
A General Overview of the Typical18Frontal-Ventral-Transverse Cirri Oxytrichidae s. l. Genera (Ciliophora, Hypotrichia)
2015-03-31 05:43SHAOChenLUXiaotengandMAHonggang
SHAO Chen, LU Xiaoteng, and MA Honggang
A General Overview of the Typical18Frontal-Ventral-Transverse Cirri OxytrichidaeGenera (Ciliophora, Hypotrichia)
SHAO Chen1), *, LU Xiaoteng1), 2), and MA Honggang2)
1),,,710049,.2),,,266003,
Oxytrichidaeciliates usually have 18 frontal-ventral-transverse cirri which are clustered to six distinct groups usually originating from six longitudinal primordia segregating 1, 3, 3, 3, 4, 4 cirri. During morphogenesis, three dorsal kineties anlagen are primarily formed. Fragmentation of kinety 3 usually present, while sometimes secondarily lost. Dorsomarginal kineties are formed, while sometimes lost. Oxytrichids tend to have overlapping characters,cell shape and size, infraciliature, pellicle features. This makes a great problem for genera separation. In the present work, all typical 18 frontal-ventral-transverse-cirri Oxytrichidaegenera were revised systematically based on their living morphology, ciliature patterns and dorsal morphogenetic features. The outline of the genera, the schematic illustrations, and the key to typical 18 frontal-ventral-transverse-cirri genera of Oxytrichidaewere clarified. Additionally, some morphological and morphogenetic patterns were summarized and compared.
Oxytrichinae; Sporadotrichida; Stylonychinae; taxon
1 Introduction
Species of the well-known hypotrichous ciliates are of highly diverse regarding their structure, morphology and ontogenetic patterns, and are commonly found in various habitats including marine, limnetic and terrestrial biotopes (Chen., 2013a, b; Foissner, 2012; Foissner and Stoeck, 2011; Foissner., 2010; Hu and Song, 2002; Hu., 2012; Jiang., 2013a, b; Jung., 2011; Li., 2010a, b, 2013; Liu., 2010; Lv., 2013; Paiva, 2012; Pan., 2012, 2013; Song., 2009; Vďačný., 2010; Wang., 2011; Xu., 2011). Among these groups, many taxa have been investigated in detail using either morphological or molecular methods (Berger, 1999, 2008, 2011; Huang., 2012; Yi and Song, 2011; Yi., 2009a, b, c, d, 2012).
The family Oxytrichidae is species-rich and morphologically diverse, in which nearly 200 valid morphospecies have been described so far (Berger, 1999, 2011; Gupta., 2001;Kamra., 2008; Qin., 2011; Singh., 2013; Song, 1990, 2004; Song and Wilbert 2002; Weisse., 2012; Wilbert and Song, 2008), ren- dering Oxytrichidae one of the largest families of Sporadotrichida.
Systematic revision of 24 Oxytrichidae genera had been provided by Berger (1999). However, in the subsequent studies, 13 more have been reported or re-estab- lished, namelyShao., 2011,Cohn, 1866,Jankowski, 1989,Gupta., 2006,Shao., 2013,Singh and Kamra, 2013,Li., 2010,Berger, 2011,Eigner, 1999,Küppers., 2011,Foissner., 2002,Foissner., 2002,Foissner., 2002 (Berger, 2008, 2011; Foissner., 2002; Gupta., 2003, 2006; Küppers., 2011; Li., 2010a; Shao., 2011, 2013a, b; Singh and Kamra, 2013; Song, 2001). The diagnoses ofBory De Saint-Vincent in Lamouroux, Bory De Saint-Vincent and Deslongchamps, 1824,Hemberger in Foissner, 1982,Stokes, 1887,Cohn, 1866 andNjine, 1979 were also revised (Shao., 2011, 2013a; Song, 2001).
The aims of this study included 1) clarifying the outline of the typical 18 frontal-ventral-transverse-cirri genera; 2) summarizing the patterns of development and determining the amount of variability during morphogenesis in oxytrichids, as well as the validity of classification schemes that are based on generic separation; 3) supplying the schematic illustrations as well as the key to typical 18 frontal-ventral-transverse-cirri genera of Oxytrichidae.
2 Oxytrichidae and Subfamily Classification
Ciliates of Oxytrichidae are mainly characterized by 18 frontal-ventral-transverse cirri clustering to six distinct groups which usually originate from six longitudinal primordia segregating 1, 3, 3, 3, 4, 4 cirri. They are also characterized by three primarily formed dorsal kineties, while fragmentation of kinety 3 presents but sometimes secondarily lost, and dorsomarginal kineties present but sometimes lost.
Based on the body flexibility, the existence of the cortical granules and the length of the adoral zone of membranelles, family Oxytrichidae was classified into two subfamilies, Oxytrichinae and Stylonychinae (Berger, 1999). Characterization of Oxytrichinae: body flexible; cortical granules present, secondarily lost several times; adoral zone of membranelles usually ≤40% of body length (Berger, 1999). Characterization of Stylonychinae: body rigid; cortical granules lacking; adoral zone of membranelles usually ≥40% of body length (Berger, 1999).
2.1 Some Morphological and Morphogenetic Patterns in Oxytrichidae
Based on the shape of the adoral zone of membranelles,andpatterns can be recognized. The former is formed like a question mark (Fig.1a), while the latter has the distal portion extending along the left margin of the cell, causing the proximal portion of the zone abruptly bending to the centre of the body (Berger, 1999; Fig.1d).
Based on the shape and arrangement of the paroral and endoral, six patterns can be classified.
pattern(Berger, 1999; Fig.1a): The paroral and endoral are both slightly curved and optically intersect at about the middle.
pattern (Berger, 1999; Fig.1b): The paroral and endoral are both slightly curved and optically intersect at about the middle, however, the paroral has a hooked distal end bending anteriorly and ventrally.
pattern (Berger, 1999; Fig.1c): The endoral is slightly curved, while the paroral is strongly curved with the anterior part bending like an archer’s bow. Both optically intersect at about the middle.
pattern (Berger, 1999; Fig.1d): The paroral and endoral are both straight and arranged in parallel, however the paroral consists of few, widely spaced cilia and extends far beyond the anterior end of the endoral.
pattern (Berger, 1999; Fig.1e): The paroral and endoral are both straight and arranged in parallel with about the same length.
pattern (Berger, 1999; Fig.1f): The paroral and endoral are both curved in the middle portion. The sinusoidal endoral is fragmented in the middle part and does not intersect with the paroral.
Based on the morphogenesis of the dorsal ciliature, six patterns can be recognized.
