基化(对照样H和M泳道均有带,而处理样仅H或M泳道有带),A3和A4代表DNA超甲基化(对照样仅H或M有一条带,而处理样H和M泳道都没带)。B型(Bl、B2、B3和 B4)代表DNA去甲基化,DNA甲基化谱带与A型相反。C型代表DNA甲基化的不确定性(二倍体与四倍体泡桐DNA甲基化差异谱带无法确定)。单态性类型为D型(对照样与处理样间DNA谱带相同)。同时,计算样品的总DNA甲基化水平[(种类Ⅱ+种类Ⅲ)/(种类Ⅰ+种类Ⅱ+种类Ⅲ)×100%]和总DNA甲基化多态性[(A型+B型+C型)/(A型+B型+C型+D型)×100%]及 DNA单态性[D型/(A型+B型+C型+D型)×100%]。
2 结果与分析
2.1 白花泡桐二倍体及其同源四倍体DNA碱基序列变化
由白花泡桐二倍体及其同源四倍体DNA的AFLP扩增结果(图1)可以看出,白花泡桐二倍体及其同源四倍体的DNA利用同一引物均可在相同的位置扩增出相同的条带,也就是说白花泡桐二倍体及其同源四倍体DNA的AFLP酶切位点没有发生变化。AFLP每对引物平均可以扩增出50~70条谱带。虽然白花泡桐二倍体及其同源四倍体的DNA经不同引物AFLP扩增后,产生片段的大小和数量存在一定的差异,该结果说明,白花泡桐二倍体染色体加倍前后DNA碱基序列没有发生变化,但同源四倍体白花泡桐DNA的高级结构可能发生了变化。
2.2 白花泡桐二倍体及其同源四倍体总DNA甲基化变化
2.2.1 白花泡桐二倍体及其同源四倍体总DNA甲基化水平变化 白花泡桐二倍体及其同源四倍体的MSAP凝胶电泳谱带统计结果(表1)表明,白花泡桐二倍体染色体加倍前后DNA甲基化水平发生了一定变化。在MSAP扩增产物的电泳凝胶上,白花泡桐二倍体扩增出的位点达到2 357个,其中全甲基化位点为283个(占总扩增位点的12.01%)、半甲基化位点为564个(占总扩增位点的23.93%)、总甲基化位点为847个(占总扩增带数的35.94%);白花泡桐同源四倍体扩增出2 512个位点,其中全甲基化位点数为325个(占总扩增带数的12.94%)、半甲基化位点为629个(占总扩增位点的25.04%)、总甲基化位点为954个(占总扩增带数的37.98%)。白花泡桐二倍体及其同源四倍体CCGG位点均以双链甲基化为主,白花泡桐同源四倍体全甲基化水平和半甲基化水平高于其二倍体。该结果说明,白花泡桐二倍体染色体加倍形成的同源四倍体的DNA发生了甲基化修饰,这也可能是同源四倍体白花泡桐与其二倍体相比形态等方面未呈现倍增的重要原因之一。

图1 白花泡桐二倍体及其同源四倍体的AFLP扩增产物部分SDS-PAGEFig.1 SDS-PAGE of partial AFLP amplification products of diploid and tetraploid of Paulownia fortunei

2.2.2 白花泡桐二倍体及其同源四倍体DNA甲基化模式的变化 由白花泡桐二倍体及其同源四倍体MSAP扩增产物的凝胶电泳图谱(图2)及其统计结果(表2和表3)可以看出,白花泡桐四倍体DNA甲基化模式不同于其二倍体。与二倍体比较,白花泡桐四倍体DNA甲基化(A型)位点数和去甲基化(B型)位点数分别为291个(占总甲基化多态性位点数的15.02%)和376个(占总甲基化多态性位点数的19.40%)。总DNA甲基化多态性为34.78%、单态性(D型)为65.22% 。这说明白花泡桐同源四倍体的DNA甲基化模式变化频率小于其二倍体。

图2 白花泡桐二倍体及其同源四倍体的DNA甲基化模式变化Fig.2 DNA methylation patterns of the diploid and tetraploid Paulownia fortunei
3 讨论
植物同源四倍体是其二倍体染色体加倍后形成的新种质。从理论上讲,同源四倍体的形态等特征应呈现其二倍体的倍增结果,但实际上并非如此[26,31-34]。有人认为,造成该结果的原因可能与植物二倍体染色体加倍过程中相关基因发生突变有关[35,36]。本试验中,白花泡桐染色体加倍前后DNA碱基序列没有发生变化,这在其它研究中也得出相同的结果[37-43]。因此,同源四倍体植物DNA碱基序列变化与否不是形态等特征未呈现其二倍体倍增结果的主要原因。研究表明,虽然在AFLP水平上没有检测到二倍体白花泡桐染色体加倍形成的同源四倍体DNA碱基序列的变化,但四倍体白花泡桐DNA甲基化水平和模式都发生了变化,并且同源四倍体白花泡桐的DNA甲基化模式变化频率小于其二倍体。众所周知,DNA甲基化可在不改变生物基因序列的情况下影响染色体的结构和基因的表达水平[44,45]。DNA甲基化常与基因表达的沉默有关,而DNA去甲基化则与启动基因的重新表达有关。本研究表明,同源四倍体白花泡的DNA甲基化水平高于其二倍体,四倍体白花泡桐DNA甲基化模式变化频率小于其二倍体。所以,同源四倍体白花泡桐DNA甲基化变化可能是形态等特征应未呈现其二倍体倍增的主要原因之一。该结论在西瓜、鸭梨、弯叶画眉草和百喜草的研究结果中[14-16,46]得到了验证。
[1] WENDEL J F.Genome evolution in polyploids[J].Plant Mol Biol,2000,42:225 -249.
