
黄瓜强雌性状的主基因+多基因混合遗传模型分析
2015-04-29 00:44陈宸崔清志陈惠明田云1
热带作物学报 2015年10期
陈宸 崔清志 陈惠明 田云1


摘 要 利用黄瓜强雌株系S-2-98和雌雄同株系95为亲本,配制杂交组合,共获得6世代群体(P1,P2,F1,F2,B1,B2),通过观察6世代群体的雌花率,应用植物数量性状主基因+多基因联合分离分析方法,对黄瓜强雌性状的遗传模型进行判别与遗传参数的估计。结果表明:黄瓜强雌性状的遗传除受主基因控制外还受多基因的影响,符合D遗传模型,即1对加性-显性主基因+加性-显性-上位性多基因遗传模型。B1、B2、F2主基因遗传率分别为5.50%、68.98%和54.65%,相应的多基因遗传率和环境变异分别为45.67%、11.81%、25.45%和48.83%、19.21%、19.90%,说明黄瓜雌花遗传还受多基因的修饰,同时环境对黄瓜的雌花也有较大的影响。
关键词 黄瓜;强雌性状;主基因+多基因混合遗传模型;遗传
中图分类号 S642.2 文献标识码 A
Genetic Analysis of Cucumber Subgynoecious Trait Using Major
Gene Plus Polygene Mixed Genetic Model
CHEN Chen1,3, CUI Qingzhi2,4, CHEN Huiming2,3, TIAN Yun1,3 *
1 College of Bioscience and Biotechnology,Hunan Agricultural University, Changsha, Hunan 410128, China
2 Hunan Vegetable Research Institute, Changsha, Hunan 410125, China
3 Hunan Agricultural Bioengineering Research Institute, Changsha, Hunan 410128, China
4 Longping Branch of Graduate School, Central South University, Changsha, Hunan 410125, China
Abstract Six generations(P1, P2, F1, F2, B1, B2)were obtained using subgynoecious line S-2-98 and monoecious line 95 as the parental cucumber(Cucumis sativus L.), and the inheritance of female flower proportion was investigated. The model of the major gene plus polygene of quantitative traits was used to estimate the genetic model and genetic parameters of cucmber subgynoecious trait. The results showed that the cucmber subgynoecious trait was controlled by one additive-dominant-epistatic major gene plus additive-dominant-epistatic polygene mixed genetic model(model D). The major gene heritability of B1, B2,and F2 was 5.50%, 68.98% and 54.65%, respectively. While the corresponding polygene heritability and environmental variance was 45.67%, 11.81%, 25.45% and 48.83%, 19.21%, 19.90%, respectively. The results implied that the environment had great influence to female flowers rate of cucumber and should be considered in breeding for cucumber of high female flower proportion.
