
细菌素Lcn972A产生菌的鉴定及其基因的克隆与序列分析
2015-05-05 11:59:46甘祥武叶志伟郭心悦郭丽琼林俊芳王艳婷廖腾达
食品工业科技 2015年21期
甘祥武,叶志伟,郭心悦,郭丽琼,林俊芳,王艳婷,廖腾达
(华南农业大学食品生物技术研究所,华南农业大学食品学院生物工程系,广东广州 510640)
细菌素Lcn972A产生菌的鉴定及其基因的克隆与序列分析
甘祥武,叶志伟,郭心悦,郭丽琼,林俊芳*,王艳婷,廖腾达
(华南农业大学食品生物技术研究所,华南农业大学食品学院生物工程系,广东广州 510640)
目的:从杏鲍菇子实体与海带中分离的63株乳酸菌中鉴定出产细菌素Lcn972A的菌株,并对细菌素编码基因进行克隆与基因序列分析。方法:采用生物信息学手段对已报道的细菌素Lcn972A编码基因进行分析比较,设计引物,分别以各乳酸菌基因组DNA为模板,利用PCR克隆技术克隆出细菌素Lcn972A的基因,利用生物信息学工具对其核酸序列和预测的蛋白序列进行分析。结果:在63株乳酸菌的基因组样品中有15组被克隆出目的条带,其中杏鲍菇乳酸菌A15与海带乳酸菌CX2样品PCR产物条带清晰稳定性好,经鉴定两样品的乳酸菌株为乳酸乳球菌(Lactococcuslactis)。对克隆的细菌素Lcn972A基因测序结果表明该基因序列全长276 bp。生物信息学分析显示该基因编码91个氨基酸,分子量大小22843.3 u,等电点5.33,氨基酸序列为亲水性,二级结构主要由α-螺旋、无规卷曲和延伸链组成。结论:从分离自杏鲍菇子实体乳酸乳球菌A15与海带乳酸乳球菌株CX2的细菌素编码基因及氨基酸序列与报道的有所不同,可能是新的细菌素,该细菌素的抗菌特点有待进一步的研究。
乳酸乳球菌,Lcn972A基因,分子鉴定,序列分析
细菌素(Bacteriocin)是由某些细菌在代谢过程中合成并分泌到环境中的具有杀灭或抑制与之相同或相似生境的其他微生物特性的杀菌蛋白或多肽物质[1]。Klaenhammer和Nes等根据氨基酸的一级结构、分子量的大小、热稳定性和抑菌谱等方面的不同,把细菌素分为硫醚抗生素、不包含羊毛硫氨酸的小分子多肽类细菌素、蛋白质类细菌素以及复合型细菌素等4类[2-3]。细菌素 Lcn972是一种非羊毛硫氨酸的小分子多肽类细菌素,它能够通过与细胞壁前体中的LipidⅡ特异性结合,影响隔膜形成,阻碍细胞分裂[4]。大部分乳酸菌细菌素对热稳定、具有高等电点和亲水特性,可以与食品一起进行加热处理[5]。目前对细菌素产生菌的研究主要集中在乳酸菌上[6]。国外对乳酸菌细菌素的研究较多,已经从鲑鱼[7]、伊朗乳制品[8]、泰国红树林[9]等物种乳酸菌的产物中分离到上百种细菌素。产细菌素的乳酸菌种类包括肉杆菌属、乳杆菌属、乳球菌属、明片株菌属、片球菌属等[10-11]。传统的筛菌方法耗时、步骤繁琐,且很难迅速筛选到细菌素。随着大量不同乳酸菌菌株基因组测序的完成,利用生物信息学的方法预测、分析和寻找新型细菌素成为可能[12-14]。由于细菌素的基因序列保守,在已知的细菌素DNA序列的基础上设计保守的PCR引物或制备寡核苷酸探针,通过采用生物信息学和分子生物学等手段能够对细菌素进行深入研究。