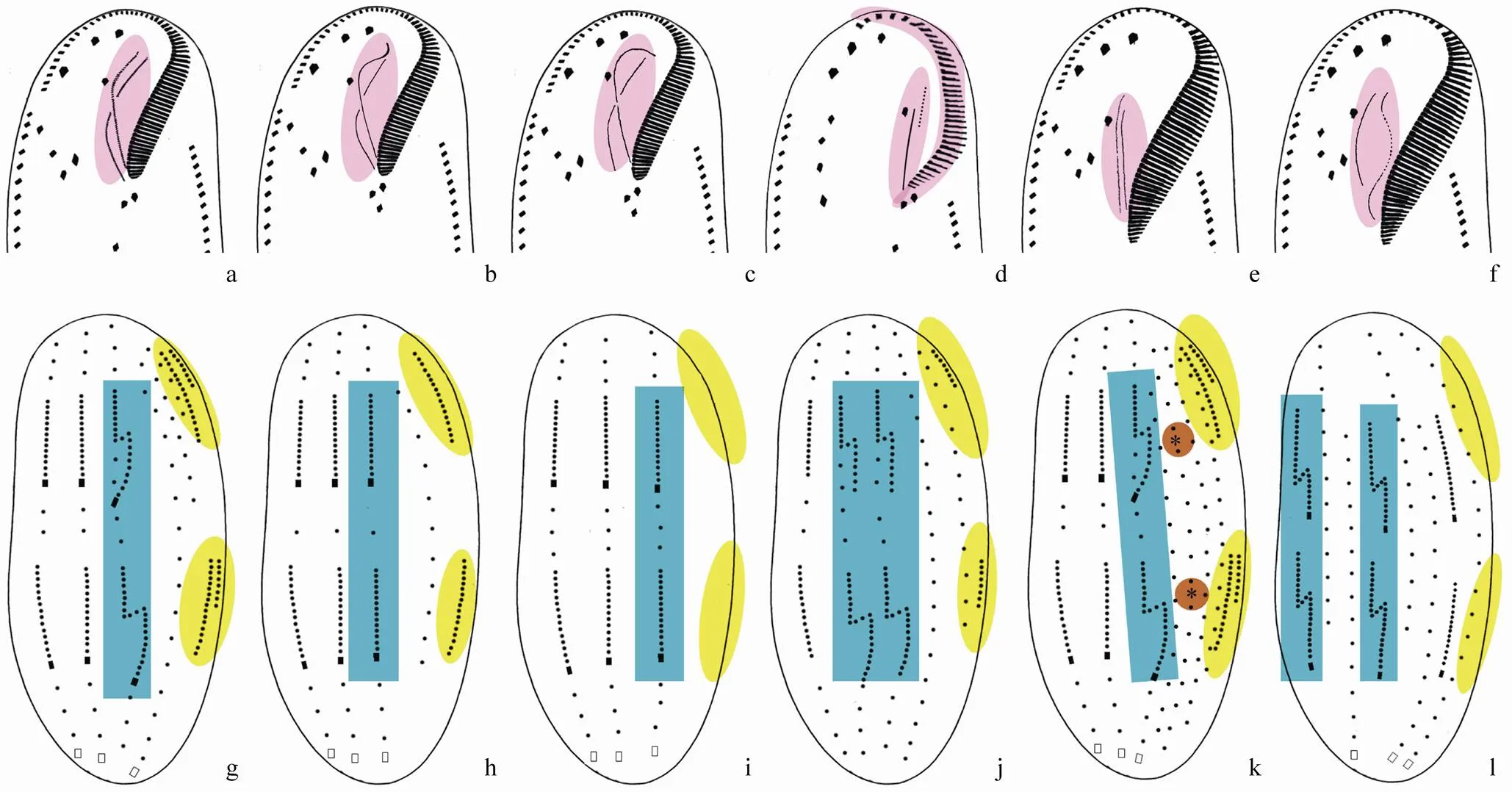
Fig.1 Schematic illustrations of shape of adoral zone of membranelles and arrangement of the paroral and endoral (a–f), as well as ontogenesis of the dorsal ciliature (g–l) in oxytrichids. (a) Oxytricha pattern; (b) Notohymena pattern; (c) Cyrtohymena pattern; (d) Gonostomum pattern; (e) Stylonychia pattern; (f) Steinia pattern; (g) Oxytricha pattern; (h) Urosomoida pattern; (i) Gonostomum pattern; (j) Tachysoma pattern; (k) Coniculostomum pattern. Asterisks show the parental dikineties which was retained during morphogenesis; (l) Hemigastrostyla pattern. Oral apparatus, dorsal kinety 3, dorsomarginal kineties and remaining of old dorsal kineties are shaded in violet, blue, yellow and green, respectively.
pattern (Berger, 1999): Kinety 3 fragments and dorsomarginal kineties are present (Fig.1g). Caudal cirri are present or absent. If present, usually three (sometimes more than three and arranged in three rows),one (or one row) each is present on kineties 1, 2, and 4. Apomorphy: Multiple fragmentation of kinety 3 is present or/and there are more than two dorsomarginal kineties (Fig.2c).
pattern (Berger, 1999): Fragmentation of kinety 3 is lost and dorsomarginal kineties are present (Fig.1h). Caudal cirri are present or absent. If present, usually three, one each is present on kineties 1–3.
pattern (Berger, 1999): Fragmentation of kinety 3 is lost and dorsomarginal kineties are absent (Fig.1i). Caudal cirri are present or absent. If present, usually three, one each is present on kineties 1–3.
pattern (Berger, 1999): Two dorsal kineties fragment and dorsomarginal kineties are present (Fig.1j). Apomorphy: Multiple fragmentation of dorsal kineties is present or/and there are more than two dorsomarginal kineties (Fig.4j).
pattern (Berger, 1999): Kinety 3 fragments and dorsomarginal kineties are present. Some parental kineties are retained during morphogenesis (Fig.1k). Caudal cirri are present or absent. If present, usually three,one each is present on kineties 1, 2 and 4.
pattern(Shao., 2011): Two dorsal kineties fragment and dorsomarginal kineties are absent (Fig.1l).
2.2 Characterization of Genera in the Subfamily Oxytrichinae
Bory De Saint-Vincent in Lamouroux, Bory De Saint-Vincent and Deslongchamps, 1824: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; frontoventral cirri in V-shaped pattern; one right and one left marginal row; five or six dorsal kineties formed inpattern; usually three caudal cirri (Berger, 1999; Shao., 2011; Fig.2a).
