
杨梅开花生物学特性
2015-03-23 08:56:44沈利芬项伟波范彩廷周明兵徐川梅
浙江农林大学学报 2015年2期
沈利芬,项伟波,范彩廷,金 鹏,周明兵,徐川梅
杨梅开花生物学特性
沈利芬,项伟波,范彩廷,金 鹏,周明兵,徐川梅
(浙江农林大学 亚热带森林培育国家重点实验室培育基地,浙江 临安 311300)
为了系统了解杨梅Myrica rubra开花生物学特性,以杨梅品种东魁Myrica rubra ‘Dongkui’雌株和雄株为材料,体视显微镜下观察其雌雄花的外部形态,采用石蜡切片、荧光及曙红染色等技术,研究其花粉活力及内部发育结构。结果表明:杨梅为风媒花,柔荑花序。雄花中每朵小花具有8~20枚花药,每枚花药具有4个花粉囊,花粉母细胞减数分裂过程中,细胞质分裂为同时型,四分体主要以四面体型为主,少量为连续型。成熟花粉粒为二细胞型,具3个萌发孔,刚散粉的花粉活力达到90%以上。杨梅雌花序中的小花主要由柱头、花柱和子房等3部分组成,单心皮,1室,子房上位,直生胚珠,单珠被,厚珠心。图4参30
经济林学;杨梅;花药;子房;生物学特性
杨梅Myrica rubra属于杨梅科Myricaceae杨梅属Myrica常绿果树,雌雄异株,为中国特有的珍贵水果,主要分布在长江流域以南各省,以浙江省最多[1]。杨梅风味独特,营养价值和经济价值高,生产成本低,已成为中国南方山区农民增收的重要经济来源[2]。同时,杨梅适应性强,能在水土流失严重,土层瘠薄地带生长,且四季常绿,是不可多得的生态型经济树种[3]。近年来,杨梅栽培面积不断增加,但是优良的品种十分匮乏,因此开展杨梅基础研究具有重要的现实意义。目前,有关杨梅的基础研究比较少,主要集中在组织培养技术摸索[4]、内源激素测定[5]、育种技术探讨[1,6-7]、种质资源调查及鉴定[8]、遗传多样性分析[9-12]、气孔结构及花粉粒形态观察等方面[13-15]。在杨梅细胞学方面仅有少量研究。李三玉等[16]曾在20世纪80年代初对杨梅前期花芽分化进行初步研究。近30 a后,Sogo等[17]对杨梅的受精过程进行了细胞学分析,之后,有关杨梅细胞生物学方面的研究几乎未见报道。本研究从杨梅雌雄花序外部特征及发育动态、内部解剖结构及花粉活力等方面对其开花生物学特性进行了详细的研究,期望为杨梅的遗传育种研究提供理论依据,并为其生殖生物学研究增添新的数据。
1 材料与方法
1.1 材料
研究所用的杨梅雌株和杨梅雄株均为品种东魁Myrica rubra ‘Dongkui’,试样采集于浙江农林大学植物园。
1.2 方法
1.2.1 杨梅雌雄花序内外部特征观察 2013年3月1至4月20日,隔2 d采集1次杨梅的雌雄花序,体视显微镜下解剖其结构,并观察其内外部特征。
1.2.2 杨梅雌雄花序内部微观结构研究 隔2 d采集1次杨梅的雌雄花序,甲醛-乙醇-醋酸混合固定液(FAA)固定,之后进行常规石蜡切片[18]及曙红染色。
1.2.3 杨梅花粉萌发活力测定 采用荧光染色法(FCR,fluorochrome reaction),参照Sato等[19]的方法略有改进。从即将散粉的花药中收集花粉,隔24 h测定1次花粉活力,花粉活力测定过程中,设置3个重复,荧光显微镜下观察并统计花粉活力。
2 结果与分析
2.1 杨梅雄花生物学特征
2.1.1 杨梅雄花序特征 杨梅雄花序为柔荑花序,长1~3 cm(图1-1),小花着生在主穗轴上,无花冠和花萼,小花主要由8~20枚花药组成,花药外面有2~3个苞片包裹(图1-2~4)。雄花序从花药形成到成熟散粉期间,其花药颜色依次经历暗红色、黄红色、鲜红色的变化。花药变成鲜红色后(图1-5),花丝伸长,花药进一步散开外露,开始散粉,通常是位于花序中下部位的小花先散粉(图1-6),随后其他部位的小花散粉。散粉过程中,花药二叉开裂,花粉较多(图1-7~8),散粉后的花药干枯脱落。