本文通过设计保守的PCR引物,从63株来源于杏鲍菇与海带的乳酸菌中鉴定细菌素,从杏鲍菇分离乳酸菌A15与海带分离乳酸菌CX2中克隆出相同Lcn972细菌素家族基因,与NCBI报道Lcn972细菌素在分子水平上存在差异,将其命名为Lcn972A,乳酸菌A15、CX2鉴定为乳酸乳球菌,对该基因与其推导的蛋白序列进行分析,为开发利用Lcn972A细菌素及其产生菌奠定基础。
1 材料与方法
1.1 材料与仪器
63株乳酸菌菌株 广州天河区市场杏鲍菇(Pleurotuseryngii)的子实体与海带(ThallusLaminariae)中分离并保存于华南农业大学食品生物技术研究所。
普通琼脂糖凝胶DNA纯化回收试剂盒、质粒小提试剂盒 TIANGEN公司;PCR产物回收试剂盒、pMD18-T克隆载体、Ex Taq DNA聚合酶 TAKARA公司;Taq DNA聚合酶 北京博迈德生物试剂公司;其他普通试剂均为国产分析纯;引物 上海捷瑞公司合成,DNA序列测序 上海美吉生物医药科技有限公司与北京六合华大基因科技股份有限公司。
乳酸菌MRS培养基[15]:蛋白胨10 g,牛肉膏8 g,酵母膏粉4 g,葡萄糖20 g,磷酸氢二钾2 g,柠檬酸三氨2 g,乙酸钠5 g,硫酸镁0.2 g,硫酸锰0.05 g,吐温80 1 g,pH(6.2±0.2),配制1 L,固体培养基加入1.2%琼脂粉。
大肠杆菌LB培养基[16]:胰蛋白胨10 g,酵母提取粉5 g,氯化钠10 g,配制1 L,固体培养基加入1.5%琼脂粉。
1.2 实验方法
1.2.1 溴钾酚紫指示剂法筛选乳酸菌 根据乳酸菌产酸特性,将样品预处理后,先进行梯度稀释,找到合适梯度后,在加有溴钾酚绿指示剂的MRS培养基上涂布,挑起使培养基变黄色的单菌落连续划线确定为纯种菌后,经革兰氏染色确定为纯种革兰氏阳性菌,将单菌落保存。
1.2.2 细菌素Lcn972A基因的引物设计 根据NCBI中检索到的以及文献报导的各Lcn972细菌素基因序列,通过生物信息学比对获得细菌素的保守序列,根据保守序列设计PCR扩增引物(上游引物LCB972 F:5′-GGATCCATGAAAACCAAGTCTCTCG-3′,下游引物LCB972 R:5′-GAGCTCTTACCAAAAGT-3′)并合成。利用溶菌酶法提取到的63株乳酸菌基因组DNA为模板[17],以LCB972 F/LCB972 R为引物进行PCR扩增。PCR的反应体系为:10×PCR Buffer(Mg2+plus)5.0 μL,dNTP Mix(2.5 mmol/L each)4 μL,引物LCB972-F(10 mol/L)和LCB972-R(10 mol/L)各1 μL,Ex-Taq(5 U/L)0.25 μL,基因组DNA 1 μL,ddH2O 37.75 μL 反应程序为:94 ℃预变性5 min;94 ℃变性30 s,37.7 ℃退火30 s,72 ℃延伸30 s,共35个循环;72 ℃延伸10 min;4 ℃保存PCR产物。PCR产物由上海美吉生物医药科技有限公司进行序列测定。
1.2.3 产细菌素乳酸菌株的分子鉴定 对能够扩增出目标条带的样品的菌株进行16S rDNA分子鉴定,16S rDNA 扩增引物采用通用引物[18],正向引物为27f:5′-AGAGTTTGATCCTGGCTCAG-3′;反向引物1495r:5′-CTACGGCTACCTTGTTACGA-3′。
