
瓢鸡SLC24A5基因多态性与肤色性状的关联分析*
2015-03-29 02:09杨向东谷合勇史宪伟
家畜生态学报 2015年7期
李 杨,杨向东,谷合勇,王 静,张 斌,史宪伟*
(1.云南农业大学动物科学与技术学院云南省动物营养与饲料重点实验室,昆明云南650201;2.昆明云岭广大种禽饲料有限公司,云南昆明650215;3.昆明学院农学院,云南昆明650214)
鸡的色素性状,包括羽色、肤色、胫色、喙色、冠色、耳叶和内脏组织等颜色,不仅是品种的重要特征,也与经济价值密切相关。鸡肤色主要有白肤色、黄肤色和黑肤色3种类型。黄肤色与黄类胡萝卜素在皮肤的沉积有关,主要受β-胡罗卜素双氧酶2(BCDO2)基因变异和表达影响[1]。黑皮肤的表现与真黑色素与褐黑素在皮肤的沉积量与比例相关[2]。到目前为止,控制皮肤黑色素的主效基因还未见报道。
SLC24A5(solute carrier family 24,member 5)是溶质载体24家族的一个成员,是依赖于钾离子的钠/钙阳离子交换体[3]。自Lamason等[4]揭示斑马鱼的金色表型是由SLC24A5基因突变以来,该基因与不同种族人群肤色的关系一直是研究热点。SLC24A5基因第3外显子的1个功能SNP是不同人种肤色变化的主要原因[5],同时,该基因已作为人类系统发育的1个重要祖先信息标记[6]。鸡SLC24A5基因位于10号染色体,编码423个氨基酸。罗娟等[7]研究了SLC24A5基因在白来航鸡、乌骨鸡和北京油鸡不同组织的表达量,发现乌骨鸡皮肤、肌肉及眼睛中SLC24A5基因表达量均显著高于非乌骨鸡[7]。本课题组已研究了SLC24A5基因多态性与他留乌骨鸡羽色和肤色性状的相关性,发现SLC24A5基因变异与肤色性状呈显著相关[8]。
瓢鸡是中国“十一五”期间畜禽品种资源调查发现的特有珍稀品种,中心产区为云南普洱市镇沅县,无尾性状是该品种的重要特性,其特征为无尾椎骨、尾棕骨、尾脂腺,无主尾羽、镰羽[9-10]。贡潘偏抽等分析了瓢鸡mtDNA D-loop序列,揭示瓢鸡具有较高的遗传多样性和较复杂的母系来源[11]。除无尾性状为该品种的典型特征外,黑肤(乌质)性状也是该品种的一个重要特点。自然群体虽以黑肤瓢鸡为主,也存在一定数量的白肤瓢鸡。为了揭示瓢鸡黑肤和白肤性状的遗传差异,本研究采用错配PCRRFLP技术研究云南瓢鸡SLC24A5基因第4外显子多态性,分析其与肤色性状的关联,以期为云南地方鸡种羽色肤色选择寻找有效的遗传标记,为中国地方鸡种的保种利用提供相应的分子遗传学依据。
1 材料与方法
1.1 样品来源
本研究96只瓢鸡样品取自云南省镇沅县瓢鸡保种场,其中白肤色鸡52只,黑肤色鸡44只。采样时记录每只鸡的性别、年龄、肤色及家系来源,受试鸡只彼此无血缘关系。利用肝素钠抗凝真空采血管从翅静脉采集全血3~5mL,4℃运至实验室,-20℃保存。
1.2 基因组DNA提取与检测
每只鸡取全血5~10μL,采用蛋白酶K与酚/氯仿标准提取法提取鸡基因组DNA。提取的DNA样品用1%凝胶电泳检测,TE缓冲液稀释备用。
1.3 引物设计
根据鸡SLC24A5全基因序列(NCBI登录号:NW_003763854.1),利用primer3(V.0.4.0)在线引物设计软件,设计1对引物扩增瓢鸡SLC24A5基因第3至第5外显子区域,引物序列为:正向引物序列为(SLC24A5-F1)5'-AGGCTACATTTGCAACGGTTTA-3';反向引物序列为(SLC24A5-R1)5'-GCTATGCCAAAACAGGTCAGAT-3',扩增片段长度为1 121bp。
