
耐低温弱光结球甘蓝种质资源评价
2015-04-02 07:57李建斌李兆虎王神云丁万霞
江苏农业学报 2015年3期
李建斌, 李兆虎, 王 红, 于 利, 王神云, 丁万霞
(1.江苏省农业科学院蔬菜研究所,江苏 南京210014;2.江苏省睢宁县王集试验站,江苏 睢宁221211)
冬春设施栽培特殊的生长环境(低温弱光、早晚湿度大等)严重影响作物的正常生长,降低产量和品质[1]。低温弱光降低了黄瓜、花椰菜等的叶绿素含量及RuBP 羧化酶活性,使得叶片气孔关闭,影响光合作用[2-3],因而显著降低了净光合速率。低温弱光胁迫使黄瓜、甜椒PSⅠ(光系统Ⅰ)发生光抑制,使光合作用下降[4]。低温弱光条件下辣椒叶片光合色素含量、净光合速率、气孔导度、胞间CO2浓度、最大光化学效率、光合电子传递量子效率等光合生理生态指标降低,影响光合作用[5-6]。低温降低番茄的光合能力,诱导结球甘蓝通过春化,发生先期抽薹现象,而且低温对结球甘蓝叶片的超微结构和Ca2+分布有明显的影响[7-9],间接影响光合作用。
结球甘蓝设施栽培是早熟春结球甘蓝栽培的主要模式之一,面积日益扩大,尤其在中国黄淮海流域及其以北地区。通过设施栽培,春结球甘蓝可提前15 d 天左右上市,通过调节定植期,可在春节前后上市,补充蔬菜市场淡季供应,还可以和其他果菜类蔬菜间套作,增加经济效益。设施栽培可减少病虫害,少用农药,减少环境污染[10-11]。目前,生产上适宜设施栽培的耐低温弱光的结球甘蓝品种相对匮乏,因此急需从筛选优异的耐低温弱光种质资源入手,开展育种和研究工作。本研究对江苏省农业科学院蔬菜研究所保存的79 份结球甘蓝种质资源进行生物学性状和光合能力评价和筛选,挖掘耐低温弱光、适宜设施栽培的种质资源,为今后设施栽培结球甘蓝育种和研究提供基础材料。
1 材料和方法
1.1 试验地概况
试验地设在江苏省徐州市睢宁县王集镇(北纬33°53',东经117°56'),位于江苏省北部,属暖温带半湿润季风气候,四季分明,无霜期210 d,北与山东省接壤。本试验采用寿光5 代日光温室。
1.2 试验材料
试验材料为江苏省农业科学院蔬菜研究所提供的结球结球甘蓝种质资源79 份,即S1 ~S79 (18 份尖球形,61 份圆球形)。2013 年12 月1 日采用双膜一帘漂浮盘育苗,次年1 月15 日(苗龄4 ~5 片真叶)定植于日光温室内,株行距30 cm×35 cm,每个材料种植40 株,3 次重复,随机区组排列,常规田间管理。
1.3 生物学性状调查
2014 年3 月10 日至20 日,结球甘蓝成熟时,每个小区选取5 株进行开展度(植株开展最大处)、叶球质量、外叶质量、外叶数(片)、病害(霜霉病、黑腐病和菌核病,以影响植株正常生长或产品商品性为标准)、抽薹率(有现蕾现象为抽薹)等低温弱光条件下与结球甘蓝种植密切相关的生物学性状调查〛
1.4 光合生理指标测定