1.2.4 Lcn972A基因序列分析 测序结果与峰图正常,接着使用NCBI网站提供的BLAST工具完成DNA序列及其推导的氨基酸序列的比对,使用在线工具Swissprot(http://www. expasy. ch/sprot/)分析Lcn972A基因编码氨基酸序列的理化性质、功能位点、信号肽、亲疏水性二级结构和三级结构。
2 结果与分析
2.1 乳酸菌的筛选与鉴定
用生理盐水将样品处理液稀释成10-1~10-6梯度,在10-6梯度下菌落利于观察,将不同类型单菌落在溴钾酚紫MRS培养基培养2~3 d,选取明显使周围培养基由紫转为黄色的单菌落,继续在加入溴钾酚紫的MRS培养基中划线培养,如图1所示,所得单菌落编号后进行革兰氏染色,将镜检呈紫色的革兰氏阳性菌编号保存,如图2所示。
图1 乳酸菌平板筛选Fig.1 Plate screening of lactic acid bacteria
图2 乳酸菌革兰氏染色镜检Fig.2 Gram staining microscopic examination of lactic acid bacteria
2.2 乳酸菌Lcn972A基因的克隆
以乳酸菌基因组DNA为模板,以Lcn972-F/Lcn972-R为引物,使用PCR法扩增Lcn972A基因,产物经凝胶电泳检测,在63株样品中,有15株样品含有与预期大小相一致的约300 bp的条带(图3),使用PCR产物纯化试剂盒将该目的片段回收并送样测序,测序结果显示15株样品中A15、CX2含有相同细菌素编码基因表达框,该细菌素基因全长为276 bp,具有完整的编码框。BLAST比对显示所得乳酸乳球菌基因与已经在NCBI报道的Lcn972基因(GenBank:AJ002203.2)的核苷酸序列的相似性为97.46%,推测氨基酸序列的相似性97.80%,将其命名为Lcn972A。
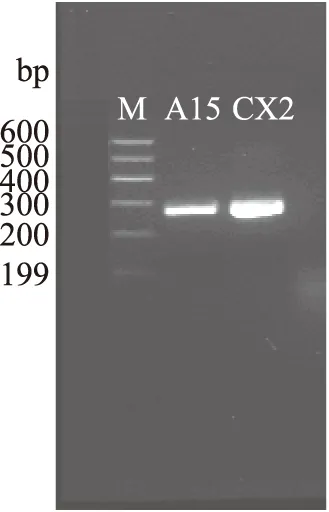
图3 Lcn972A基因PCR克隆电泳图Fig.3 The PCR clone of the Lcn972A gene注:泳道1为杏鲍菇乳酸菌A15,泳道2为海带乳酸菌CX2,图4同。
2.3 产细菌素乳酸菌的分子鉴定
经测序鉴定15株乳酸菌样品中A15、CX2含有细菌素基因,对克隆出Lcn972A基因的乳酸菌进行16S rDNA分子鉴定(图4),在DNA MakerⅢ约1400 bp处出现荧光条带。PCR产物测序后,将所得测序结果进行BLAST比对,结果显示待测乳酸菌与乳酸乳球菌(Lactococcuslactis)同源性达99%,推测其为乳酸乳球菌。
2.4 乳酸乳球菌Lcn972A基因的生物信息学分析
Lcn972A基因编码91个氨基酸残基,使用ExPASy网站的上的ProtParam分析软件[19]计算其相对分子质量大小为22843.3 u,理论等电点为5.33,蛋白质不稳定指数为50.52,属于不稳定类蛋白,脂肪系数为32.25,总平均疏水指数为0.759。该氨基酸序列功能位点的预测由在线分析工具 Predict Protein与ScanProsite 完成[20],预测结果显示其包含PKC磷酸化位点、N-酰基化位点、N-糖基化位点(图5)。利用 NCBI 网站 CDD 程序[21]对该氨基酸序列进行保守结构域分析,结果表明该蛋白属于乳球菌素972家族,结构域序列号为cl09891,结构域匹配E值为1.88e-22。利用SignalP 4.1 Serve对该氨基酸序列进行信号肽预测,结果显示其结构存在信号肽特征[22]。