根据SLC24A5基因C482GSNP所在位置,重新设计一对错配PCR-RFLP检测引物,引入Nhe I酶切识别位点,用于建立鸡SLC24A5基因错配Nhe I PCR-RFLP遗传标记。引物的正向序列(SLC24A5-Nhe I F):5'TGCAGCATGCGGGCTGCTCGCTA3';反向序列(SLC24A5-Nhe I R):5'TCTAAACAGCGGCCAACAGGATA3',扩增片段产物为389bp。
1.4 PCR扩增及测序
鸡SLC24A5基因的PCR扩增总体积为50 μL,包含5.0μL 10×buffers,1μL dNTPs(2.5 mmol),1.5μL正向和反向引物(10μmol),0.5μL Taq酶(10U/μL),4μL模板DNA(约50ng),36.5μL ddH2O。PCR反应条件为94℃预变性3 min;94℃变性45s,58℃退火45s,72℃延伸90 s,40个循环;72℃延伸8min;4℃保存。
从白肤色和黑肤色两个群体各随机选取6个个体进行PCR扩增与测序,检测SLC24A5变异位点。利用正反两个引物双向测序,测序公司为中美泰和生物科技有限公司(北京,中国)。
1.5 SLC24A5基因NheI PCR-RFLP分子标记的建立
鸡SLC24A5基因Nhe I PCR-RFLP检测技术采用滕召纯等[8]建立的方法。
1.6 统计分析
利用BioEdit(2.0)软件查看序列质量,DNAS- tar(5.0)软件包SeqMan程序查看变异位点,MegAlign程序进行序列比对。采用Excel软件计算基因频率、基因型频率、多态信息含量(PIC)与基因杂合度(He),利用SPSS(19.0)统计软件进行基因型与肤色χ2独立性检验。
2 结果与分析
2.1 鸡SLC24A5基因PCR产物扩增结果
以瓢鸡血液基因组DNA为模板,用SLC24A5-F1/R1引物扩增,扩增区域为SLC24A5基因第3至第5外显子区域,长度为1 121bp。PCR扩增得到与预期片段大小一致的产物,扩增效果良好,可用于后续的DNA测序和RFLP分析。
2.2 基因测序与SNP检测
从白肤色和黑肤色鸡群随机各选取6只瓢鸡进行个体测序,NCBI Blast同源性比较,所测序列与GenBank的SLC24A5基因序列(基因登录号:NW_003763854.1)的同源性达98%以上,证明扩增的产物为瓢鸡SLC24A5基因。瓢鸡个体间序列比对后发现,在第4外显子编码区出现变异,该变异在482位核苷酸由G变为C(c.G 482C),为异义突变,引起161位苏氨酸变为丝氨酸(T161S)。图1是G 482C位点SNP所在位置的测序峰图。
2.3 瓢鸡SLC24A5基因错配Nhe I PCR-RFLP基因型检测
采用错配RFLP技术,新设计的引物(SLC24A5 Nhe I F/SLC24A5 Nhe I R)引入1个Nhe I酶切位点,建立SLC24A5基因错配Nhe I PCR-RFLP标记。PCR扩增瓢鸡DNA,得到1条389bp片段,经NheI酶切后,出现3种基因型,GG型为363bp与26bp两个片段,CC型为389bp 1个片段,GC型为389bp,363bp与26bp 3个片段。SLC24A5基因错配Nhe I PCR-RFLP检测电泳图见图2。
2.4 多态信息含量(PIC)与群体杂合度(He)
根据基因型频率与基因频率(表1),计算了PIC和He两个基因多态性指标,瓢鸡SLC24A5基因SNP的PIC为0.328,He为0.4160,均处于中度多态程度,表明瓢鸡在肤色性状方面受到自然选择和人工选择的影响较小。
2.5 SLC24A5基因G482C与肤色的相关性分析