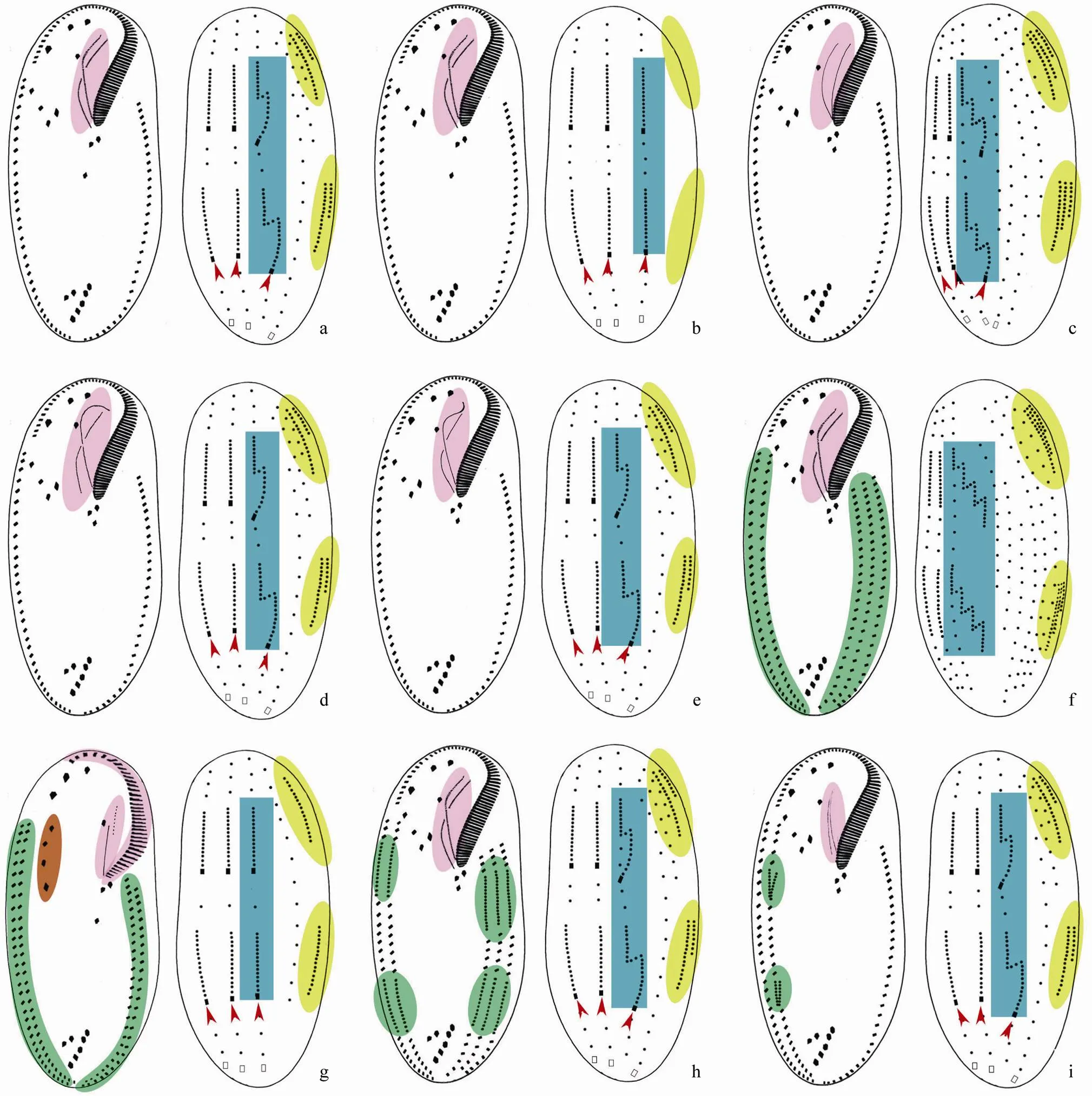
Fig.2 Diagram of the ventral and dorsal views of infraciliature and formation patterns of dorsal kineties anlagen and caudal cirri in Oxytrichidae s. l. genera. (a) Oxytricha; (b) Heterooxytricha; (c) Australocirrus; (d) Cyrtohymena; (e) Notohymena; (f) Ponturostyla; (g) Parurosoma; (h) Architricha; (i) Allotricha. Red arrows indicate caudal cirri. Oral apparatus, dorsal kinety 3, dorsomarginal kineties, multi-marginal rows/anlagen and some frontoventral cirri which are aligned in a row are shaded in violet, blue, yellow, green and brown, respectively.
Shao., 2011: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; frontoventral cirri in V- shaped pattern; one right and one left marginal row; three dorsal kineties formed inpattern; three caudal cirri (Shao., 2011; Fig.2b).
Blatterer and Foissner, 1988: adoral zone of membranelles formed like a question mark; endoral and paroral curved, not distinctly intersecting; frontoventral cirri in V-shaped pattern; one right and one left marginal row; more than six dorsal kineties formed in atypicalpattern, that is, more than two dorsomarginal kineties and kinety 3 with multiple fragmentation; caudal cirri present (Berger, 1999; Fig.2c).
Foissner, 1989: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; frontoventral cirri in V-shaped pattern; one right and one left marginal row; six dorsal kineties formed inpattern; three caudal cirri (Berger, 1999; Fig.2d).
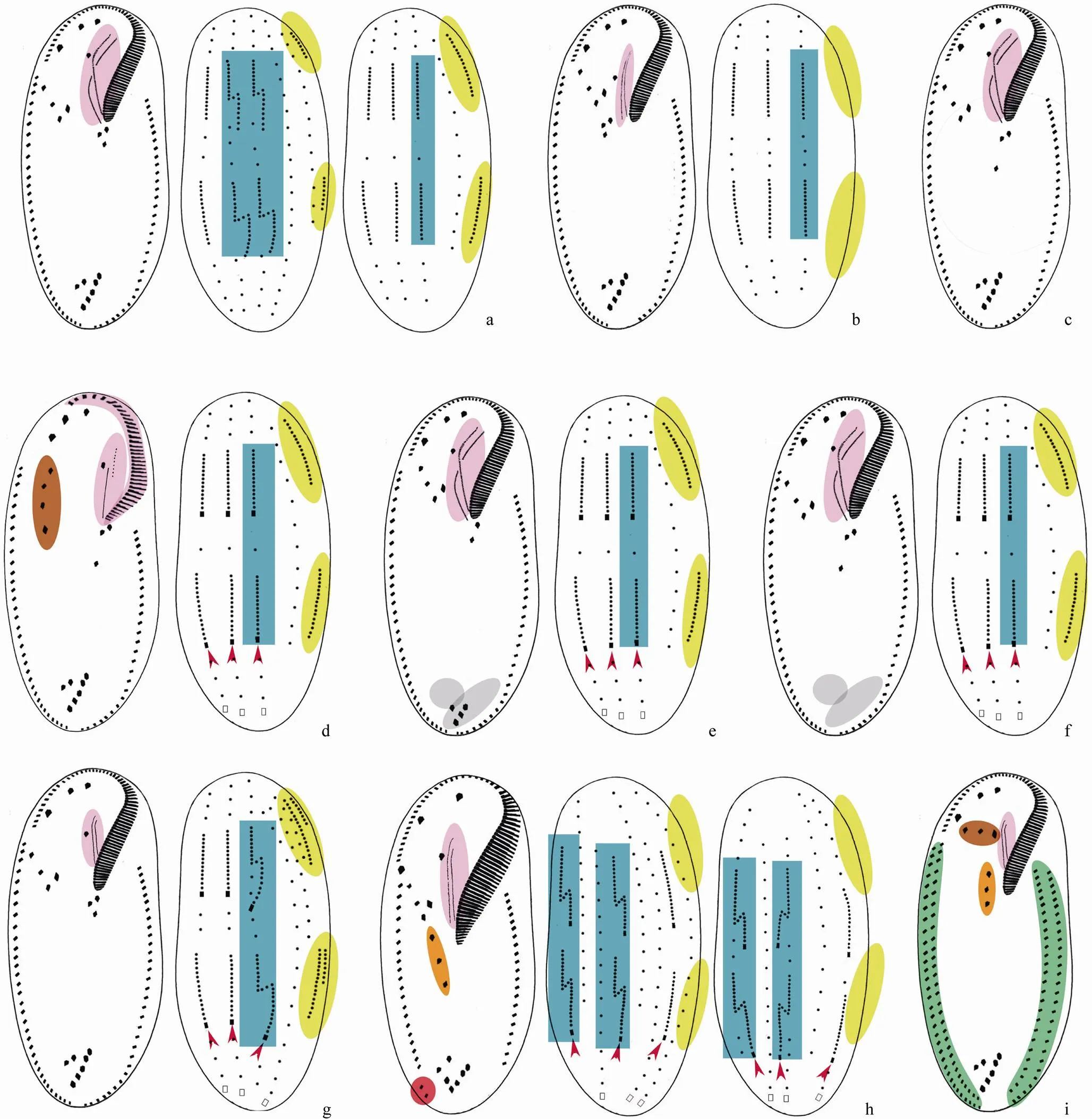
Fig.3 Diagram of the ventral and dorsal views of infraciliature and formation patterns of dorsal kineties anlagen and caudal cirri in Oxytrichidae s. l. genera. (a) Tachysoma; (b) Heterotachysoma; (c) Rubrioxytricha; (d) Urosoma; (e) Urosomoida; (f) Paraurosomoida; (g) Pseudostrombidium; (h) Hemigastrostyla; (i) Onychodromopsis. Red arrows indicate caudal cirri. Oral apparatus, dorsal kinety 3, dorsomarginal kineties, multi-marginal rows, pretransverse ventral and transverse cirri which are absent/reduced, extra cirri, some postoral ventral cirri which are aligned in a row and some frontoventral cirri which are aligned in a row are shaded in violet, blue, yellow, green, grey, red, orange and brown, respectively.