图5 Lcn972A基因序列及预测的氨基酸序列Fig.5 Nucleotide sequence and predicted amino acid sequence of Lcn972A gene注:“▭”:ATG 或 TAA(ATG or TAA);“ ”:N-糖基化位点(N-glycosylation site);“-”:蛋白激酶C磷酸化作用位点(Protein kinase C phosphorylation site);“…… ”:N-酰基化作用位点(N-myristoylation site)。
使用ExPASy的ProtScale程序,在线分析氨基酸序列亲疏水性[23]。疏水性预测的方法依赖于疏水性的衡量尺度,每个氨基酸根据其一系列的物理特性(例如,溶解性、跨越水-汽相时产生的自由能等),被赋予一个数值以代表其疏水性。分析结果表明该氨基酸序列为亲水性,该氨基酸序列前端出现明显的疏水信号,进一步证明了预测是正确的,该氨基酸序列存在转运信号肽序列(图6,图中正值越大表示越疏水,负值越大表示越亲水,介于+0.5和 0.5之间的为两性氨基酸。)。
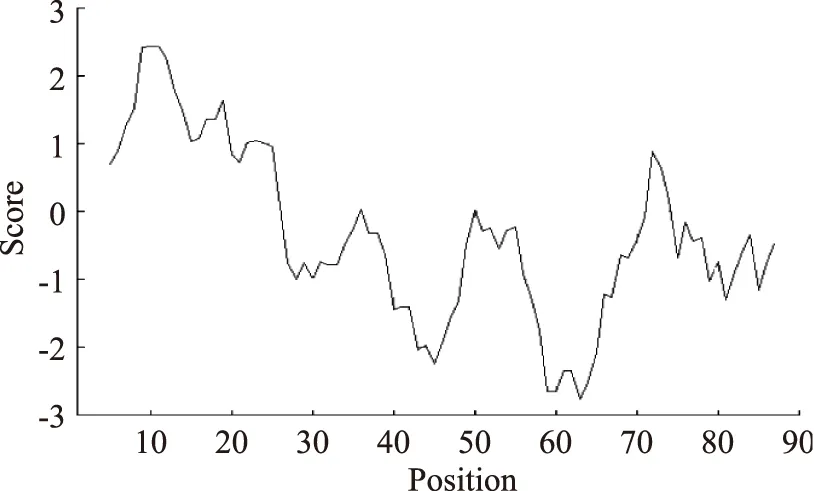
图6 氨基酸序列亲疏水性预测Fig.6 The hydrophilic predicted of amino acid sequence
在线工具Predict protein分析显示该Lcn972A氨基酸序列中,共有α-螺旋3处,占总二级结构的 37.36%;无规卷曲6处,占总二级结构的51.65%;延伸链2处,占总二级结构的10.99%。该Lcn972A主要由α-螺旋、无规卷曲和延伸链组成(图7)。
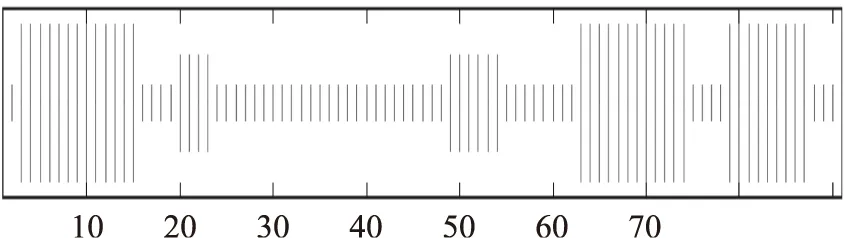
图7 蛋白质二级结构分布图Fig.7 The secondary structure of the Lcn972A protein 注:长竖条纹代表α-螺旋(α-helix),短竖条纹无规卷曲(Random coil),中竖条纹代表延伸链(Extended strand)。
用在线分析软件Phyre2 library预测该蛋白的三级结构[24],在单一最高模板模式下置信度最高可达100%,但未能完全覆盖整个结构,具有明显的空间结构,主要由α-螺旋和无规卷曲构成(图8)。

图8 Lcn972A基因细菌素三级结构图谱Fig.8 Three-Stage Prediction of the Lcn972A Protein
3 结论与讨论
近年来研究显示细菌素具有稳定的抗菌特性,且不会形成抗逆性,其中乳酸菌细菌素在食品的保存和对肠道致病菌的抑制作用方面效果明显,在食品发酵、乳制品行业中有广阔的发展前景。
