
钙调蛋白和钙/钙调蛋白依赖性蛋白激酶Ⅱ在大鼠海马发育中的变化规律
2015-03-25 02:49:15王少博牟心红付正英张引国
武警医学 2015年1期
王少博,牟心红,孟 斌,付正英,张引国
钙调蛋白和钙/钙调蛋白依赖性蛋白激酶Ⅱ在大鼠海马发育中的变化规律
王少博1,牟心红2,孟 斌3,付正英4,张引国5
目的 探讨钙调蛋白(calmodulin, CaM)、钙/钙调蛋白依赖性激酶Ⅱ(Ca2+/calmodulin kinaseⅡ, CaMKⅡ)在大鼠海马发育中的变化规律。方法 以出生后第1、7、14、21、30和 50天的大鼠为研究对象, 采用实时定量PCR和Western blot方法分别检测海马CaM和CaMKⅡ的mRNA及蛋白的表达。结果 大鼠海马CaM及CaMKⅡ在出生后第1天时表达较弱,第14~30天均快速增加(P<0.05或P<0.01),呈增量表达;第50天后表达持续在高水平(CaMmRNA和CaMKⅡmRNA相对值分别为2.242±0.154和2.418±0.329;CaM蛋白和CaMKⅡ蛋白相对值分别为0.782±0.061和2.194±0.256;P<0.05或P<0.01),但较出生后第21天已有减弱趋势。结论 CaM和CaMKⅡ在海马中的表达变化具有一定的时序性,此模式可能与突触的发育及脑成熟变化密切相关,也与学习记忆功能的形成密切相关。
钙调蛋白;钙/钙调蛋白依赖性激酶Ⅱ;海马;认知
中枢神经系统在发育过程中,无论是突触数目还是神经回路的模式都是时刻变化的。与此同时,作为信号传递基本单位的突触,其传递效能也不是固定不变的。学习和记忆是脑的重要功能,一般认为其神经基础是突触的可塑性及新神经回路的形成。海马是位于大脑底部边缘系统的结构,是调节学习和记忆的核心区域[1]。长时程增强(long term potentiation,LTP)和长时程抑制(long term depression,LTD)是海马突触可塑性的两种主要表现形式。
CaM是广泛存在于细胞内的一种信号系统调控蛋白,也是细胞内最普遍存在的Ca2+感受器。它通过与多种蛋白的相互作用,调控着突触小泡的发生、运输及再填充,从而传递胞内Ca2+浓度变化的信号,对神经递质的释放及突触电生理活动起到至关重要的调控作用[2 -5]。
CaMKⅡ位于突触后致密体(postsynaptic density,PSD)中,为PSD的主要成分之一,在海马中表达丰富,占脑内总蛋白的0.5%~1.0%及海马蛋白的2%。 CaMKⅡ在神经递质的合成与释放、骨架蛋白磷酸化、突触可塑性、基因表达和脑内葡萄糖代谢等方面有着重要的生物学作用,是神经信息传导通路中的重要因子[2 -5]。
本实验通过研究不同发育时期大鼠海马中CaM、CaMKⅡ的mRNA和蛋白表达量,旨在研究发育中突触可塑性的形成,为认知障碍相关疾病的治疗提供理论依据。
1 材料与方法
1.1 动物与取材 选用成年Wistar大鼠[军事医学科学院动物所提供,许可证号:SCXK-(军)2012-0004], 体重270~290 g。雌鼠与雄鼠适应3~4 d后,按照3∶1比例合笼。以次晨发现阴栓或阴道分泌物镜检发现精子者确定为妊娠第0天。取孕鼠6只,待分娩后取出生后第1、7、14、21、30、50 天 的仔鼠断头处死(从每窝中随机取 1 只,每组内6只鼠)。在冰上取出海马, 分别用于 RNA 和蛋白质的提取。
1.2 主要试剂 CaM兔单克隆抗体(Abcam 公司)、CaMKⅡ兔单克隆抗体(Epitomics公司)、HRP羊抗兔IgG 抗体(Abgent公司)、RNA提取试剂盒 (D9086)、反转录试剂盒 (DRR014A)、DNA Marker (DL500)和SYBR Green试剂盒(RR820A,TaKaRa公司)、琼脂糖(Spain公司)、PAFR蛋白裂解液(北京普利莱基因技术有限公司)。引物序列见表1。

表1 CaM、 CaMKⅡ 和β-actin基因的引物序列
1.3 Real-time PCR法定量检测CaM和 CaMKⅡ mRNA水平 根据Trizol试剂盒说明匀浆海马组织并提取总RNA;依照反转录试盒说明书,取 RNA 溶液在反转录酶作用下反转录成 cDNA。然后,以cDNA为模板进行实时定量荧光检测,结果采用 2-△△Ct进行分析。PCR引物由北京鼎国昌盛技术生物技术有限公司合成。