Blatterer and Foissner, 1988: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; frontoventral cirri in V-shaped pattern; one right and one left marginal row; six dorsal kineties formed inpattern; usually three, occasionally more than three caudal cirri, arranged in three rows (Berger, 1999; Fig.2e).
Jankowski, 1989: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; frontoventral cirri in V-shaped pattern; on each side, more than one marginal cirral row, which derived from a single anlage apokinetally; more than six dorsal kineties formed in atypicalpattern, that is, more than two dorsomarginal kineties and kinety 3 with multiple fragmentation; caudal cirri absent (Song, 2001; Fig.2f).
Gelei, 1954: adoral zone of membranelles, as well as endoral and paroral inpattern; frontoventral cirri arranged in a row; two right and two left marginal rows; four or five dorsal kineties formed very likely inpattern; caudal cirri present (Berger, 1999; Fig.2g).
Gupta., 2006: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; frontoventral cirri in V-shaped pattern; more than one marginal cirral row on each side and two anlagen develop separately within eachmarginal row; five or six dorsal kineties formed inpattern; three caudal cirri (Gupta., 2006; Fig.2h).
Sterki, 1878: adoral zone of membranelles formed like a question mark; endoral and paroral inorpattern; frontoventral cirri in V-shaped pattern; two or more right and one or more left marginal rows developed from a single anlage intrakinetally on each side; dorsal kineties formed inpattern; caudal cirri present (Berger, 1999; Petz and Foissner, 1996; Fig.2i).
Stokes, 1887: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; frontoventral cirri in V-shaped pattern or arranged in a row; one right and one left marginal row; four or six dorsal kineties formed in(intwo kineties (very likely) fragment) orpattern; caudal cirri absent (Shao., 2013a; Song and Warren, 1999; Fig.3a).
Shao., 2013: adoral zone of membranelles formed like an atypical question mark; endoral and paroral inorpattern; postoral ventral cirri to right of or behind buccal vertex; one right and one left marginal row; three dorsal kineties formed inpattern; caudal cirri absent (Shao., 2013a; Fig.3b).
Berger, 1999: cytoplasm usually distinctly orange to reddish or brownish; adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; frontoventral cirri in V- shaped pattern; one right and one left marginal row; four or five dorsal kineties formed very likely inpattern; one or two caudal cirri (Berger, 1999; Fig.3c).
Kowalewskiego, 1882: adoral zone of membranelles, as well as endoral and paroral inpattern; frontoventral cirri arranged in a row; one right and one left marginal row; four dorsal kineties formed inpattern; three caudal cirri(Berger, 1999; Fig.3d).
Hemberger in Foissner, 1982: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; frontoventral cirri in V-shaped pattern; number of postoral ventral cirri, and/or pretransverse ventral cirri, and/or transverse cirri usually reduced; one right and one left marginal row; four dorsal kineties formed inpattern; three caudal cirri(Shao., 2011; Fig.3e).
Singh and Kamra, 2013: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; frontoventral cirri in V-shaped pattern; pretransverse ventral and transverse cirri absent; one right and one left marginal row; four dorsal kineties formed inpattern; three caudal cirri(Singh and Kamra, 2013; Fig.3f).
Horváth, 1933: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; frontoventral cirri in V- shaped pattern; one right and one left marginal row; six dorsal kineties formed inpattern; more than four caudal cirri (Berger, 1999; Fig.3g).
Song and Wilbert, 1997: anterior end of the body more or less (narrowed) cephalized, adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; distal portion of adoral zone extending far posteriorly; 18–21 frontal-ven- tral-transverse cirri, some frontoventral and postoral ventral cirri form more or less distinct row; one right and one left marginal row; two ‘extra cirri’ present; four to six, usually five dorsal kineties formed inpattern; three caudal cirri (Shao., 2011; Fig.3h).
Li., 2010: anterior end of the body more or less (narrowed) cephalized, adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; distal portion of adoral zone extending far posteriorly; some frontoventral and the postoral ventral cirri form more or less distinct row; two ‘extra cirri’ present; three dorsal kineties formed inpattern; three caudal cirri(Li., 2010a; Fig.4a).
Stokes, 1887: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; frontoventral cirri in short transverse row or absent; postoral ventral cirri in longitudinal row right of buccal vertex; two or more right and two left marginal rows; dorsal kineties unknown; caudal cirri (very likely) present (Berger, 1999; Petz and Foissner, 1996; Fig.3i).
Foissner., 2002: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; five frontal-ventral-transverse cirral anlagen in total; frontoventral cirri in V-shaped pattern; number of postoral ventral and transverse cirri reduced; pretranverse ventral cirri absent; one right and one left marginal row; dorsal kineties formed inpattern; caudal cirri present (Berger, 2008; Foissner., 2002; Fig.4j).
Foissner., 2002: adoral zone of membranelles bipartite; endoral and paroral almost straight, short, and side by side; five frontal-ventral- transverse cirral anlagen in total; frontoventral cirri in V-shaped pattern; postoral ventral, pretransverse ventral, and transverse cirri absent; one left and one right marginal row; dorsal kineties formed inpattern; caudal cirri absent (Berger, 2008; Foissner., 2002; Fig.5e).
Foissner., 2002: adoral zone of membranelles, as well as endoral and paroral inpattern; five frontal-ventral-transverse cirral anlagen in total; frontoventral cirri arranged in a row; number of postoral ventral, pretransverse ventral, and transverse cirri reduced or groups even lacking; one right and one left marginal row; dorsal kineties formed inpattern; three caudal cirri (Berger, 2008; Foissner., 2002; Fig.5f).
Fig.4 Diagram of the ventral and dorsal views of infraciliature and formation patterns of dorsal kineties anlagen and caudal cirri in Oxytrichidaegenera. (a); (b); (c); (d); (e);(f);(g); (h); (i); (j). Red arrows indicate caudal cirri. Oral apparatus, dorsal kinety 3, dorsomarginal kineties, multi-marginal rows, pretransverse ventral and transverse cirri which are absent/reduced, extra cirri, some postoral ventral cirri which are aligned in a row/absent/reduced and the remaining of old dorsal kineties are shaded in violet, blue, yellow, green, grey, red, orange and brown, respectively.