传统的产细菌素乳酸菌的筛选是通过细菌素的功能性测定:首先配制合适培养基培养目的菌,通过琼脂扩散法、牛津杯法、溶钙圈法,筛选出对指示菌(单增李斯特菌等革兰氏阳性菌)具有明显抑茵作用的菌株[25-27],在排除有机酸和过氧化氢的作用后,对仍具有明显抑菌活性的菌株,用胰蛋白酶对其发酵液进行处理后,若其抑菌活性下降明显,说明代谢产物中含有蛋白性质的抑菌物质,从而确定有可能为细菌素类物质。但这类鉴定方式有明显的局限性,工作量大,操作复杂,重复性差,对于结果的把握会有误差,准确性弱,难以达到大规模高通量的筛选要求。随着大量不同乳酸菌菌株基因组测序的完成,基因组序列在挖掘新型高效的细菌素方面扮演着重要的角色,使得利用生物信息学的方法预测和寻找新型细菌素成为可能。根据已知的细菌素DNA 序列设计保守的PCR引物或制备寡核苷酸探针,可以在大范围内的乳酸菌群中挖掘出依赖功能性测定时不易发现的细菌素。其中基于PCR方法的快速筛选可以作为生态学研究中的有效手段,可以鉴定出产细菌素的益生菌,并将其应用在食品工业中[28-29]。
本实验室也采用传统的牛津杯法筛选细菌素,但抑菌圈差异不明显且耗时较多,故将分子生物学手段运用到细菌素的筛选,根据已报道的乳酸菌细菌素DNA序列设计合适引物,对杏鲍菇与海带源的乳酸菌进行分子克隆,63株乳酸菌中杏鲍菇乳酸菌A15与海带乳酸菌CX2样品含有Lcn972A细菌素基因,得到结果快速直观,对克隆得到Lcn972A细菌素基因进行序列分析,结果显示其与NCBI全基因组测序菌株细菌素基因相似性为97.46%,氨基酸序列相似性97.80%,分子量大小22843.3 u,等电点5.33,为亲水性氨基酸;因此,本实验利用基因克隆方法筛选细菌素的方法具有一定的现实意义,并且对下一步研究乳酸乳球菌中Lcn972细菌素家族的分子功能以及抑菌谱范围的确定,细菌素合成过程调控机理的揭示具有重要意义,有利于该基因与其产生菌进一步的研究。
[1]Konisky J.Colicine and other bacteriocins with established modes of action[J].Annu Rev Microbia,1982,36:125-144.
[2]Klaehammer T R.Genetics of Bacteriocins Produced by Lactic Acid Bacteria[J]. FEMS Microbiology Reviews,1993,12:39-86.
[3]Nes I F,Diep D B,Havarstein L S. Biosynthesis of Bacteriocins in Lactic Acid Bacteria[J]. Antonie van Leeuwenhoek,1996,70:113-128.
[4]Clara Roces,Verónica Pérez,Jan Kok,et al.The Putative Lactococcal Extracytoplasmic Function Anti-SigmaFactor Llmg2447 Determines Resistance to the Cell Wall-Active
Bacteriocin Lcn972[J].Antimicrobial Agents and Chemotherapy,2012,56(11):5520-5527.
[5]Cleveland J,Thomas J M,Ingolf F N,et al.Bacteriocin:Safe,natural antimicrobials for food pres-Ervation[J].Inter Food Microbial,2001,71(1):1-20.
[6]杨瑾.乳酸菌细菌素分离纯化及生物学特性[D].杭州:浙江工商大学,2007.