1.4 Western blot法检测CaM和CaMKⅡ蛋白的表达 将组织加入含1%蛋白酶抑制药的1 ml蛋白裂解液中,匀浆后冰上静置30 min。12000 r/min 4 ℃离心10 min取上清。采用BCA法进行蛋白定量,根据上样量计算上样体积,100 ℃蛋白变性8 min备用。制备12% SDS-PAGE凝胶,微量加样器顺序加样后电泳至条带到达目的蛋白相应区域。以电压25 V电流2.5 mA,CaM转膜9 min,CaMKⅡ 转膜13 min,8%奶粉37 ℃封闭孵育1 h,加入一抗37 ℃ 封闭1 h,4 ℃过夜。TBST洗膜加二抗37 ℃孵育1 h。
洗膜后取ECL试剂A液、B液等体积混合加在膜上3~5 min,暗室胶片曝光、显影、定影,扫描胶片并保存图片,Image J软件分析条带光密度值,将目的条带与β-actin的光密度比值作为目的蛋白的相对量。
1.5 统计学处理 所得数据输入 SPSS 统计软件进行单因素方差分析,以P<0.05 表示组间差异具有显著性。
2 结 果
2.1 CaMmRNA及CaMKⅡmRNA在大鼠海马发育中的变化 由表2可见,出生后第1天的 CaM mRNA相对表达量较低,出生后第7~50天的CaM mRNA相对表达量明显增高,差异具有统计学意义(均P<0.01)。CaMKⅡmRNA在出生后第1~7天相对表达量较低,出生后第14~50 天相对表达量明显增高,差异具有统计学意义(P<0.05或P<0.01)。
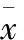
表2 CaMmRNA、 CaMKⅡmRNA在大鼠海马发育中的变化 (n=6;±s)
注:与第1天组比较,①P<0.05, ②P<0.01
2.2 CaM蛋白及CaMKⅡ蛋白在海马发育中的表达 使用Western blot方法揭示各组CaM蛋白和CaMKⅡ蛋白的表达。由表3可见,CaM蛋白在出生后第1天相对表达量较低,出生后第7~50天的相对表达量明显增高,差异具有统计学意义(P<0.05或P<0.01)。CaMKⅡ蛋白在出生后第1~7天相对表达量较低,出生后第14~50天相对表达量明显增高 (P<0.01)。
表3 CaM蛋白、 CaMKⅡ蛋白在大鼠海马发育中的变化 (n=6;±s)
注:与第1天组比较,①P<0.05, ②P<0.01
3 讨 论
活动依赖的突触可塑性发育存在一个关键期,其调控分子和细胞机制的核心是细胞内钙水平的变化。也就是说,启动活动依赖的突触可塑性的各种信号,包括神经递质及其他神经营养因子等,最终都会影响细胞内的Ca2+水平。细胞内Ca2+增加可以激活一系列激酶,从而导致细胞骨架的修饰以及神经元树突的形态改变,并且通过基因表达来体现功能改变[6]。
钙调蛋白是一种在所有真核生物体内均有表达的识别Ca2+的信号蛋白,在脑内主要分布在海马、纹状体和大脑皮质,大约占脑蛋白的0.5%。CaM存在于神经元的许多亚细胞成分中,在胞浆液、线粒体、微粒体和突触泡浆等部分都有分布。钙调蛋白浓度与脑的正常功能有着重要的联系[7]。它能与Ca2+结合,将胞内Ca2+浓度变化的信号传递给多种依赖Ca2+而发挥生物学功能的蛋白。CaMKⅡ占PSD组分蛋白总量的20%~30%,是一种钙依赖性蛋白激酶,在海马突触中分布密集。CaMKⅡ是神经系统特异表达的蛋白,并且具有发育阶段特异性,即只在生后表达。Silva等[8]研究证明当用基因打靶技术敲除小鼠的CaMKⅡ亚基后海马和新皮层的细胞形态和NMDA受体通道功能都正常,但在海马脑片上不能够诱导出LTP。由此也说明CaMKⅡ在突触可塑性中起着重要作用。
本实验对大鼠海马CaM、CaMKⅡ的发育研究发现, 在出生后第1天,无论是基因水平还是蛋白水平表达均较弱;出生后第14~30天,其表达明显增强;在出生后第50天,其基因和蛋白水平表达均持续在高水平。由于LTP的产生,主要是由突触后Ca2+浓度的升高,大量Ca2+可作为第二信使进一步激活胞内的生化级联反应,导致突触的可塑性变化。内流的Ca2+与CaM形成Ca2+/CaM复合物,从而使CaMKⅡ激活,导致AMPA受体磷酸化增加;Ca2+/CaM还可以激活腺苷酸环化酶使PKA活化,最终使突触后膜上AMPA受体数量增加。活化的CaMKⅡ还能激活Ras鸟苷酸交换因子活化Ras-ERK通路,促进LTP的启动。因此推测CaM、CaMKⅡ出生后的变化可能与突触的发育和成熟有关。以往的研究表明,脑的发育与突触的形成之间存在着某种平行关系。