2014 年3 月1 日,所有材料进入结球期时,采用便携式光合作用测定系统(Li-6400,美国)进行光合作用测定。测定时取每株距生长点第3 张完全展开叶片(大小、颜色基本一致),保持叶片自然生长角度,每个材料测定10 株,每株重复读数3 次。测定时间选晴朗无风光照充足的3 d,从早上9∶ 00到晚上17∶ 00,中午12∶ 30 -13∶ 30 停测1 h。叶室(2 cm×3 cm)内设定温度(20 ±2)℃,大气CO2浓度为(380 ± 20)μmol/(m2·s),光照度为1 000 μmol/(m2·s)[12],测定包括净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)等光合生理指标。
1.5 数据统计分析
采用Excel 2007 对数据进行描述性统计作图,用SPSS19.0 统计分析软件对数据进行相关性分析。
2 结果与分析
2.1 不同种质资源的生物学性状
2.1.1 抽薹性和抗病性 耐抽薹和抗病是筛选育种材料的首要条件。3 月20 日,通过田间材料抽薹性调查发现,供试79 份材料中,S77 和S15 叶球生长没有达到采收的程度,生育期长,熟性偏晚,2 份均为尖球形,不适宜低温弱光条件下设施栽培。低温弱光条件下,经过在一定低温下的生长,大部分材料冬性较强,没有发生抽薹现象。有8 份材料冬性弱,通过春化,发生抽薹现象,其中5 份圆球形,3 份尖球形。S75 抽薹植株最多,抽薹率达到87.5%,S53 和S60 2 个材料抽薹率为40.0%,虽然比S75低47.5%(表1),但早春结球甘蓝栽培,只要发生抽薹现象,给生产会带来严重的损失,所以无论抽薹率高低,只要发生抽薹现象,就淘汰。有6 份材料发生病害,影响结球甘蓝正常生长或产品形成,S18 霜霉病严重,100%发病,圆球形;S19 和S20 发生黑腐病,均为圆球形,不仅影响生长,而且叶球商品性极差;S61、S74 和S78 发生菌核病,球腐烂,严重影响产品器官,均为尖球型;S78 既发生抽薹又有菌核病(表1)。根据抽薹性和病害情况,淘汰13 份材料。

表1 结球结球甘蓝种质抽薹和发病材料表现Table 1 The information of bolting and diseased materials of cabbage (Brassica oleracea var. Capitata)germplasm
2.1.2 其他生物学特性 开展度、单球质量、外叶数等生物学特性是鉴定结球甘蓝种质资源的重要指标。通过熟性、抽薹性和病害调查淘汰15 个材料。剩余64 个材料其他重要生物学性状见表2,11 个尖球形材料,53 个圆球型材料。64 个材料的单球质量在0.37 kg 至1.12 kg 之间。S37 单球质量最小,仅0.37 kg;S3 最大,达到1.12 kg。根据不同地域生产和消费需求,选择适宜的球形进行组配,同时要考虑到新品种的单球质量,可以组配单球质量达到0.5 ~2.0 kg 的新品种。所有材料开展度在28.5 cm 至47.8 cm 之间,可以进行组配,但值得注意的是组配时2 个开展度偏大的材料,易导致后代开展度偏大。S12 和S13外叶数分别是10 片和12 片,稍微偏多,如果植株外叶数多,会影响通风和透光情况,增加病害的发生,不适宜低温弱光设施条件下栽培,更不适宜套作。其他材料的外叶数为6 ~9 片,7 片叶居多。S37、S71、S72外叶质量占球质量50%以上,说明光合产物利用率不高,S72 外叶质量达到球质量的56.53%,而S3 仅达到球质量的15.63%。根据其他生物学特性的初步筛选结果,淘汰5 份材料。