[7]Immacolata Anacarso,Patrizia Messi,Carla Condò,et al.A bacteriocin-like substance produced fromLactobacillus pentosus 39 is a natural antagonist for the control of Aeromonas hydrophila and Listeria monocytogenes in fresh salmon fillets[J].LWT-Food Science and Technology,2014,55(2):604-611.
[8]Mahdieh Iranmanesha,Hamid Ezzatpanaha,Naheed Mojgani.Antibacterial activity and cholesterol assimilation of lactic acidbacteria isolated from traditional Iranian dairy products[J].LWT-Food Science and Technology,2014,58(2):355-359.
[9]Noraphat Hwanhlem,Jean-Marc Chobert,Aran H-Kittikun.Bacteriocin-producing lactic acid bacteria isolated from mangrove forests in southern Thailand as potential bio-control agents in food:Isolation,screening and optimization[J].Food Control,2014,41(2014):202-211.
[10]Deraz S F,Karlsson E N,Hedstr M,et al.Purification and characterisation of acidocin D20079,abacteriocin produced by Lactobacillus acidophilus DSM 20079[J]. Journal of Biotechnology,2005,117(4):343-353.
[11]Jianyin Miao,Haoxian Guo,Yangwen Ou,et al.Purification and characterization of bacteriocin F1,a novel bacteriocin produced byLactobacillusparacaseisubsp.toleransFX-6 from Tibetan kefir,a traditional fermented milk from Tibet,China[J].Food Control,2014,42(2014)48-53.
[12]Michat Wieckowicz,Marcin Schmidt,Sip A,et al. Development of a PCR-based assay for rapid detection of class IIa bacteriocin genes[J]. Letters in Applied Microbiology,2011,52(3):281-289.
[13]Franca Rossi,Marta Marzotto,Silvia Cremonese,et al.Diversity of Streptococcus thermophilu sin bacteriocin production;inhibitory spectrum and occurrence of thermophilin genes[J].Food Microbiology,2013,35(2013):27-33.
[14]Sunita J Macwana,Peter M Muriana. A‘bacteriocin PCR array’for identification of bacteriocin related structural genes in lactic acid bacteria[J].Journal of Microbiological Methods,2012,88(2):197-204.
[15]凌代文,东秀珠.乳酸细菌分类鉴定及实验方法[M]. 北京:中国轻工业出版社,1999.
[16]仲崇艳.LB 培养基在鱼类致病菌药敏实验中的应用研究[J].动物科学,2014,13:290-292.
[17]刘晓侠,林建平,岑沛霖.微生物基因组DNA提取方法的比较与改进[J].嘉兴学院学报,2007,19(3):48-50.
[18]张洁,徐桂花,尤丽琴.16S rDNA 序列分析法鉴定乳酸菌[J].农产品加工,2009(4):47-49.
[19]Amanzadeh E,Mohabatkar H,Biria D,et al.Classification of DNA Minor and Major Grooves Binding Proteins According to the NLSs by Data Analysis Methods[J].Appl Biochem Biotechnol. 2014,174(1):437-451.
[20]Burkhard Rost,Guy Yachda and Jinfeng Liu.The predict protein server[J]. Nucleic Acids Research,2004,32:321-326.
[21]Marchler-Bauer A,Lu S N,Anderson J B,et al.CDD:a Conserved domain database for the functional annotation of proteins[J]. Nucleic Acids Res,2011,39(D):225-259.
[22]Bendtsen JD1,Nielsen H,von Heijne G,et al.Improved prediction of signal peptides:Signal P 3.0[J].J Mol Biol.2004,340(4):783-795.
[23]Xia XY,Wu QY,An LM,et al.A novel P20R mutation in the alpha-B crystallin gene causes autosomal dominant congenital posterior polar cataracts in a Chinese family[J].BMC Ophthalmol. 2014,14(1):108.
[24]Nema V,Pal SK.Exploration of freely available web-interfaces for comparative homology modelling of microbial proteins[J].Bioinformation.2013,9(15):796-801.
[25]张金兰.戊糖乳杆菌素的纯化鉴定及对托盘包装冷却肉的防腐保鲜效果研究[D].北京,中国农业大学,2009.