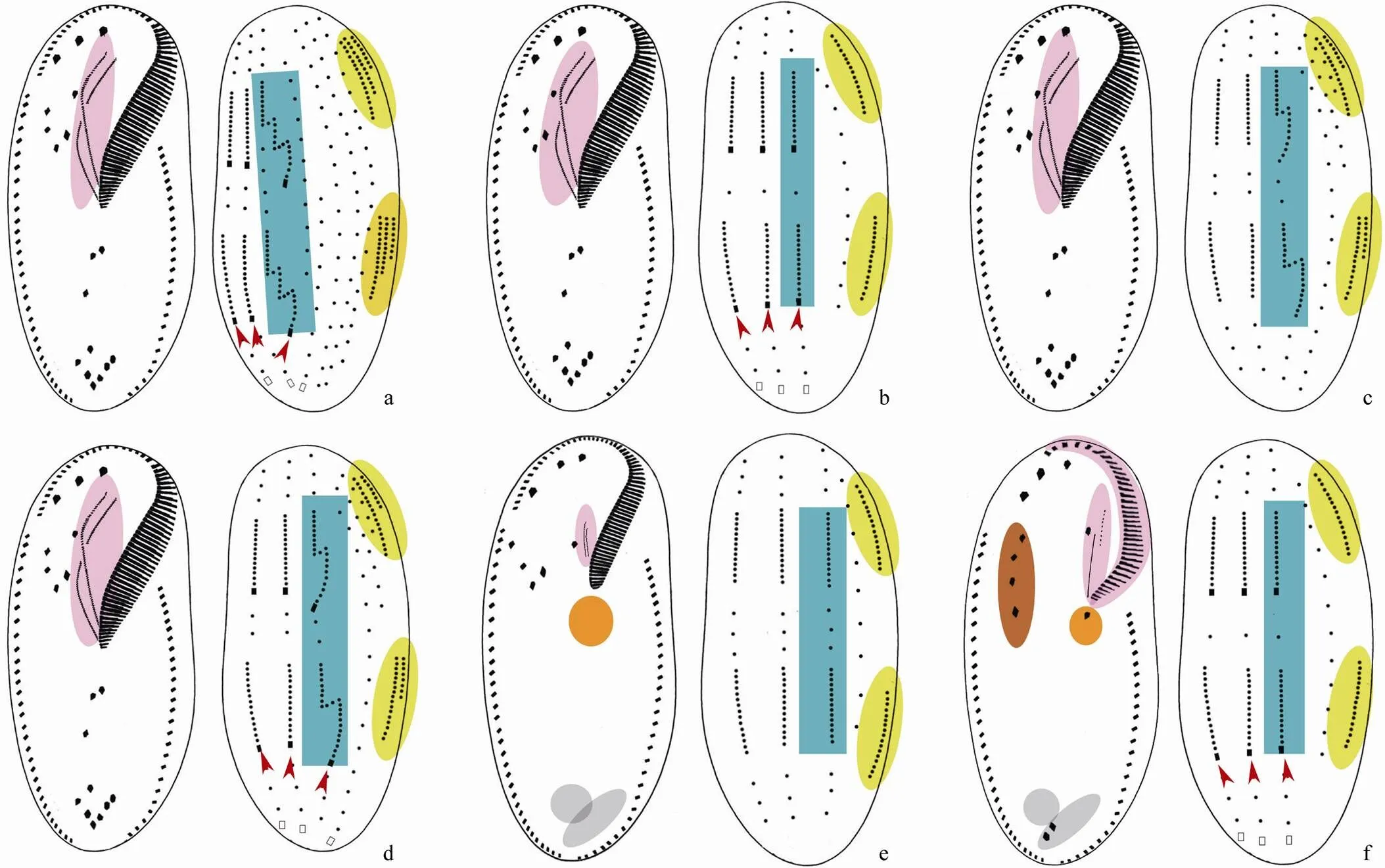
Fig.5 Diagram of the ventral and dorsal views of infraciliature and formation patterns of dorsal kineties anlagen and caudal cirri in Oxytrichidae s. l. genera. (a) Rigidocortex; (b) Parasterkiella; (c) Histriculus; (d) Sterkiella; (e)Vermioxytricha; (f) Hemiurosoma. Red arrows indicate caudal cirri. Oral apparatus, dorsal kinety 3, dorsomarginal kineties, pretransverse ventral and transverse cirri which are absent/reduced, some postoral ventral cirri which are aligned in a row and some frontoventral cirri which are aligned in a row are shaded in violet, blue, yellow, grey, orange and brown respectively.
2.3 Characterization of Genera in Subfamily Stylonychinae
Ehrenberg, 1830: body distinctly broader anteriorly than posteriorly, pellicle distinctly rigid; adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; transverse cirri separated into two groups; one right and one left marginal row; six dorsal kineties formed inpattern; three caudal cirri often distinctly elongated (Berger, 1999; Fig.4h).
Eigner, 1999: body elliptical to obovoid in shape, pellicle comparatively rigid; adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; transverse cirri arranged in J-shape; one right and one left marginal row; six dorsal kineties formed inpattern; three caudal cirri often distinctly elongated (Eigner, 1999; Shao., 2013; Fig.4i).
Cohn, 1866: adoral zone of membranelles bipartite, distal adoral membranelles widely spaced and radially arranged; endoral and paroral short and in parallel; buccal lip with distinct spine; one right and one left marginal row; more than three dorsal kineties, dorsal morphogenesis unknown; caudal cirri present (Shao., 2011; Fig.4d).
Dragesco, 1963: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; one right and one left marginal row; caudal cirri absent(Berger, 1999; Fig.4e).
Stein, 1859: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; two or more right and one or more left marginal rows, no parental marginal cirri retained after division; dorsal morphogenesis inpattern; three caudal cirri (Berger, 1999; Gupta., 2003; Fig.4f).
Njine, 1979: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; two or more right and one or more left marginal rows, parental marginal cirri partially retained after division; six or more dorsal kineties formed inpattern; caudal cirri present and usually distinctly elongated (Berger, 1999; Kamra., 1994; Fig.4g).
Berger, 2011: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; one right and one left marginal row; six dorsal kineties formed inpattern; three caudal cirri (Berger, 2011; Fig.4b).
Diesing, 1866: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern; frontoventral cirri in V-shaped pattern; one right and one left marginal row; six dorsal kineties formed inpattern; three caudal cirri(Berger, 1999; Voss and Foissner, 1996; Fig.4c).
Berger, 1999: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern;one right and one left marginal row; more than six dorsal kineties formed in atypicalpattern, that is, kinety 3 with multiple fragmentation and more than two dorsomarginal kineties; caudal cirri present (Berger, 1999; Fig.5a).
Küppers., 2011: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern;one right and one left marginal row; four dorsal kineties formed inpattern; caudal cirri present (Küppers., 2011; Fig.5b).
Corliss, 1960: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern;one right and one left marginal row; six dorsal kineties formed inpattern; caudal cirri absent (Berger, 1999; Fig.5c).
Foissner., 1991: adoral zone of membranelles formed like a question mark; endoral and paroral inpattern;one right and one left marginal row; six dorsal kineties formed inpattern; three caudal cirri(Berger, 1999; Fig.5d).