2.2 光合生理指标及与生物产量的关系
2.2.1 光合生理指标基本特性 光合作用是植物生物产量的主要决定因素之一。3 月5 日测定不同材料的光合参数,评价剩余59 份结球甘蓝种质资源耐低温弱光的能力。光合参数指标测定结果见图1,59 份材料的净光合速率(Pn)平均值为21.90 μmol/(m2·s),分布在14.57 μmol/(m2·s)(S27)和31.26 μmol/(m2·s)(S3)之间,最大值和最小值相差16.69 μmol/(m2·s),大部分材料主要集中在20 ~ 25 μmol/(m2·s),2 份 材 料 低 于 15 μmol/(m2·s)(S27 和 S34),2 份材料超过30 μmol/(m2·s)(S2 和S3)。Pn越大说明材料的光合利用率相对越高。不同材料的Pn差异较大,反映了不同材料对低温弱光的适应能力差异较大,在组配选育适应设施栽培的新品种时可作为一个参考指标,不要选择2 个光合能力弱的材料进行组配,尽量选择2 个强光合能力,或者一强一弱的组配。
叶片的气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)等指标对植株的光合作用也有一定的影响作用。Gs是评价植物叶片气孔调节能力的重要指标,与叶片CO2吸收能力有很大关系。本试验在特定光照度和特定相对湿度条件下测得59 份材料的Gs,其中S11 最低,仅为0.25 mol/(m2·s),S1 最高,达到0.71 mol/(m2·s)。Gs最大和最小并非与Pn同步,大部分材料Gs主要集中在0.4 ~0.6 mol/(m2·s)。CO2是光合作用的反应物之一,Ci在一定范围内越大,光合能力就越强。59 份材料的Ci主要集中在200 ~300 μmol/mol,S11 的Ci最低,为209.00 μmol/mol,与Gs同步;S1 和S36 Ci达到330 μmol/mol以上;S35 最高,达到426.62 μmol/mol。Tr是植物在一定时间内单位叶面积蒸腾的水量,反应植物对水分的利用效率。59 份材料中Tr最小的是S26,为2.18 mmol/(m2·s);最大是S66,为6.65 mmol/(m2·s)。

表2 结球结球甘蓝种质材料的生物学性状Table 2 The biological traits of cabbage
通过上述光合参数指标的分析,发现S2 和S3光合能力最强,Gs、Ci、Tr3 个参数分别大约是59 份材 料 的 平 均 值 0.53 mol/(m2·s)、258.67 μmol/mol、4.37 mmol/(m2·s)。S27 光合能力最弱,但Tr为2.44 mmol/(m2·s),可能对水的利用率比较高,Gs和Ci基本趋于平均水平。S11 的Gs和Ci均最小,光合能力也低于平均值 21.93 μmol/(m2·s)。

图1 结球结球甘蓝种质材料的光合生理指标Fig.1 The photosynthetic indexes of cabbage