Key words Cucumber; Subgynoecious trait; Major gene plus polygene mixed genetic model; Inheritance
doi 10.3969/j.issn.1000-2561.2015.10.007
黄瓜(Cucumis Sativus L.)不仅是世界上重要的蔬菜作物之一,也是研究植物性别表达的重要模式植物[1]。黄瓜的花可分为雌花、雄花和两性花,3种花芽在早期都具有雄蕊和雌蕊的两性花原基,两性花原基中如果雌蕊滞育则发育成雄花,如果雄蕊滞育则发育成雌花,雄蕊和雌蕊都发育则形成两性花[2-4]。研究表明:黄瓜的性别主要由3个基因(全雌性F、两性花M和全雄系A)、其它修饰基因(雌性增强基因In-F、三性花基因Tr、隐性雌性基因gy、两性花基因m-2)与环境条件、激素等因素相互作用从而共同调控[5]。F、M、A 3种主要基因共同作用形成不同的性别类型:全雌(F_M_A_),全雄(ffM_aa),雌雄同株(ffM_A_),两性花株(FFM_A_)和雄花两性花株(ffmmA_)[6]。2011年Chen等[5]采用孟德尔分离分析方法新发现与F、M基因独立遗传的两个强雌主效基因Mod-F和mod-F2,植株末端表现为连续雌花节,但仍无法有效区分黄瓜强雌性状的主基因和多基因效应。鉴于此,本研究以黄瓜强雌株系S-2-98和雌雄同株系95为材料,配制6世代群体,采用植物数量性状主基因+多基因混合遗传模型分析方法对黄瓜强雌性状进行遗传分析,以期为黄瓜高雌花率新品种选育及种质资源创新提供依据。
1 材料与方法
1.1 材料
供试材料包括强雌株系S-2-98和雌雄同株系95,材料的性别表现、来源及有关说明见表1。
试验材料强雌型S-2-98(P1)和雌雄同株型95(P2)的雌花率分别为63%和23%,2012年春季在湖南省蔬菜研究所试验农场种植亲本P1、P2,开花时配制杂交F1代组合,秋季种植亲本P1、P2和F1代,F1代分别与双亲进行回交和自交,获得B1、B2、和F2代种子。2013年春播种杂交组合6个世代的种子,采用营养钵育苗,移栽于防虫网隔离的大棚内,各世代群体株系的样本容量分别为19(P1);8(P2);35(F1);189(F2)、18(B1)和187(B2)。观察各世代植株主蔓的第1雌花节位、雌花率、连续雌花起始节位、连续雌花节位数等性状,总节位数范围20~34节之间。
1.2 方法
采用盖钧镒[7]植物数量性状主基因+多基因混合遗传模型多世代联合分析方法。对P1×P2组合的6个世代P1、P2、F1、B1、B2和F2的黄瓜雌花率进行极大似然分析,采用AIC准则(Akaikes Information Criterion)、似然比检验以及一组适合性测验(均匀性检验、Smirnov检验和Kolmogorov检验的5个统计量U12、U22、U32、nW2、Dn),从中选出最佳最适模型及其相应的一组成分分布参数,用最小二乘法估计相应的遗传参数,并估计主基因和多基因效应值、方差和遗传率等遗传参数。
2 结果与分析
2.1 亲本及各世代群体雌花率的分布
雌雄同株黄瓜品种分为两类。一类为普通雌雄同株,雌花率较低,平均为25%~45%左右,雄花节位数与雌花节位数比率为3以上,末端无连续性雌花节位。另一类为雌雄同株的强雌株(简称强雌株),第1~7节多数为雄花,雌花率较高,为64%~94%,雄花节位数与雌花节位数比率为0.49~0.63,末端8~11节以上为连续的雌花节位。S-2-98×95配制的6世代群体的雌花率的结果见表2。P1亲本雌花率平均值为0.805,P2亲本雌花率平均值为0.339,F1和B1代雌花平均值分别为0.684和0.693,雌花率均介于双亲之间,略高于中亲值。而F2及B2分离世代雌花率变异均为多峰偏态连续分布,呈现出明显的主基因+多基因遗传的特征。