[2] SWAPNA M,SIVARAJU K,SHARMA R K,et al.Single-strand conformationalpolymorphism ofESTSSRs:a potential tool for diversity analysis and varietal identification in sugarcane[J].Plant Mol Biol Rep.2011,29:505 -513.
[3] SONG K,LU P,TANG K,et al.Rapid genome change in synthetic polyploids of Brassica and its implications for polyploid evolution[J].Proc Natl Acad Sci,1995,92:7719-7723.
[4] PIRES J C,ZHAO J W,SCHRANZ M E,et al.Flowering time divergence and genomic rearrangements in resynthesized polyploids(Brassica)[J].Biol J Linn Soc Lond,2004,82:675 -688.
[5] LU B B,PAN X Z,ZHANG L,et al.A genome-wide comparison of genes responsive to autopolyploidy in Isatis indigotica using arabidopsis thaliana affymetrix genechips[J].Plant Mol Biol Rep,2006,24:197 -204.
[6] ADAMS K L,CRONN R,PERCIFIELD R,et al.Genes duplicated by polyploidy show unequal contributions to thetranscriptome and organspecific reciprocal silencing[J].ProcNatlAcad Sci,2003,100:4649-4654.
[7] KASHKUSH K,FELDMAN M,LEVY A A.Gene loss,silencing and activation in a newly synthesized wheat allotetraploid[J].Genetics,2002,160:1651 -1659.
[8] LIU B,WENDEL J F.Epigenetic phenomena and the evolution of plant allopolyploids[J].Mol Phylogenet Evol,2003,29:365 -379.
[9] WANG J,TIAN L,MADLUNG A,et al.Stochastic and epigenetic changes of gene expession in Arabidopsis polyploids[J].Genetics,2004,167:1961 -1973.
[10] HAN F P,FEDAK G,OUELLET T,et al.Rapid genomic changes in interspecific and intergenetic hybrids and allopolyploids of Triticeae[J].Genome,2003,46:716-723.
[11] MADLUNG A,MASUELLI R W,WATSON B,et al.Remodeling of DNA methylation and phenotypic and transcriptional changes in synthetic Arabidopsis allotetraploids[J].Plant Physiol,2002,129:733 -746.
[12] ZHANG H Y,PENG H,LI Y,et al.Analysis of DNA cytosine methylation between haploid and its corresponding diploid rice[J].Chin Sci Bull,2006,51:1529 -1535.
[13] CHEN L,CHEN J.Changes of cytosine methylation induced by wide hybridization and allopolyploidy in Cucumis[J].Genome,2008,51:789 -799.
[14] OCHOGAVIA A,CERVIGNI G,SELVA J P ,et al.Variation in cytosine methylation patternsduring ploidy level conversions in Eragrostis curvula[J].Plant Mol Biol,2009,70:17 -29.
[15] RODRIGUZE M P,CERVIGNI G D L,QUARIN C L,et al.Frequencies and variation in cytosine methylation patterns in diploid and tetraploid cytotypes of Paspalum notatum.Biologia[J].Plantarum,2012,56(2):276-282.
[16]胡宝全,王春国,方成泉,等.不同倍性鸭梨基因组DNA甲基化水平与模式分析[J].南开大学学报:自然科学版,2011,44(2):32-37.
[17] BAO A L,HU Q,XUE Z Y,et al.DNA methylation in genomes of several annual herbaceous and woody perennial plants of varying ploidy as detected by MSAP[J].Plant Mol Biol Rep,2011,29:784 -793.
[18]HILU K W.Polyploidy and the evolution of domesticated plants[J].Am J Bot,1993,80:1494 - 1499.
[19] LIU B,WENDEL J F.Non-mendelian phenomena in allopolyploid genome evolution[J].Curr Genomics,2002,3:489 -505.
[20]范国强,曹艳春,赵振利,等.白花泡桐同源四倍体的诱导[J].林业科学,2007,43(4):30 -36.
[21]范国强,杨志清,曹艳春,等.毛泡桐同源四倍体的诱导[J].植物生理学通讯,2007,43(1):109-111.