3 Keys to Typical 18 Frontal-Ventral-Transverse Cirri Genera of Oxytrichidae

1Body flexible…………………………………………………………..…………………………………………………………2 1.1Body rigid………………………...………………………….............…………………………………………………………23 2Anterior cell end generally cephalized, with two ‘extra cirri’………......………………………………………………………3 2.1Anterior cell not cephalized, no ‘extra cirri’………...........…………………………………………………………………4 3Dorsal kineties developed in Hemigastrostyla pattern……………..…………………………………………...Hemigastrostyla 3.1Dorsal kineties developed in Gonostomum pattern……………….……………………………………....………Apogastrostyla 4Paroral and endoral in Cyrtohymena or Notohymena pattern…………….………….................………….……………………5 4.1Paroral and endoral not in Cyrtohymena or Notohymena pattern….….………………......……………….……………………6 5Paroral and endoral in Cyrtohymena pattern………………………..........…….………………………..…………Cyrtohymena 5.1Paroral and endoral in Notohymena pattern…………………….…………………………...……………………....Notohymena 6More than one marginal row on either or both sides…………..……………………………………......……………………….7 6.1One marginal row on each side………………………………...………………………………………………………………11 7Frontoventral cirri in transverse row or absent………..………………………………………………………Onychodromopsis 7.1Frontoventral cirri in longitudinal row or in V-shaped pattern………………………………………………………………….8 8Caudal cirri absent……………………………...……………………………………………………………………Ponturostyla 8.1Caudal cirri present………………………………………………………………………………………………………………9 9Adoral zone formed like a question mark; dorsal kineties formed in Oxytricha pattern………………………………………10 9.1Adoral zone in Gonostomum pattern; dorsal kineties formed in Urosomoida pattern…………..………………......Parurosoma 10Two anlagen develop separately within each marginal row……………………...……………………………..........Architricha 10.1Groups of marginal cirri develop from a single anlage……….………………………………..………………………Allotricha 11Caudal cirri absent…………………………………………………………………………………………………………...…12 11.1Caudal cirri present………………………………………………………………………………………………………..……14 12Adoral zone of bipartite, less than ten frontal-ventral-transverse cirri........……...............………………………Vermioxytricha 12.1Adoral zone of continues, with ca. 18 frontal-ventral-transverse cirri..........………………………………………..................13 13Four or six dorsal kineties…………………………………………………………………..........…………………. .Tachysoma 13.1Three dorsal kineties…………………………………………………………..............………………......…...Heterotachysoma 14Cytoplasm orange to reddish or brownish………………………………………………………………………. .Rubrioxytricha 14.1Cytoplasm colourless………………………………………………………..………………………………………………….15 15Dorsal kineties form in Urosomoida pattern……………….…………………………………………………………………..16 15.1Dorsal kineties not form in Urosomoida pattern…………………………………………………………….…………………20 16Adoral zone in Gonostomum-pattern, frontoventral cirri in a single row……………………………………………………...17 16.1Adoral zone in non-Gonostomum-pattern, frontoventral cirri in V-shaped pattern…………………………....……………….18 17Five FVT-anlagen, less than ten frontal-ventral-transverse cirri...............……...........………………....………....Hemiurosoma 17.1Six FVT-anlagen, about 18 frontal-ventral-transverse cirri…………………….………………………………………Urosoma 18Five FVT-anlagen, forming less than ten frontal-ventral-transverse cirri...........…………..........……………….........Erimophry 18.1Six FVT-anlagen, forming ca. 18 frontal-ventral-transverse cirri.........………………….......……...........................................19 19Transverse and pretransverse ventral cirri present……………………….....…………………….............................Urosomoida 19.1Transverse and pretransverse ventral cirri absent………………….............………………...............................Paraurosomoida 20More than four caudal cirri…………………………………………….…………………….........................Pseudostrombidium 20.1Three caudal cirri or fewer……………………………....................................................………………..…………...….........21 21More than six dorsal kineties……………………………………………………………………………………...Australocirrus 21.1Six dorsal kineties or fewer………………………………………………………………………………………………….…22 22Dorsal kineties form in Gonostomum pattern………………………………………………...............................Heterooxytricha 22.1Dorsal kineties form in Oxytricha pattern……………………………………………………………...........................Oxytricha 23Paroral and endoral in Cyrtohymena pattern…………………………………………………................................Rigidohymena 23.1Paroral and endoral not in Cyrtohymena pattern…………………………………………………………………………….…24 24Paroral and endoral in Steinia pattern……………………………………………………………………………………...Steinia 24.1Paroral and endoral not in Steinia pattern………………………………………………………………………....……….....…25 25Adoral zone bipartite, buccal lip with distinct spine…………………………………………………………….......Actinotricha 25.1Adoral zone in continued structure, buccal lip not with distinct spine…………………………………………………...……26 26More than one marginal row on either or both sides……………………………………………………………………...........27 26.1One marginal row on each side……………………………………………………………………………………...................28 27Paroral and endoral in Oxytricha pattern, neither parental marginal cirri nor dorsal kinetiesretained after division……………………………………………………………………………………………….Pleurotricha 27.1Paroral and endoral in Stylonychia pattern, parental marginal cirri and dorsal kineties partiallyretained after division…………………………………………………………………………………………...Coniculostomum 28More than six dorsal kineties ………………………………………………………………………………….........Rigidocortex 28.1Six dorsal kineties or fewer……………………………………………………………………………………………….........29 29Paroral and endoral in Oxytricha pattern……………………………………………………………………………….............30 29.1Paroral and endoral in Stylonychia pattern……………………………………………………………………………………..32 30Six dorsal kineties………………………………………………………………………………………………………...........31 30.1Four dorsal kineties…………………………………………………………………………………………..........Parasterkiella 31Caudal cirri absent…………………………………………………………………………………………….............Histriculus 31.1Caudal cirri present……………………………………………………………………………………………………..Sterkiella 32Caudal cirri absent……………………………………………………………………………………….............Parastylonychia 32.1Caudal cirri present…………………………………………………………………………………………………………….33 33Transverse cirri not in two distinct groups……………………………………………………………………………Tetmemena 33.1Transverse cirri separated into two groups…………………………………………………………………………...Stylonychia
4 Summary of the Typical 18 Frontal- Ventral-Transverse-Cirri Oxytrichidae Genera
Above all, there is consensus among most taxonomists who are familiar with hypotrichs, and it is well recognized that the following characters are important for genera separation in Oxytrichidae (Berger, 1999, 2008, 2011; Berger and Foissner, 1997; Foissner., 2002; Gupta., 2003, 2006;Küppers., 2011; Li., 2010a; Shao., 2011, 2013a, b; Singh and Kamra, 2013; Song, 2001): 1) living morphology,., body shape, flexible/rigid, as well as cortical granules present/absent; 2) the pattern of the buccal ciliature,.. the undulating membranes and adoral zone of membranelles; 3) the pattern of the dorsal ciliature,.. the number and developmental pattern of dorsal kineties; 4) number and origin of marginal cirral rows; 5) caudal and extra cirri present/absent; and 6) the pattern of the ventral ciliature, in particular the number of frontal-ventral-transverse cirri and arrangement of frontoventral cirri.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (31372148) and the Fundamental Research Funds for the Central Universities.
Berger, H., 1999. Monograph of the Oxytrichidae (Ciliophora, Hypotrichia)., 78 (1): 1-1080.
Berger, H., 2008. Monograph of the Amphisiellidae and Trache- lostylidae (Ciliophora, Hypotricha)., 88 (1): 1-737.
Berger, H., 2011. Monograph of the Gonostomatidae and Ka- hliellidae (Ciliophora, Hypotricha).,90 (1): 1-741.
Berger, H., and Foissner, W., 1997. Cladistic relationships and generic characterization of oxytrichid hypotrichs (Protozoa, Ciliophora)., 148 (2): 125-155.
Chen, L. Y., Liu, W. W., Liu, A., Al-Farraj, S. A., Al-Rasheid, K. A. S., Warren, A., and Shao, C., 2013a. Morphology and molecular phylogeny of a new marine hypotrichous ciliate,n. sp. (Ciliophora, Hypotrichia).,60 (6): 588-600.
Chen, X. M., Hu, X. Z., Lin, X. F., Al-Rasheid, K. A. S., Ma, H. G., and Miao, M., 2013b. Morphology, ontogeny and molecular phylogeny of a new brackish water ciliatesp. n. (Ciliophora, Hypotricha) from southern China., 49 (4): 611-622.
Eigner, P., 1999. Comparison of divisional morphogenesis in four morphologically different clones of the genusand update of the natural hypotrich system (Ciliophora, Hypotrichida)., 35 (1): 34- 48.