利用错配Nhe I PCR-RFLP标记对瓢鸡SLC24A5基因进行个体基因分型,基因型频率、基因频率及χ2独立性检验结果见表1。从表1可知,G等位基因在白肤色群体和黑肤色群体均为优势基因,而黑肤色群体C等位基因频率(0.372)略高于白肤色(0.24)。两个群体基因型间χ2独立性检验达到显著水平(P=0.023)。统计分析表明,SLC24A5基因G482C位点与瓢鸡肤色具有一定的相关性。

图1 瓢鸡SLC24A5基因G482C位点SNP测序峰图Fig.1 Sequencing image of SLC24A5gene G482C SNP in Piao chicken
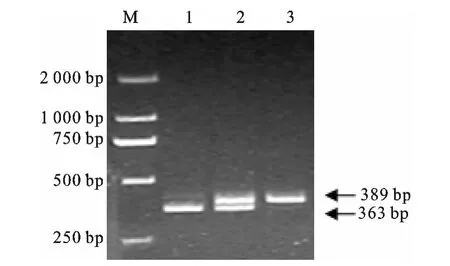
图2 瓢鸡SLC14A5基因Nhe I PCR-RFLP电泳图M.DGL2000分子量标记,1.GG基因型;2.GC基因型;3.CC基因型Fig.2 NheI PCR-RFLP image of SLC24A5gene in Piao chickenM.DGL 2000molecular marker;1.GG genotype;2.GC genotype;3.CC genotype
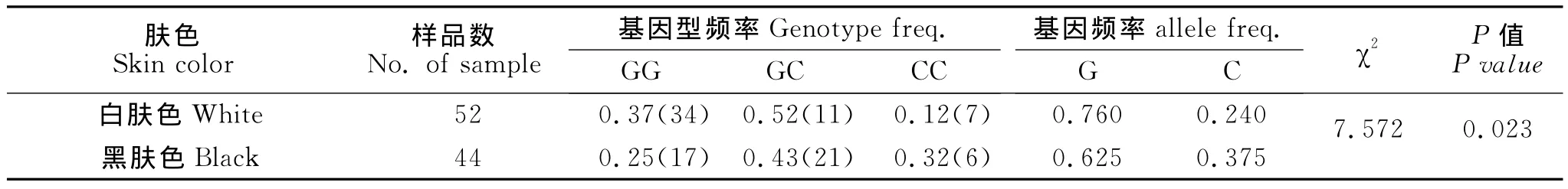
表1 瓢鸡SLC24A5基因G482C与肤色性状相关分析表Table 1 Association between SLC24A5G482Cgenotypes and skin color in Piao chicken
3 讨 论
本研究采用错配PCR-RFLP技术研究了SLC24A5基因外显子4的1个功能SNP与鸡肤色性状的相关性。该SNP序列(CTGCTCTCTA[C/G]CGTGGTAAGA)不能被现有的限制性内切酶所识别,常规的RFLP技术无法检测该位点SNP基因型。本文在SNP位点前方新设计了1个引物(SLC24A5-Nhe I F:5'TGCAGCATGCGGGCTGCTCGCTA3'),改变了引物3'第4位碱基,由原来的碱基T变为碱基G,这样扩增的PCR产物就引入了Nhe I酶切位点(G^CTAGC),扩增产物既能被内切酶消化产生不同基因型片段,又不改变原有的SNP序列。该技术对没有内切酶识别位点而又有重要功能的单个SNP检测,是1种花费不高且快速简便的方法。
鸡肤色主要有白色、黄色和黑色3种类型,白皮肤属于祖先型,对黄皮肤和黑皮肤均为显性(W*W),黄皮肤为隐性(W*Y),黑皮肤为不完全显性(W*B)[12]。到目前为止,已揭示BCDO2基因是控制黄皮肤的主效基因[1],而黑肤色的主效基因还未见报道。有人比较MC1R基因、SLC24A5基因和TYR基因在他留乌骨鸡不同羽色和肤色性状间的遗传差异,结果显示,MC1R基因和TYR基因与羽色有一定关系,而SLC24A5基因与肤色(乌质性状)性状显著相关[8,13]。本文对SLC24A5基因G482C在瓢鸡两个不同肤色群体的研究表明,虽然基因型与肤色的相关性在两个品种中都达到显著水平,但基因型分布和等位基因频率却相差较大。如他留乌骨鸡黑肤色(乌质)群体中G基因频率为0.62,与瓢鸡黑肤色群体 G基因频率很接近(0.63),但在非乌骨群体中,他留乌骨鸡的C等位基因为0.73,而在瓢鸡则为0.24。显然,除SLC24A5基因之外,还会有其他基因也参与鸡皮肤色素的沉积与扩散。另外,SLC24A5基因在不同鸡种基因频率的差异可能不仅与肤色性状有关,还与鸡种的遗传背景以及基因纯合度有关。
[1] Eriksson J,Larson G,Gunnarsson U.Identification of the yellow skin gene reveals a hybrid origin of the domestic chicken[J].PLoS Genet,2008,4(2):e1000010.