[22]范国强,魏真真,杨志清.南方泡桐同源四倍体的诱导及其体外植株再生研究[J].西北农林科技大学学报:自然科学版:2009,37(10):83-90.
[23]范国强,杨志清,曹艳春,等.秋水仙素诱导兰考泡桐同源四倍体[J].核农学报,2006,20(6):473 -476.
[24]范国强,翟晓巧,魏真真,等.豫杂一号泡桐体细胞同源四倍体诱导及其体外植株再生[J].东北林业大学报,2010,38(12):22-26.
[25]赵振利,何 佳,赵晓改,等.泡桐9501体外植株再生体系的建立及体细胞同源四倍体诱导[J].河南农业大学学报,2011,45(1):59-65.
[26]张晓申,翟晓巧,范国强,等.四倍体泡桐叶片显微结构观察及其抗逆性分析[J].河南农业大学学报,2012,46(6):646-650.
[27]翟晓巧,张晓申,赵振利,等.四倍体白花泡桐木材的物理特性研究[J].河南农业大学学报,2012,46(6):651-657.
[28]张延召,曹喜兵,翟晓巧,等.适用于AFLP分析的泡桐DNA提取方法研究[J].河南农业大学学报,2009,43(6):610-614.
[29]曹喜兵,何 佳,翟晓巧,等.泡桐AFLP反应体系的建立及引物筛选[J].河南农业大学学报,2010,44(2):145-150.
[30]曹喜兵,赵改丽,范国强.泡桐MSAP反应体系的建立[J].河南农业大学学报,2012,46(5):536 -541.
[31]雷 春,陈劲枫,张晓青,等.不同倍性黄瓜的形态和一些生理生化指标比较[J].植物生理学通讯,2005,41(4):471-474.
[32]朱必才,高立荣.同源四倍体荞麦的研究[J].遗传,1988,10(6):6 -8.
[33]杨鹏鸣,黄群策,秦广雍,等.同源四倍体水稻若干性状的初步研究[J].西南农业学报,2011,24(2):396-399.
[34]刘庆忠,刘鹏,赵红军,等.同源四倍体皇家嘎啦苹果的生物学及光合生理特性研究[J].中国农业科学,2002,35(12):1573-1578.
[35]刘文革,王 鸣,阎志红.西瓜二倍体及同源多倍体遗传差异的AFLP分析[J].果树学报,2004,21(1):46-49.
[36]王卓伟,余茂德,鲁 成.桑树二倍体及人工诱导的同源四倍体遗传差异的AFLP分析[J].植物学通报,2002,19(2):194-200.
[37]焦 锋,楼程富,张有做,等.桑树变异株系基因组DNA扩增多态性RAPD研究[J].蚕业科学,2001,27(3):165-169.
[38]林 强,邱长玉,朱方容,等.桑树二倍体及其人工诱导同源四倍体遗传差异的RAPD分析[J].南方农业学报,2011,42(1):11-15.
[39]栾 丽,孔繁伦,何 涛,等.用SSR标记检测同源四倍体与二倍体水稻的遗传差异[J].应用与环境生物学报,2004,10(5):556-558.
[40]聂丽娟,王子成,王一帆,等.二倍体和四倍体西瓜的DNA甲基化差异分析[J].核农学报,2009,23(1):80-84.
[41]赵卫国,苗雪霞,黄勇平.桑树二倍体及其同源四倍体遗传差异的 ISSR分析[J].蚕业科学,2005,31(4):393-397.
[42] SONG K M,OSBORN T C.A method for examining expression of homologous genes in plant polyploid[J].Plant Mole cular Biology,1994,26:1065 -1071.
[43] STUPAR R M,BHASKAR P B,YANDELL B S,et al.Phenotypic and transcriptomic changes associated with Potato autopoly ploidization[J].Genetics,2007,176:2055-2067.
[44] MARTELOTTO L G,ORTIZ J P A,STEIN J,et al.A comprehensive analysis of gene expression alterations in a newly synthesized Paspalum notatum autotetraploid[J].Plant Sci,2005,169:211 -220.
[45] MARTELOTTO L G,ORTIZ J P A,STEIN J,et al.Genome rearrangements derived from autopolyploidization in Paspalum sp[J].Plant Sci,2007,172:970 -977.
[46]王春国,古 瑜,陈成彬,等.不同倍性西瓜基因组DNA甲基化水平与模式的MSAP[J].分子细胞生物学报,2009,42(2):118-125.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12 07:09:08
现代园艺(2017年13期)2018-01-19 02:28:17
江苏农业科学(2017年16期)2017-10-27 12:48:11
现代农业科技(2017年8期)2017-06-10 09:32:18
语文教学与研究(读写天地)(2016年9期)2016-12-17 08:30:24
中成药(2016年4期)2016-05-17 06:07:54
安徽林业科技(2015年6期)2015-09-25 10:46:38
中国果业信息(2015年6期)2015-01-23 17:38:07
吐鲁番(2014年2期)2014-02-28 16:54:49