2.2 遗传模型适应性检测
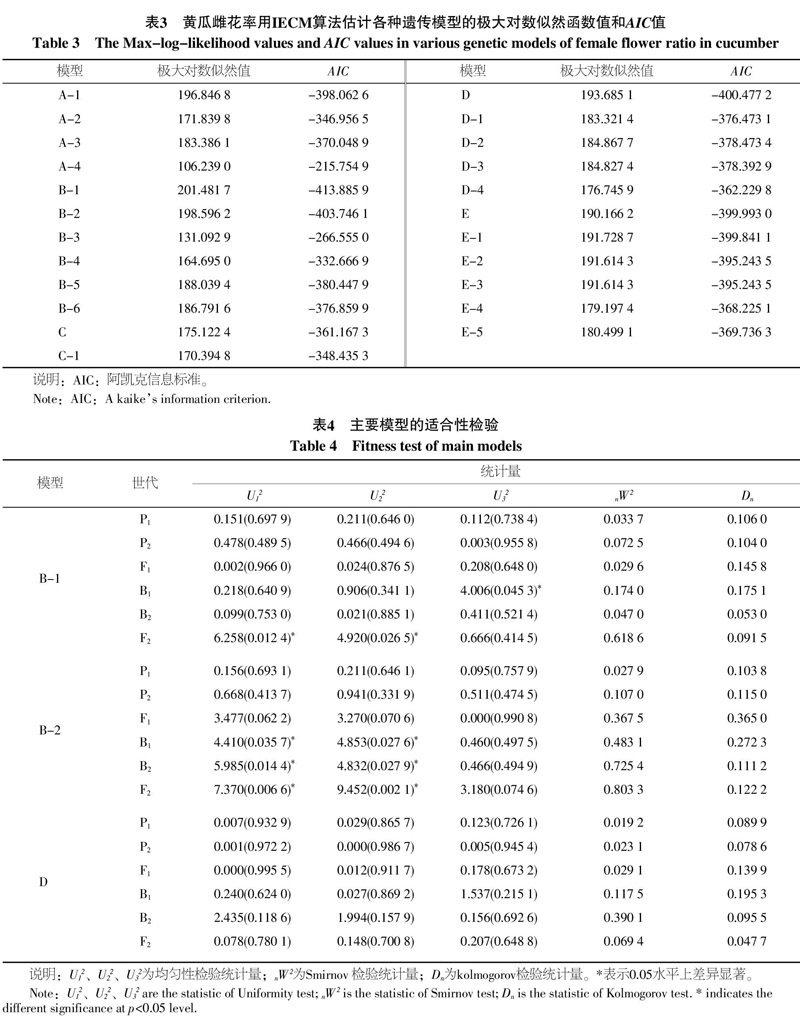
通过对P1、P2、F1、F2、B1和B2共6个世代黄瓜植物强雌花率进行联合分离分析,分别计算出5类23种不同模型的极大对数似然值和AIC值,结果列于表3。B-1模型具有最小AIC值,为最佳可能模型,与之相近的还有B-2、D、E、E-1模型。但经一组适合性检验的结果(表4),B-1虽然AIC值最低,但适合性检验有3个差异显著,B-2的AIC值相对第二小,同样适合性检验有6个差异显著,D模型AIC值第三小,其适合性检验没有发现差异显著,AIC第4、5小的E、E-1模型的适合性检验分别也没有差异显著性。因此,选择D模型为最优模型,黄瓜强雌性基因遗传属于一对主基因+多基因混合遗传模型(一对加性-显性主基因+加性-显性-上位性多基因遗传模型)。
2.3 遗传参数估计
在D模型下,该模型的极大似然估计值见表5,μ1、μ2、μ3为P1、P2、F1世代中的单个主效基因型平均值,μ41、μ42和μ51、μ52为B1和B2世代中的2种主效基因平均值,μ61、μ62、μ63为F2代世代中3种主效基因型平均值,σ2、σ24、σ25、σ26分别为F1、B1、B2和F2各成分分布方差。
根据D模型不同成分的分布参数,计算出的极大似然估计值,并由此计算出各有关一阶和二阶遗传参数。从表6可以看出,黄瓜雌花率性状的主基因的加性效应值为0.152 8,显性效应值0.153 0,说明亲本的高的雌花率对后代有一定的影响,在育种实践中,要选择雌花率较高的双亲,以提高后代的雌花率,同时主基因的杂种优势较明显,配制杂交新组合要考虑主基因的特殊配合力。主基因遗传率在B1、B2和F2世代中分别为5.50%、68.98%和54.65%,B2世代中的主基因遗传率最高;多基因遗传率则在B1世代中最高,为45.67%,其次是B2和F2,分别为11.81%和25.45%。这表明控制S-2-98×95组合强雌系基因主要为呈1对主基因控制,主基因遗传率大于相应世代的多基因遗传率,尤其表现在B2和F2代,同时其遗传还受多基因的修饰。各群体的环境变异为19.21%~48.83%,可见环境对黄瓜的雌花率也有较大的影响。
3 讨论与结论
前人采用世代平均数法和方差组分法等经典数量遗传学方法研究黄瓜雌花率,都只能估测多基因的总体效应,或者采用孟德尔分离分析方法追踪个别末端表现连续雌花的强雌性基因或全雌性基因[8-9]。近年来,盖钧镒等[7]提出了植物数量性状主基因+多基因遗传理论,即数量性状可以由大小不等的多对基因控制,大者表现为可个别检测的主基因,小者表现为难以在实验条件下个别检测的微效基因,数量性状由主基因与多基因共同控制是数量性状遗传的基本模型或普遍模型,而纯主基因或纯多基因的遗传模型只是特例。以此理论为依据,形成了植物数量性状主基因+多基因混合遗传模型分析方法。该方法在小麦、烟草、水稻、油茶、甜瓜、不结球白菜、黄瓜等农作物的主要经济性状遗传分析方面得到了广泛应用[10-16]。
本研究采用这一方法对组合S-2-98×95的6个世代群体(P1、P2、F1、F2、B1、B2)进行了联合分析,明确了黄瓜强雌性基因受一对主基因+多基因混合遗传模型(D模型),区分了主基因和多基因效应。进一步证实了黄瓜强雌性状受主基因、多基因和环境因素等共同控制,至少存在1对效应较大的主基因。这与时秋香[17]采用同样研究材料通过对强雌性基因QTL分析,共得到3个QTL:Mod-F1a、Mod-F1b、Mod-F1c。其中Mod-F1a位于第三号染色体,是主效QTL,遗传贡献率为49.8%~54.6%;Mod-F1b与Mod-F1c位于第六号染色体,是微效QTL,遗传贡献率分别为6.3%~7.2%和4.2%~9.1%,这一结果与本研究结果基本吻合。
参考文献
[1] 陈惠明, 卢向阳, 许 亮, 等. 黄瓜性别决定相关基因和性别表达机制[J]. 植物生理学通讯, 2005, 41(1): 7-13.