[2] Chedekel M R,Subbarao K V,Bhan P,et al.Biosynthetic and structural studies on pheomelanin[J].Biochim Biophys Acta,1987,12(2):239-243.
[3] Quillen E E,Shriver M D.SLC24A5:exchanging genetic and biochemical knowledge[J].Pigment Cell Melanoma Res,2008, 21(3):344-345.
[4] Lamason R L,Mohideen M A,Mest J R,et al.SLC24A5,a putative cation exchanger,affects pigmentation in zebrafish and humans[J].Science,2005,310(5755):1 782-1 786.
[5] Stokowski R P,Pant P V K,Dadd T,et al.A genomewide association study of skin pigmentation in a south asian population[J].Am J Hum Genet,2007,81(6):1 119-1 132.
[6] Giardina E,Pietrangeli I,Martinez-Labarga C.Haplotypes in SLC24A5gene as ancestry informative markers in different populations[J].Current Genomics,2008,9(2):110-114.
[7] 罗 娟,张 炜,于 冰,等.鸡SLC24A5基因的cDNA克隆、表达分析及其与黑色素沉积的关系研究[J].生物化学与生物物理进展,2008,35(1):69-76.
[8] 滕召纯,孙利民,陆晓屏,等.他留乌骨鸡SLC24A5基因Nhe I mismatch PCR-RFLP标记与羽色及肤色相关性研究[J].中国家禽,2014,36(15):6-9.
[9] 朱图寿,谢强明,李才勇,等.瓢鸡保护与利用的研究进展[J].中国畜牧兽医,2011,38(5):250-252.
[10] 宋 迟,韩 威,胡 艳,等.无尾瓢鸡尾部骨骼缺失观察[J].中国家禽,2014,36(18):53-56.
[11] 贡潘偏抽,刘丽仙,李大林,等.基于线粒体DNA控制区(mtDNA D-loop)序列分析瓢鸡的遗传多样性[J].云南农业大学学报:自然科学版,2011,26(2):211-214.
[12] Smyth J R.Genetic of plumage,skin and eye pigmentation in chickens[M]∥Crawford R D.Poultry Breeding and genetics.Amsterdam:Elsevier,1990:109-167.
[13] 滕召纯,陆哓屏,王玉祥,等.MC1R基因TaqⅠPCR-RFLP标记与他留乌骨鸡羽色性状的相关性研究[J].中国家禽,2013,35(20):11-14.
猜你喜欢
中国畜禽种业(2020年7期)2020-12-16
疯狂英语·新悦读(2020年4期)2020-06-18
好孩子画报(2020年3期)2020-05-14
小天使·四年级语数英综合(2019年9期)2019-11-09
江苏农业科学(2018年21期)2018-12-06
中国畜牧兽医文摘(2016年7期)2016-01-30
现代检验医学杂志(2015年6期)2015-02-06
实验动物与比较医学(2014年5期)2014-02-28
华东理工大学学报(自然科学版)(2014年3期)2014-02-27
中国糖料(2013年1期)2013-01-22