图1 杨梅雄花序结构及散粉特性Figure 1 Structure and pollination characteristics of Myrica rubra male flowers
2.1.2 杨梅雄花序显微结构及发育 杨梅花药具有4个花粉囊,中部具有药隔(图2-1),花粉囊壁具有纤维层、中层和绒毡层3层(图2-2)。3月上旬,花药中的多数花粉母细胞处于减数第1次分裂前期(图2-2),3月中旬,则处于四分体发育时期,大部分四分体以四面体型存在(图2-3,淡色箭头所示),少数排列在一个平面上以十字交叉型存在(图2-3黑色箭头所示),故杨梅花粉母细胞胞质分裂主要以同时型为主,少量为连续型。3月中旬到4月初,由四分体分离出来的小孢子经过一系列的发育过程发育成熟,成熟花粉粒具有2个细胞核和3个萌发孔(图2-4)。
2.1.3 杨梅花粉活力 荧光染料本身不产生荧光,无极性,可以自由地透过完整的原生质膜,进入原生质后,与原生质内的酯酶作用形成一种能产生荧光的极性物质,荧光越强,表明花粉活力越强[20]。荧光染色结果表明:刚散粉的杨梅花粉染色活力非常强,在90%以上(图2-5),室温条件下放置24 h后,对其活力影响不大,48 h之后,其花粉活力急剧下降,仅有约69%的花粉具有染色活力,72 h之后,约52%的花粉具有染色活力, 96 h之后,花粉荧光非常弱,几乎不具有染色活力。
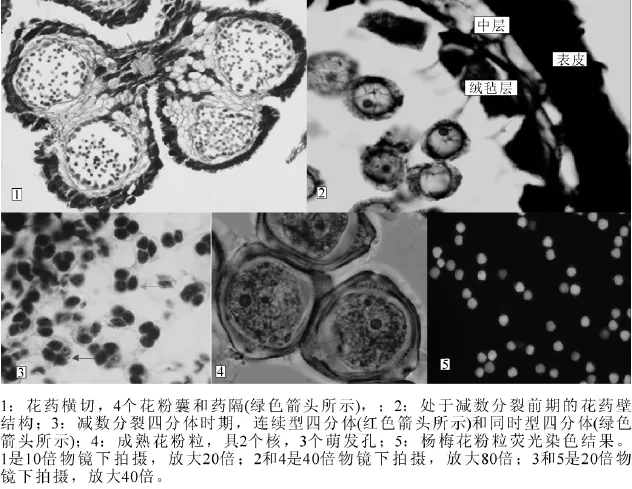
图2 杨梅雄花序内部结构Figure 2 Internal structure of Myrica rubra male flower
2.2 杨梅雌花生物学特征
2.2.1 杨梅雌花序特征及发育 杨梅雌花与雄花类似,为柔荑花序,长0.5~1.5 cm(图3-1),小花着生在同一花序轴上,主要由柱头、花柱和子房3个主要部分组成,花柱极短,柱头两裂,呈 “Y”状,小花外面有2~4个苞片包被(图3-7~10)。3月初,杨梅的雌花仍由苞片包被,整个雌花序成绿色(图3-2)。3月中下旬,杨梅雌花苞片展开,柱头开始外露,通常位于花序中上部的小花柱头最先显现(图3-3),随后,其他部位的柱头陆续外露展开,外露的柱头呈 “Y”状张开不断向外生长,最后完全展开(图3-4~9)。与雄花序相类似,杨梅雌花序在发育过程中,其柱头颜色依次经历着淡红色—鲜红色—红色—深红色的变化过程,外部的苞片也经历着嫩绿色—黄绿色—中间绿色边缘微红色—绿色等不同程度的颜色变化过程。
2.2.2 杨梅雌花内部微观结构 杨梅子房为单心皮,1室,其底部与花托相连,其余部分均独立,属于上位子房(图4-1),胚珠各方向生长速度均匀一致,珠孔、合点和珠柄三者在一条直线上,在胚珠类型上属于直生胚珠,单珠被(图4-2~4)。珠被在发育过程中并非把珠心全部包住,而在珠心顶部留1个小孔,即珠孔(图4-5)。杨梅大孢子母细胞形成时,在大孢子母细胞外层有4~5层细胞包裹,因此在珠心类型上,杨梅属于厚珠心型(图4-6)。
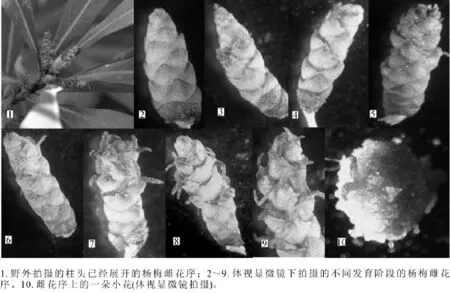
图3 杨梅雌花序结构及发育Figure 3 Structure and development of pistillate inflorescence in Myrica rubra

图4 杨梅子房及胚珠结构Figure 4 Ovary and ovule structure of Myrica rubra
3 小结与讨论