[2] Atsmon D, Galun E. A morphogenetic study of staminate, pistillate and hermaphrodite flowers in Cucumis sativus L[J]. Phytomorphology, 1960, 10: 110-115.
[3] Goffinet M C, Bates D M, Robinson R W. Comparative ontogeny of male and female flowers of Cucumis sativus in biology and utilization of the cucurbitaceae[M]. New York: Cornell University Press, 1990: 288-304.
[4] Malepszy S, Niemirowicz-Szczytt K. Sex determination in cucumber(Cucumis sativus L.)as a model system for molecular biology[J]. Plant Sci, 1991, 80(1-2): 39-47.
[5] Chen H, Tian Y, Lu X, et al. The inheritance of two novel subgynoecious genes in cucumber(Cucumis sativus L.)[J]. Sci Hortic, 2011, 127(3): 464-467.
[6] Perl-Treves R. Male to female conversion along the cucumber shoot: Approaches to studying sex genes and floral development in Cucumis sativus. In Sex Determination in Plants[M]. UK: Bios Scientific Publishers, 1999: 189-216.
[7] 盖钧镒, 章元明, 王建康. 植物数量性状遗传体系[M]. 北京: 科学出版社, 2003: 224-260.
[8] Galun E. Study of the inheritance of sex expression in the cucumber. The interaction of major genes with modifying genetic and non-genetic factors[J]. Genetica, 1962, 32(1): 134-163
[9] Kubicki B. Investigations of sex determination in cucumber.(Cucumis sativus L.). V. Genes controlling intensity of femaleness[J]. Genet Polon, 1969, 10(1-2): 69-86
[10] Irfaq M, Ajab M, Khattak G S, et al. Genetic behavior of controlling area under disease progress curve for stripe rust(Puccinia striiformis f. sp. tritici)in two wheat(Triticum aestivum)crosses[J]. Phytopathology, 2009, 99(11): 1 265-1 272.
[11] 段韫丹, 邱 杨, 汪精磊, 等. 萝卜不同抗源对黑腐病抗性的遗传分析[J]. 植物遗传资源学报, 2015, 16(1): 1-6.
[12] 陈小翠, 代帅帅, 张兴伟, 等. 烤烟CMV抗性的主基因+多基因混合遗传模型分析[J]. 植物遗传资源学报, 2014, 15(6): 1 278-1 286.
[13] 周清元, 崔 翠, 阴 涛, 等. 甘蓝型油菜角果长度的主基因+多基因混合遗传模型[J]. 作物学报, 2014, 40(8): 1 493-1 500.
[14] 林婷婷, 王建军, 王 立, 等. 不结球白菜维生素C含量主基因+多基因遗传分析[J]. 作物学报, 2014, 40(10): 1 733-1 739.
[15] 张 宁, 张 显, 张 勇, 等. 甜瓜远缘群体果实糖含量相关性状遗传分析[J]. 植物遗传资源学报, 2014, 15(5): 932-939.
[16] 闫立英, 娄丽娜, 李晓丽, 等. 雌雄同株黄瓜单性结实性遗传分析[J]. 中国农业科学, 2010, 43(6): 1 295-1 301.
[17] 时秋香. 黄瓜性别决定基因M与强雌性基因QTL定位[D]. 济南: 山东农业大学, 2009.
猜你喜欢
区域治理(2022年40期)2022-11-27
疯狂英语·新悦读(2022年8期)2022-09-20
中老年保健(2021年5期)2021-08-24
华人时刊(2021年21期)2021-03-09
今日农业(2020年16期)2020-12-14
小学生学习指导(低年级)(2020年10期)2020-11-09
作文大王·低年级(2019年11期)2019-12-02
动漫界·幼教365(小班)(2019年10期)2019-10-28
动漫界·幼教365(大班)(2019年10期)2019-10-28
动漫界·幼教365(中班)(2019年10期)2019-10-28