2.2.2 不同材料光合生理指标之间的关系 不同作物光合生理指标存在差异,同一作物不同材料之间也有很大的差别。本试验59 份结球甘蓝材料光合生理指标之间的相关性存在很大差异(表3)。Pn与Gs、Ci、Tr的相关性比较复杂。Pn与Gs的相关性,59 份材料中3 份极显著正相关,6 份显著正相关,1 份极显著负相关,3 份显著负相关;S41 相关系数高达0.833,该材料的光合作用主要受气孔因素限制,而S46 的相关系数为-0.801,表明该材料的光合作用主要受气孔因素负调控,共有20 个材料Pn与Gs的相关系数为负数。Pn与Tr的相关性,59 份材料中2 份极显著正相关,8 份显著正相关,1 份极显著负相关,3 份显著负相关;S41 相关系数高达0.832,而S46 的相关系数为-0.834,共有21 个材料的Pn与Tr的相关系数为负值。Pn与Ci的相关性,59 份材料中7 份极显著负相关,10 份显著负相关,有15 份材料正相关,而且相关性弱,S47 的相关系数为-0.939,显著负相关,S10 的相关系数为0.497。Pn与Gs和Tr的相关系数基本趋势一致,而与Ci的相关性主要趋于负相关,但相对复杂。Gs与Tr的相关性,59 份材料中48 份极显著正相关,7 份显著正相关,S17、S68、S73 3 份材料的相关系数分别为0.627、0.544 和0.564。大部分材料Gs和蒸腾作用相关性极大,Gs明显影响Tr,Gs越大蒸腾作用越强;Gs与Ci的相关性,59 份材料中34 份材料极显著正相关,12 份显著正相关,1 份负相关,S35 的相关系数为-0.483,可能与植株的CO2利用率有关系。Tr与Ci的相关性,59 份材料中24 份极显著正相关,12份显著正相关,1 份显著负相关。蒸腾作用和CO2浓度基本趋于正相关,可能主要是因为Gs的中介作用。S46 和S47 2 个材料4 个光合生理生态指标之间的关系比较紧密,相关性比较大。
2.2.3 结球甘蓝光合生理指标与生物产量之间的关系 在植物生长过程中,生物产量和光合作用有着密不可分的关系。59 个材料的生物产量与Pn的相关系数达到0.584,极显著正相关,与Gs的相关系数是0.168,与Tr和Ci的相关系数分别为0.083 和-0.079。所以,影响结球甘蓝生物产量的光合参数指标大小依次为Pn>Gs>Tr>Ci。
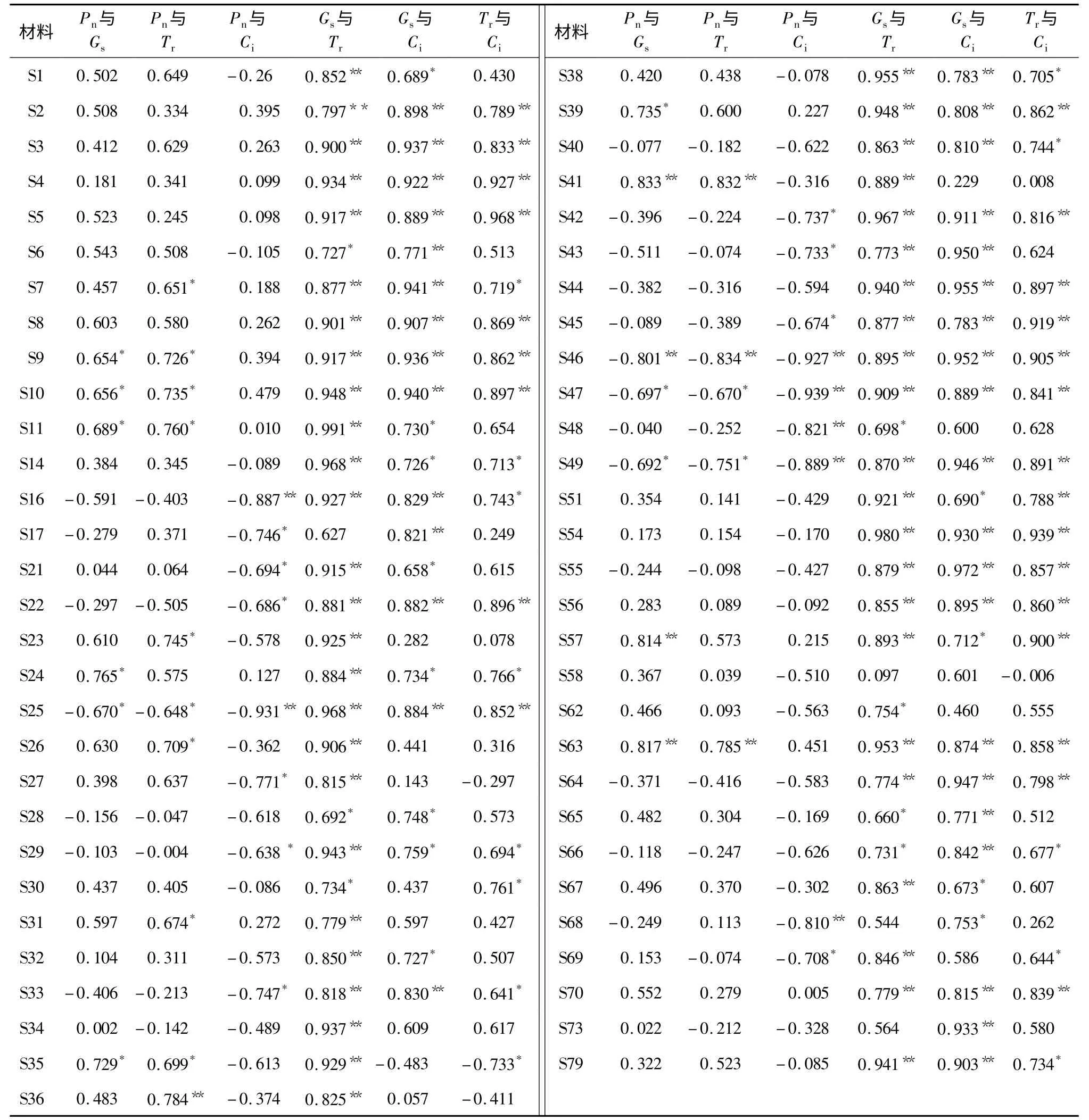
表3 结球结球甘蓝种质材料的光合生理指标相关性分析Table 3 Correlation analysis among photosynthetic indexes of cabbage
3 讨论
3.1 耐低温弱光结球甘蓝种质评价和筛选方法
本试验采用生物学性状观察调查与光合生理指标分析相结合的方法,进行耐低温弱光结球甘蓝种质资源的评价和筛选。适宜低温弱光设施栽培的专性资源对某些生物学性状和光合能力要求稍高,如保护地种植的黄瓜、番茄、辣椒品种主要要求是耐低温弱光、抗病性强、早熟等[13-15]。低温弱光条件下,首先考虑到结球甘蓝种质资源的冬性强弱,冬性强遇低温不易发生抽薹。由于结球甘蓝需经过低温春化才能开花,且需要的产品是叶球,所以该性状的要求与其他茄果类等蔬菜不同,本试验将发生抽薹的8 个材料全部淘汰。