Foissner, W., 2012.nov. gen., nov. spec., a secondarily oligomerized hypotrich (Ciliophora, Hypotricha, Schmidingerotrichidae nov. fam.) from hypersaline soils of Africa., 48 (3): 237-251.
Foissner, W., Agatha, S., and Berger, H., 2002. Soil ciliates (Protozoa, Ciliophora) from Namibia (Southwest Africa), with emphasis on two contrasting environments, the Etosha region and the Namib desert. Part I: Text and line drawings. Part II: Photographs., 5 (1): 1-1459.
Foissner, W., and Stoeck, T., 2011.nov. gen., nov. spec., a gonostomatid ciliate (Ciliophora, Hypo- tricha) from tank bromeliads (Bromeliaceae) with de novo originating dorsal kineties., 47 (1): 29-50.
Foissner, W., Shi, X. B., Wang, R. Q., and Warren, A., 2010. A reinvestigation ofpopulations, including the description ofnov. spec. (Ciliophora, Hypotricha)., 49 (2): 87-105.
Gupta, R., Kamra, K., and Sapra, G. R., 2006. Morphology and cell division of the oxytrichidsnov. gen., nov. sp., and(Müller, 1773), Corliss, 1960 (Ciliophora, Hypotrichida)., 42 (1): 29-48.
Gupta, R., Kamra, K., Arora, S., and Sapra, G. R., 2001.sp. n., a new oxytrichid (Ciliophora: Hypotrichida) ciliate from the river Yamuna, Delhi, India., 40 (1): 75-82.
Gupta, R., Kamra, K., Arora, S., and Sapra, G. R., 2003.(Shi, Warren and Song 2002) nov. comb. (Cili- ophora: Hypotrichida): morphology and ontogenesis of an Indian population; redefinition of the genus., 39 (3): 275-285.
Hu, X. Z., and Song, W. B., 2002. Studies on the ectocommen- sal ciliate,nov. spec. (Protozoa: Ciliophora: Hypotrichida) from the mantle cavity of the scallop.,481 (1): 173-179.
Hu, X. Z., Huang, J., and Warren, A., 2012. The morphology and phylogeny of two euplotid ciliates,spec. nov. andBorror, 1965 (Protozoa, Ciliophora, Euplotida)., 62 (11): 2757-2773.
Huang, J., Dunthorn, M., and Song, W. B., 2012. Expanding character sampling for the molecular phylogeny of euplotid ciliates (Protozoa, Ciliophora) using three markers, with a focus on the family Uronychiidae.,63 (3): 598-605.
Jiang, J. M., Huang, J., Li, L. Q., Shao, C., Al-Rasheid, K. A. S., Al-Farraj, S. A., and Chen, Z. G., 2013a. Morphology, ontogeny, and molecular phylogeny of two novel bakuellid-like hypotrichs (Ciliophora: Hypotricha), with establishment of two new genera., 49 (1): 78- 92.
Jiang, J. M., Xing, Y., Miao, M., Shao, C., Warren, A., and Song, W. B., 2013b. Two new marine ciliates,n. sp. andn. sp. (Ciliophora, Spirotrichea), with phylogenetic analyses inferred from the small-subunit rRNA gene sequences., 60 (4): 388-398.
Jung, J., Baek, Y., Sanghee, K., Choi, H., and Min, G., 2011. A new marine ciliate,nov. spec. (Ciliophora, Urostylida) from the Antarctic Ocean., 50 (4): 289-300.
Kamra, K., Kumar, S., and Sapra, G., 2008. Species ofand(Ciliophora, Hypotrichida, Oxytrichidae) from the valley of flowers, India, with descriptions ofsp nov,sp nov andsp nov., 48 (3): 372-388.
Kamra, K., Sapra, G. R., and Ammermann, D., 1994.n. sp., a new hypotrich ciliate: description and systematic relationships., 30 (1): 55-67.
Küppers, G. C., Paiva, T. S., Borges, B. N., Harada, M. L., Garraza, G. G., and Mataloni, G., 2011. An Antarctic hypotrich- ous ciliate,(Foissner) nov. gen., nov. comb., recorded in Argentinean peat-bogs: Morphology, morphogenesis, and molecular phylogeny., 47 (2): 103-123.
Li, F. C., Xing, Y., Li, J. M., Al-Rasheid, K. A. S., He, S. K., and Shao, C., 2013. Morphology, morphogenesis and small subunit rRNA gene sequence of a soil hypotrichous ciliate,(Ciliophora, Kahliellidae), from the shoreline of the Yellow River, north China., 60 (3): 247-256.
Li, L. Q., Huang, J., Song, W. B., Shin, M. K., Al-Rasheid, K. AS., and Berger, H., 2010a.(Kahl, 1932) gen. nov., comb. nov. (Ciliophora, Hypotricha): Morphology, notes on cell division, SSU rRNA gene sequence data, and neotypification., 49 (3): 195- 212.
Li, L. Q., Song, W. B., Al-Rasheid, K. A. S., Warren, A., Li, Z., Xu, Y., and Shao, C., 2010b. Morphology and morphogenesis of a new marine hypotrichous ciliate (Protozoa, Ciliophora, Pseudoamphisiellidae), including a report on the small subunit rRNA gene sequence., 158 (2): 231-243.
Liu, W. W., Shao, C., Gong, J., Li, J. M., Lin, X. F., and Song, W. B., 2010. Morphology, morphogenesis and molecular phylogeny of a new marine urostylid ciliate (Ciliophora, Stichotrichia) from the South China Sea, and an overview of the convergent evolution of midventral complex within the Spirotrichea.158 (4): 697-710.
Lv, Z., Chen, L. Y., Chen, L. Y., Shao, C., Miao, M., and Warren, A., 2013. Morphogenesis and molecular phylogeny of a new freshwater ciliate,n. sp. (Ciliophora, Oxytrichidae).,60 (5): 455-466.
Paiva, T. D. S., Borges B. D. N., Silva-Neto, I. D. D., and Harada, M. L., 2012. Morphology and 18S rDNA phylogeny of(Ciliophora, Hypotricha) from Brazil with redefinition of the genus., 62 (1): 229-241.
Pan, Y., Li, J. M., Li, L. F., Hu, X. Z., Al-Rasheid, K. A. S., and Warren, A., 2013. Ontogeny and molecular phylogeny of a new marine ciliate genus,g. n. (Protozoa, Ciliophora, Hypotricha), with description of a new species., 49 (2): 298-311.
Pan, Y., Li, L. Q., Shao, C., Hu, X. Z., Ma, H. G., Al-Rasheid, K. A. S., and Warren, A., 2012. Morphology and ontogenesis of a marine ciliate,(Dujardin, 1841) Kahl, 1932 (Ciliophora, Euplotida)., 51 (1): 29- 38.
Petz, W., and Foissner, W., 1996. Morphology and morphogenesis ofBerger and Foissner andStokes, two hypotrichs (Protozoa: Ciliophora) from Antarctic soils., 35 (2): 257- 280.
Qin, Y. H., Qiu, Z. J., Shao, C., Warren, A., and Shen, Z., 2011. Morphological redescription and morphogenesis of(Wrześniowski, 1866) Berger, 1999 (Ciliophora, Hypotrichida)., 50 (3): 163-174.
Shao, C., Ding, Y., Al-Rasheid, K. A. S., Al-Farraj, S. A., Warren, A., and Song, W. B., 2013a. Establishment of a new hypotrichous genus,n. gen. and notes on the morphogenesis of(Ciliophora, Hypotrichia).,49 (1): 93- 105.