根据植物花粉传播媒介的不同,可以将植物分为风媒植物和虫媒植物,风媒植物通常雌雄异花或异株,花被小甚至无花被,无香味,浅色泽,花序密集成穗状或柔荑状,花粉小、量大、质量轻,含水量少[21]。杨梅开花时雌花序与雄花序均为上中部先开放,然后从花序顶端向基部渐次开放,雌花序基部发育差。本研究结果表明:杨梅的雌、雄花均为柔荑花序,无花冠和花萼,雄花无香味和蜜腺,具有数枚花药(每朵小花中有8~20枚花药)这与李三玉等[16]的研究结果相似,同时花粉量十分充足,这种结构特征表明,在传粉生物学上杨梅属于典型的风媒花。
植物雌雄蕊开放过程中的动态变化特征与植物的育性是密切相关的,是植物长期适应生态环境变化,保障其自身在自然条件下繁育成功的进化结果[22]。通过对花芽分化和形成及结构特征的研究,掌握其生长特性和规律,对指导杨梅高效栽培管理,促进其优质高产,解决大小年结果等问题具有重大意义。杨梅雌雄异株。雄花序单独或数条丛生于叶腋,有锥形、棒形、塔形和圆柱形等4类[23],通常不分枝呈单穗状,每苞片腋内生1雄花。雄花具2~4枚卵形小苞片及4~6枚雄蕊;花药椭圆形,暗红色,无毛。雌花序常单生于叶腋,较雄花序短且细瘦,苞片和雄花的苞片相似,密接而成覆瓦状排列,每苞片腋内生1雌花。子房卵形,光滑且极小,顶端有短花柱及2个鲜红色的细长柱头,其内侧为具乳头状凸起的柱头面。杨梅雌雄花的开放动态结果表明,3月下旬到4月初,杨梅花粉粒发育成熟,此时杨梅雌花的柱头大部分已经外露等待授粉,杨梅虽为雌雄异株植物,但其雌雄花发育进程非常一致,并不像一些植物存在雌雄异熟现象[24]。另外,散粉期杨梅花丝伸长,花药松弛外露,成熟的柱头呈“Y”状充分展开,且花柱表面具有大量类似糖类物质的粘性物存在,雌雄花的这种结构特征对于杨梅有效传粉十分有利。同时花粉活力测定结果表明,自然条件下杨梅花粉可保持3~4 d的活力,集中散粉期很短,约4~6 d。因此,我们认为在繁育系统中,杨梅雌雄花序在结构及发育进程上的协同发展,在一定程度上弥补了杨梅花粉寿命较短的不足,这应该是其提高繁殖能力的一种生存策略。
植物花粉母细胞减数分裂过程中,细胞质分裂有连续型和同时型2种主要类型。连续型分裂所形成的四分体排列在同一个平面上,成十字交叉形状,例如水稻Oryza sativa,小麦Triticum aestivum,玉米Zea mays及孝顺竹Bambusa multiplex等的四分体[25-26]。细胞质同时型分裂所形成的四分体呈四面体型,例如油菜Brassica campestris,烟草Nicotiana debneyi,花生Arachis hypogaea和蚕豆Vicia geminiflora等植物的四分体[27]。本研究结果表明:杨梅花粉母细胞减数分裂过程中,细胞质分裂以同时型分裂为主,2种类型的四分体同时存在,以四面体型四分体居多,这种表现形式与迪勃纳氏烟草类似[27]。杨梅雌花的胚胎学研究结果表明:杨梅为单心皮,1室,子房上位,直生胚珠,单珠被,厚珠心。在进化顺序上,一般认为双珠被比单珠被原始,厚珠心比薄珠心原始,倒生胚珠比直生胚珠原始[28-30]。根据杨梅胚珠结构的综合表现,我们推测杨梅该物种可能处于相对进化的位置。
[1] 戴斌,徐春明,王利芬,等.杨梅优选单株 ‘苏杨2号’的生物学特性及其ISSR鉴定[J].江西农业大学学报,2012,34(4): 676-681. DAI Bin,XU Chunming,WANG Lifen,et al.Biological characteristics and ISSR identification of‘SY-2’waxberry [J].Acta Agric Univ Jiangxi,2012,34(4):676-681.