其次是抗病性,抗病性弱会导致减产甚至绝收,所以抗病性是任何作物特定条件下生产必须考虑的因素之一[16]。本试验通过调查病害情况,淘汰6 个感病材料。结球甘蓝的其他一些生物学性状,如开展度、外叶数、丰产性、叶与叶球质量比等,是种质资源筛选的基本要求。植株的开展度太大,占地面积大,不适宜密植,影响通风,在组配时是主要考虑的因素之一;丰产性是育种的基本目标,所以该性状也是评价和筛选的一个重要指标;叶与叶球质量比反映光合产物的利用率,叶与叶球质量比高,说明光合产物对外叶的贡献率高,对产品的贡献率低,所以在特殊的环境条件下,尽量选择叶与叶球质量比小的资源,外叶数多也说明光合产物对产品的贡献率低,而且影响通风。本试验通过调查和观察,筛选掉S15 和S77 2 份熟性偏晚的材料、S12 和S13 2 份外叶数偏多的材料以及S37、S71、S72 3 份外叶质量占球质量50%以上的材料。低温弱光条件直接或间接影响作物的光合作用[2-3],所以通过光合生理指标测定分析评价结球甘蓝种质资源耐低温弱光能力,类似于辣椒和番茄耐低温弱光评价指标[8,14],筛选种质资源,为新品种选育提供依据。耐低温弱光性要求种质的光合能力强、冬性强(不易抽薹)、抗寒性强,所以需通过合理科学的评价和筛选方法获得优异的种质资源。
3.2 光合生理指标评价结球甘蓝种质资源耐低温弱光能力
光合作用为作物生物产量的形成提供了主要的物质基础,所以对材料光合特性的了解,可以指导育种与生产。本试验采用光合生理指标对59份结球甘蓝材料进行光合能力评价,这些材料中最大净光合速率(Pn)是31. 26 μmol/(m2·s)(S3),最小是14.57 μmol/(m2·s)(S27),两者相差16.69 μmol/(m2·s),差异较大,说明不同材料对低温弱光的适应能力有差异。不同辣椒品种和油桐品种等也存在光合效率的差异[4,15,17]。气孔导度(Gs)是气孔张开的程度,即叶片与外界进行气体交换的程度,调节气孔开度的大小使植物在损失水分较少的条件下获取最多的CO2,控制着植物的光合作用和蒸腾作用[18]。本试验中大部分材料Gs与蒸腾速率(Tr)、胞间CO2浓度(Ci)存在正相关关系。本试验59 份材料的Gs在0. 25 mol/(m2·s)(S11)和0. 71 mol/(m2·s)(S1)之间,主 要 分 布 在 0. 4 mol/(m2·s)至 0. 6 mol/(m2·s)之间,每个材料气孔调节能力有差异,适应环境气体变化的能力不同。Ci是外界CO2气体进入叶肉细胞过程中所受各种驱动力和阻力以及叶片内部光合作用和呼吸作用最终平衡的结果[19-20]。本试验59 份材料的Ci主要集中在200 μmol/mol和300 μmol/mol之间。蒸腾作用是植物对水分吸收和运输的主要动力,促进矿质元素在植物体内运输,降低植物叶片温度,使植物体内许多生理活动正常进行[21]。59 份材料Tr最小为2.18 mol/(m2·s)(S26 ),最 大 为 6. 65 mol/(m2·s)(S66)。王美云等认为Tr越小对水分的利用率越高[22],S26 可能对水分的利用率最高。