Shao, C., Pan, X. M., Jiang, J. M., Ma, H. G., Al-Rasheid, K. AS., Warren, A., and Lin, X. F., 2013b.A redescription of the oxytrichid(Müller, 1786) Eigner, 1999 and notes on morphogenesis in the marine urostylidLei., 2005 (Ciliophora, Hypotri- chia)., 49 (2): 272-282.
Shao, C., Song, W. B., Al-Rasheid, K. A. S., and Berger, H., 2011. Redefinition and reassignment of the 18-cirri genera,,, and(Ciliophora, Hypotricha), and description of one new genus and two new species., 50 (4): 263-287.
Singh, J., and Kamra, K., 2013.gen. nov., sp. nov., an oxytrichid (Ciliophora, Hypotricha) from Kyongnosla Alpine Sanctuary, including note on non-oxytri- chid Dorsomarginalia., 49 (4): 600-610.
Singh, J., Kamra, K. and Sapra, G. R., 2013. Morphology, ontogenesis, and molecular phylogeny of an Indian population of(), including remarks on the subgenus.,49 (2): 283- 297.
Song, W. B., 1990. A comparative analysis of the morphology and morphogenesis of(Engelmann, 1862) (Ciliophora, Hypotrichida) and related species., 37 (3): 249-257.
Song, W. B., 2001. Morphology and morphogenesis of the marine ciliate(Dragesco & Dragesco- Kernéis, 1986) Jankowski, 1989 (Ciliophora, Hypotrichida, Oxytrichidae)., 37 (2): 181- 197.
Song, W. B., 2004. Morphogenesis ofa (Ciliophoa, Hypotrichida, Oxytrichidae) during binary fission., 40 (3): 245-254.
Song, W. B., and Warren, A., 1999. Observations on morpho- genesis in a marine ciliate(Protozoa: Cili- ophora: Hypotrichida)., 79 (1): 35-38.
Song, W. B., and Wilbert, N., 1997. Morphological investigations on some free living ciliates (Protozoa, Ciliophora) from China seas with description of a new hypotrichous genus,nov. gen., 148 (4): 413-444.
Song, W. B., and Wilbert, N., 2002. Faunistic studies on marine ciliates from the Antarctic benthic area, including descriptions of one epizoic from, 6 new species and 2 new genera (Protozoa: Ciliophora).41 (1): 23-61.
Song, W. B., Warren, A., and Hu, X. Z., 2009.. Science Press, Beijing, 518pp.
Song, W. B., Wilbert, N., Li, L. Q., and Zhang, Q. Q., 2011. Re-evaluation on the diversity of the polyphyletic genus(Ciliophora, Hypotricha): ontogenetic, mor- phologic, and molecular data suggest the establishment of a new genusnov. gen., 58 (1):11-21.
Vďačný, P., Tirjaková, E., Tóthová, T., Pristaš, P., and Havorský, P., 2010. Morphological and phylogenetical studies on a new soil hypotrich ciliate:spec. nov. (Hypotrichia, Kahliellidae)., 46 (4): 319- 333.
Voss, H. J., and Foissner, W., 1996. Divisional morphogenesis in(Ciliophora, Hypotrichida): A compara- tive light and scanning electron microscopic study., 32 (1): 31-46.
Wang, Y. G., Hu, X., Z., Al-Rasheid, K. A. S., and Warren, A., 2011. Two urostylid ciliates,spec. nov. and(Lin & Song, 2004) Xu., 2006 (Ciliophora, Stichotrichia), from a,63 (3): 1740-1750.
Weisse, T., Moser, M., Scheffel, U., Stadler, P., Berendonk, T., Weithoff, G., and Berger, H., 2012. Systematics and species-specific response to pH ofsp. nov. andsp. (Ciliophora, Hypotricha) from acid mining lakes.,49 (2):255- 271.
Wilbert, N., and Song, W. B., 2008. A further study on littoral ciliates (Protozoa, Ciliophora) near King George Island, Antarctica, with description of a new genus and seven new species., 42 (13-14): 979-1012.
Xu, Y., Huang, J., Hu, X. Z., Al-Rasheid, K. A. S., and Song, W. B., 2011. Taxonomy, ontogeny and molecular phylogeny ofspec. nov. (Ciliophora, Hypotri- chida) from the Yellow Sea, China.,61 (8): 2000-2014.
Yi, Z. Z., and Song, W. B., 2011. Evolution of the order Urostylida (Protozoa, Ciliophora): New hypotheses based on multi-gene information and identification of localized incongruence., 6 (3):e17471.
Yi, Z. Z., Clamp, J. C., Al-Rasheid, K. A. S., Al-Khedhairy, A. A., Chen, Z. G., and Song, W. B., 2009a. Evolutionary relationship and species separation of four morphologically similar stichotrichous ciliates (Protozoa, Ciliophora) inferred from PCR-RFLP patterns, ITS2 region sequence, and ITS2 secondary structures., 19 (5): 581-586.
Yi, Z. Z., Katz, L., and Song, W. B., 2012. Assessing whether alpha-tubulin sequences are suitable for phylogenetic reconstruction of Ciliophora with insights into its evolution in euplotids.,7 (7): e40635.
Yi, Z. Z., Song, W. B., Clamp, J. C., Chen, Z. G.., Gao, S., and Zhang, Q. Q., 2009b. Reconsideration of systematic relationships within the order Euplotida (Protista, Ciliophora) using new sequences of the gene coding for small-subunit rRNA and testing the use of combined data sets to construct phylogenies of the-complex., 50 (3): 599-607.
Yi, Z. Z., Song, W. B., Gong, J., Warren, A., Al-Rasheid, K. A. S., Al-Arifi, S., and Al-Khedhairy, A. A., 2009c. Phylogeny of six oligohymenophoreans (Protozoa, Ciliophora) inferred from small subunit rRNA gene sequences.,38 (3): 323-331.
Yi, Z. Z., Song, W. B., Stoeck, T., Al-Rasheid K. A. S., Al-Khedhairy, A. A., Gong, J., Ma, H. G., and Chen, Z. G., 2009d. Phylogenetic analyses suggest that(Ciliophora, Urostylida) should represent an urostylid family, based on small subunit rRNA and alpha-tubulin gene sequence information.,157 (2): 227-236.
(Edited by Qiu Yantao)
10.1007/s11802-015-2482-7
(September 11, 2013; revised October 15, 2013; accepted July 10, 2014)
. Tel: 0086-29-82668463 Ext 423 E-mail: shaochen@mail.xjtu.edu.cn
ISSN 1672-5182, 2015 14 (3): 522-532
© Ocean University of China, Science Press and Spring-Verlag Berlin Heidelberg 2015
 Journal of Ocean University of China2015年3期
Journal of Ocean University of China2015年3期
- Journal of Ocean University of China的其它文章
- Monte Carlo Simulation of in situ Gamma-Spectra Recorded by NaI (Tl) Detector in the Marine Environment
- Effects of Waterborne Cu and Cd on Anti-oxidative Response,Lipid Peroxidation and Heavy Metals Accumulation in Abalone Haliotis discus hannai Ino
- Application of a Delay-Difference Model for the Stock Assessment of Southern Atlantic Albacore (Thunnus alalunga)
- Invasion and Morphological Variation of the Non- Indigenous Barnacle Chthamalus challengeri (Hoek, 1883) in Yangshan Port and its Surrounding Areas
- The Burial of Biogenic Silica, Organic Carbon and Organic Nitrogen in the Sediments of the East China Sea
- Hydrochemistry of the Natural Low pH Groundwater in the Coastal Aquifers near Beihai, China