在脑发育关键期(胎儿期和生后早期),神经元形成大量新突触连接,使脑中突触的数量最终达到成熟脑的两倍[9,10]。这一关键期对于人类而言是从妊娠后3个月至出生后2年,而啮齿类动物则为胚胎后期至出生后3~4周。在脑快速发育期之后,突触联系的减少是机体对早期产生的过多的突触进行修剪[11,12],以适应脑的成熟和神经网络的构建过程。在本研究中,CaM、CaMKⅡ出生后第1天达较弱,可能与海马突触发育尚不成熟有关;CaM、CaMKⅡ出生后第14~30天 的表达明显增强,显然正反映了海马的快速发育和突触联系增多过程。出生后第50天其基因和蛋白水平表达处于较高水平,但较出生后第21天已有所减弱趋势,说明随着脑发育成熟,机体对多余的突触进行修剪。因此,CaM和CaMKⅡ在海马中的特异性表达与突触的发育及成熟变化密切相关。
总之,CaM和CaMKⅡ的表达与神经系统突触可塑性的发育和成熟密切相关,同时也与学习和记忆功能密切相关,因而对CaM和CaMKⅡ在神经系统中发育规律的研究,可为认知功能障碍相关疾病的治疗提供理论依据。
[1] McLachlan R S. A brief review of the anatomy and physiology of the limbic system [J]. Can J Neurol Sci, 2009, 36(Suppl 2):S84-87.
[2] Mizuno K, Giese K P. Hippocampus-dependent memory formation: do memory type-specific mechanisms exist[J]. J Pharm acol Sci , 2005,98(3):191-197.
[3] LinK F, Chang R C, Suen K C. Modulation of calcium/calmodulin in kinase-Ⅱprovides partial neuroprotection against beta-amyloid peptide toxicity [J]. Eur J Neurosci, 2004,19(8):2047 -2055.
[4] Shonesy B C, Jalan-Sakrikar N, Cavener V S,etal. CaMKII: a molecular substrate for synaptic plasticity and memory[J]. Prog Mol Biol Transl Sci, 2014,122:61-87.
[5] Hell J W. CaMKⅡ: claiming center stage in postsynaptic function and organization[J]. Neuron, 2014,81(2):249-265.
[6] Coultrap S J, Bayer K U. CaMKⅡregulation in information processing and storage[J]. Trends N eurosci,2012,35(10):607-618.
[7] 董先平,智 刚,徐天乐. 钙调素参与离子通道和受体功能的调控[J]. 自然科学进展,2002,12(3):232-239.
[8] Silva A J, Paylor R, Wehner J M,etal. Imparied spatial learning in alphe-calcium-calmodulin kinaseⅡ mufant mice [J]. Science, 1992,257:206-211.
[9] Kalkman C J, Peelen L, Moons K G,etal. Behavior and development in children and age at the time of first anesthetic exposure [J]. Anesthesiology, 2009,110(4):805-812.
[10] Sanders R D, Xu J, Shu Y,etal. Dexmedetomidine attenuates isoflurane-induced neurocognitive impairment in neonatal rats [J]. Anesthesiology, 2009,110(5):1077-1085.