本试验研究了不同结球甘蓝材料光合生理指标之间的关系,发现不同材料不同指标之间相关性差异较大。Pn与Gs、Tr、Ci之间的相关性不大,但有少数材料之间存在显著相关。Pn与Gs相关系数,3 份材料极显著正相关,6 份材料显著正相关,说明这些材料的光合作用主要受气孔因素限制;1 份材料极显著负相关,3 份材料显著负相关,说明这些材料的光合作用主要受气孔因素负调控。导致Pn差异的既有气孔因素也有非气孔因素。气孔导度与净光合速率之间的耦合系数体现了不同作物之间水-碳交换比例的差异,反映了气孔限制和内部生理生化过程限制在不同作物上所占的比例[23]。Pn与Gs相关性小或不相关,说明Pn的差异不是Gs差异导致CO2供应差异所致,而是由于非气孔因素(叶肉细胞使气孔扩散的阻抗增加、CO2溶解度下降、Rubisco 酶对CO2的亲和力降低、RuBP 再生能力下降或光合机构关键成分的稳定性因逆境胁迫而降低等)阻碍了CO2的利用[24-25],本试验59 份材料中46 份材料的Pn与Gs相关性不强。许大全认为Ci下降,引起光合速率降低[24],本试验59 份材料中Pn与Ci的相关性比较复杂,其中7 份材料极显著负相关,10 份材料显著负相关,有15 份材料正相关,其余都是负相关。陈根云等发现水稻和小麦的Pn与Ci之间也存在复杂的相关关系[26]。结球甘蓝Pn与Tr之间的关系也比较复杂,59 份材料中2 份极显著正相关,8 份显著正相关,1 份极显著负相关,3 份显著负相关。其他作物,如油桐,Pn与Tr的日变化有很强的正相关关系[25],与结球甘蓝不同,可能与研究的对象和群体大小有关。因为在不同情况下决定叶片光合速率高低的主要因子不同,不同因子之间的相互作用是错综复杂的[26]。
本试验研究发现影响结球甘蓝生物产量的光合参数指标大小依次为Pn>Gs>Tr>Ci,而且Pn对生物产量有极显著的正相关性。
本试验采用了较合理的评价、鉴定方法,获得了一批优异的耐低温弱光的结球甘蓝种质资源。
[1] 艾希珍,张振贤,何启伟,等. 日光温室主要生态因子变化规律及其对黄瓜光合作用的影响[J]. 应用与环境生物学报,2002,8(1):41-46.
[2] 张红梅,余纪柱,金海军.低温弱光对黄瓜植株生长、光合特性的影响[J].沈阳农业大学学报,2006,37(3):339-342.
[3] 李光庆,谢祝捷,姚雪琴,等.花椰菜叶绿素荧光参数与耐寒性的关系研究[J].园艺学报,2010,37(12):2001-2006.
[4] LI X G,MENG Q W,JIANG G Q,et al. The susceptibility of cucumberand sweet pepper to chilling under low irradiance is related to energydissipation and water-water cycle[J]. Photosynthetica,2003,41(2):259-265.
[5] 张志刚,尚庆茂. 低温、弱光及盐胁迫下辣椒叶片的光合特性[J]. 中国农业科学,2010,43(1):123-131.
[6] 何 勇,符庆功,朱祝军. 低温弱光对辣椒叶片光合作用、叶绿素荧光猝灭及光能分配的影响[J]. 核农学报,2013,27(4):479-486.