[26]Wolf C E,Gibbons W R.Improved method for quantification of the bacteriocin nisin[J]. Journal of Applied Bacteriology,1996,80(4):453-457.
[27]Nabil B O,Hikmate A,Keleke S,et al. Bacteriocin-producing Lactobacillus strains isolated from poto poto,a Congolese fermented maize product,and genetic fingerprinting of their plantaricin operons[J].International Journal of Food Microbiology,2008,127(1-2):18-25.
[28]诸永志,姚丽娅,徐为民,等.乳酸细菌素应用于肉制品防腐剂的研究进展[J].食品科技,2008(2):136-139.
[29]LiuW&N.Hansen.Some chemical and physical properties of Nisin,a smallprotcin antibiotic produced by Lactococcus lactis[J].Appl. Environ Microbiol,1990,56:2551-2558.
Identification of the bacteriocin Lcn972A-producing bacteria and the cloning and sequence analysis of its bacteriocin gene
GAN Xiang-wu,YE Zhi-wei,GUO Xin-yue,GUO Li-qiong,LIN Jun-fang*,WANG Yan-ting,LIAO Teng-da
(Department of Bioengineering,College of Food Science;Biotechnology institute,South China Agricultural University,Guangzhou 510640,China)
Objective:In this study,bacteriocin Lcn972A-producing bacteria were screened from 63 strains of lactic acid bacteria isolated fromPleurotuseryngiifruit body andThallusLaminariae. Subsequently,gene for this bacteriocin was cloned by PCR methods and analyzed by bioinformatics tools. Method:Primers were designed based on the results of bioinformatics analysis of reported genes for lcn972A. Subsequently,genes for Lcn927A were isolated from the genomic DNA of 63 strains of lactic acid bacteria respectively. Finally,the nucleotide sequence and predicted protein sequence were analyzed by a series of bioinformatics tools. Result:Genes for Lcn927A were isolated from 15 genomic samples among 63 strains of Lactic acid bacteria. Two of the positive stains were characterized asLactococcuslactisfrom Pleurotus eryngii fruitbody A15 and Thallus Laminariae CX2. The result of sequencing showed that the full length of the Lcn927A gene was 276 bp,which coded for 91 amino acid residues,with molecular weight of 22843.3 u,theoretical PI of 5.33,hydrophilic amino acid,the secondary structure of protein consist of α-helix,random coil and extended strand. Conclusion:The nucleotide sequence and predicted protein sequence of present Lcn927A gene ofLactococcuslactisA15 and CX2 isolated from Pleurotus eryngii fruitbody and Thallus Laminariae were different from those that hadbeen publicly reported,and the antimicrobial properties of this gene need to be identified in the future study.
Lactococcuslactis;Lcn972A gene;molecular identification;sequence analysis
2015-01-23
甘祥武(1989-),男,硕士,研究方向:食品微生物与生物技术,E-mail:gxwere@163.com。
*通讯作者:林俊芳(1962-),男,博士,研究方向:食品微生物与天然产物,E-mail:linjf@scau.edu.cn。
国家自然科学基金资助(31071837,31272217);广东省攻关项目资助(2013B010404041)。
TS201.3
A
1002-0306(2015)21-0187-05
10.13386/j.issn1002-0306.2015.21.030
猜你喜欢
天津农学院学报(2024年1期)2024-04-07 11:20:56
现代畜牧科技(2021年9期)2021-10-13 06:38:44
中华养生保健(2020年8期)2021-01-14 01:13:58
心肺血管病杂志(2020年3期)2021-01-14 00:42:38
国际呼吸杂志(2019年1期)2019-01-28 09:37:06
现代检验医学杂志(2016年2期)2016-11-14 02:38:02
环境科技(2016年3期)2016-11-08 12:14:10
腹腔镜外科杂志(2016年12期)2016-06-01 12:10:09
河北科技大学学报(2015年6期)2015-03-11 16:16:51
中国洗涤用品工业(2015年7期)2015-02-28 19:02:39