[11] Chia-Chien Chen, Ju Lu, Yi Zuo. Spatiotemporal dynamics of dendritic spines in the living brain [J]. Front Neuroanat, 2014,8:28-32.
[12] Uesaka N, Uchigashima M, Mikuni T,etal. Retrograde semaphorin signaling regulates synapse elimination in the developing mouse brain[J]. Science, 2014,344(6187):1020-1023.
(2014-08-12收稿 2014-11-15修回)
(责任编辑 梁秋野)
敬告:请作者投稿时务必在论文中注明所引用的参考文献,否则不予刊用。每篇论文的参考文献数量应为6条以上,并尽量引用近5年内发表的参考文献,切勿引用未发表的论文、内部资料、未汇编出版的会议论文及个人通信等。
论著要写出中英文摘要及关键词,其他文章只列出关键词,置于正文之前。一律采用结构式摘要,即包括目的、方法、结果和结论四部分。中文摘要200字左右,英文摘要与中文摘要相对应。关键词3~8个,应按中国医学科学院医学情报研究所编译出版的《医学主题词注释字顺表》规范使用,中医药文章参照《中医药主题词表》。中国图书分类号请根据文章内容的学科分类从《中国图书馆分类法》中查得。
Expression of Calmodulin and Ca2+/calmodulin kinaseⅡ in rat hippocampus during development stage
WANG Shaobo1, MOU Xinhong2, MENG Bin3, FU Zhengying4, and ZHANG Yinguo5.
1.Tianjin Uninversity of Traditional Chinese Medicine, Tianjin 300193, China; 2.Department of Basic Experimental Technology, 3.Department of Health, 4.Department of Gynaecology and Obstetrics of Affiliated Hospital, 5.Department of Physiology and Pathophysiology, Logistics University of Chinese People’s Armed Police Forces, Tianjin 300309,China
Objective To study the changes in expressions of CaM、CaMKⅡmRNA and protein in the hippocampus of rat during postnatal development. Methods Real-time PCR and Western blot were used to detect the different expressions of mRNA and protein levels of CaM,CaMKⅡ,respectively, in postnatal day 1, day 7, day 14, day 21, day 30 and day 50. Results The expression of CaM and CaMKⅡwere weaker in the first day after birth, but increased rapidly at postnatal days 14-30 (P<0.05 orP<0.01). The expressions continued to a higher level at postnatal day 50 (the relative values of CaMmRNA and CaMKⅡmRNA were 2.242±0.154 and 2.418±0.329, respectively; the relative values of CaM protein and CaMKⅡprotein were 0.782±0.061 and 2.194±0.256, respectively)(P<0.05 orP<0.01), but had low expression trend compared with postnatal day 21. Conclusions There is certain time sequence in the expressions of CaM/CaMKⅡmRNA and protein in the hippocampus of rat during postnatal development. This pattern may be closely related to the change of synaptic development and brain maturity. It is also closely related to the formation of learning and memory.
calmodulin;Ca2+/calmodulin kinaseⅡ; hippocampus;cognition
国家自然科学基金(81271224); 武警后勤学院创新团队基金 (WHTD 201308)
王少博,硕士研究生在读, E-mail:shaobo03786@163.com
1. 天津 300193, 天津中医药大学; 2.基础实验技术教研室, 3.卫生教研室, 4.附属医院妇产科, 5.生理学与病理生理学教研室,天津300309, 武警后勤学院
张引国, E-mail: yinguozh@sohu.com
R388.1
猜你喜欢
作文周刊·小学二年级版(2022年20期)2022-05-05 01:33:06
昆明医科大学学报(2022年1期)2022-02-28 07:43:34
昆明医科大学学报(2021年8期)2021-08-13 08:59:20
昆明医科大学学报(2021年6期)2021-07-31 07:40:08
创新作文(小学版)(2019年10期)2019-09-25 08:12:28
中华家教(2018年7期)2018-08-01 06:32:38
小学生学习指导(低年级)(2017年5期)2017-05-04 04:14:38
文学少年(原创儿童文学)(2016年16期)2016-02-28 17:50:19
西南医科大学学报(2015年1期)2015-08-22 13:01:48
作文与考试·小学高年级版(2015年17期)2015-05-30 10:48:04