[7] 毛丽萍,任 君,张剑国,等. 日光温室秋冬茬番茄果实发育期的适宜夜温[J]. 生态应用学报,2014,25(5):1408-1414.
[8] 韩灿功,赵凤莲,石红梅,等.几起越冬甘蓝发生未熟抽薹的调查分析[J].中国蔬菜,2005(5):38-39.
[9] 曾爱松,严继勇,宋丽晓,等.甘蓝幼苗叶片超微结构及细胞内Ca2+分布对低温的响应[J]. 华北农学报,2011,26(6):129-135.
[10] 安匀彬. 日光温室甘蓝栽培技术[J]. 中国蔬菜,2009 (3):38-39.
[11] 孙永生,赵 珺,崔秀琴. 早春日光温室番茄与结球甘蓝套作栽培技术[J]. 辽宁农业科学,2006(4):62.
[12] 王美玲,艾希珍,丁 飞,等. 紫结球甘蓝和普通甘蓝光合特性的比较[J]. 园艺学报,2008,35(4):547-552.
[13] 李建吾,马长生,孙治强,等. 保护地黄瓜新品种新世纪[J].园艺学报,2005,32(4):763.
[14] 吴秀娟,裴孝伯,吴良锁,等. 合肥地区春季大棚光温环境与番茄耐低温弱光指标的筛选[J]. 中国农学通报,2009,25(4):162-166.
[15] 董言香,张其安,方 凌,等. 紫色辣椒新品种紫燕1 号[J].园艺学报,2007,34(6):1595.
[16] 王忠华,贾育林,夏英武. 植物抗病分子机制研究进展[J]. 植物学通报,2004,21(5):521-530.
[17] 李 泽,谭晓风,袁 军,等. 4 个油桐品种光合特性的日变化研究[J]. 中国农学通报,2013,29(25):12-15.
[18] 于 强,谢贤群,孙菽芬,等. 植物光合生产能力与冠层蒸散模拟研究进展[J]. 生态学报,1999,19(5):744-753.
[19] 原向阳,郭平毅,张丽光,等. 草甘膦对大豆光合特性日变化的影响[J]. 山西农业大学学报:自然科学版,2010,30(6):500-504.
[20] 范彩霞,刘桂花,周 敏,等. 3 种阔叶树幼树光合特性的比较研究[J]. 安徽农业大学学报,2010,37(2):324-327.
[21] 陈兆波,张 翼,王 沛,等. 香紫苏开花期蒸腾和光合作用日变化特征及其影响因子研究[J]. 西北植物学报,2007,27(6):1202-1208.
[22] 王美云,李少昆,赵 明. 关于玉米光合作用与叶片水分利用效率关系的研究[J]. 作物学报,1997,23(3):345-352.
[23] 王建林,林荣芳,于贵瑞,等. 光合CO2作用下C3 和C4 作物气孔导度-光合速率耦合关系的差异[J]. 华北农学报,2008,23(1):71-75.
[24] 许大全. 光合效率[M]. 上海:上海科学技术出版社,2002:32-34.
[25] FLEXAS J,MEDRANO H. Drought-inhibition of photosynthesis in C3 plants:Stomatal and non-stomatal limitations revisited[J].Annalsof Botany,2002,89(2):183-189.
[26] 陈根云,陈 娟,许大全. 关于净光合速率和胞间CO2浓度关系的思考[J]. 植物生理学通讯,2010,46(1):64-66.
猜你喜欢
天天爱科学(2022年12期)2022-11-10
动漫界·幼教365(中班)(2020年8期)2020-06-29
现代园艺(2017年23期)2018-01-18
上海农业学报(2017年3期)2017-04-10
长江蔬菜(2016年10期)2016-12-01
上海农业学报(2016年5期)2016-02-10
上海蔬菜(2015年2期)2015-12-26
动物营养学报(2015年10期)2015-12-01
苏州科技大学学报(工程技术版)(2015年3期)2015-02-28
农家科技中旬版(2014年1期)2014-04-18