[2] 何新华,陈力耕,陈怡,等.中国杨梅资源及利用研究评述[J].果树学报,2004,21(5):467-471. HE Xinhua,CHEN Ligeng,CHEN Yi,et al.Review on germplasm resources of Myrica and their exploitation in China [J].J Fruit Sci,2004,21(5):467-471.
[3] 王白坡,郑勇平,黎章矩,等.浙江省杨梅资源的利用及生态效益[J].浙江林学院学报,2001,18(2):155-160. WANG Baipo,ZHENG Yongping,LI Zhangju,et al.Utilization of Myrica rubra resources in Zhejiang and their ecological effect[J].J Zhejiang For Coll,2001,18(2):155-160.
[4] 李晓强,岑秀芬,韦鹏霄,等.杨梅组织培养防污染与防褐化研究[J].安徽农业科学,2012,38(19):9965-9967.LI Xiaoqiang,CENG Xiufen,WEI Pengxiao,et al.Study on prevention of pollution and browning in tissue culture of red bayberry(Myrica rubra Sieb et Zucc.)[J].J Anhui Agric Sci,2012,38(19):9965-9967.
[5] 许伟东.杨梅花芽生理分化期叶片内源激素含量变化[J].江西农业学报,2010,22(5):54-56. XU Weidong.Variation of endogenous hormone content in leaves of Myrica rubra at flower bud physiological differentiation stage[J].Acta Agric Jiangxi,2010,22(5):54-56.
[6] 谢小波,求盈盈,郑锡良,等.杨梅种间杂交及杂种F1的胚培养[J].果树学报,2009,26(4):507-510. XIE Xiaobo,QIU Yingying,ZHENG Xiliang,et al.Studies on the crossing between Myrica rubra and M.nana and embryo culture in vitro of its hybrid F1[J].J Fruit Sci,2009,26(4):507-510.
[7] 王涛,谢小波,戚行江,等.乌梅类杨梅大果型新品种黑晶的选育[J].中国南方果树,2008,37(1):29-30. WANG Tao,XIE Xiaobo,QI Xingjiang,et al.Breeding of large fruit type bayberry new variety[J].China Southern Fruit,2008,37(1):29-30.
[8] 钱皆兵,陈子敏,陈俊伟,等.优质大果杨梅新品系乌紫杨梅的生物学特性及其RAPD鉴定[J].果树学报,2007,24(1):64-67. QIAN Jiebing, CHEN Zimin,CHEN Junwei,et al.Biological characteristics and RAPD identification of a new high quality and large fruit bayberry line Wuzi[J].J Fruit Sci,2007,24(1):64-67.
[9] 潘鸿,何新华,李一伟,等.广西野生杨梅资源遗传多样性的ISSR分析[J].果树学报,2008,25(3):353-357. PAN Hong,HE Xinhua,LI Yiwei,et al.Genetic diversity of wild Myrica resources in Guangxi analysed by intersimple sequence repeats(ISSRs)[J].J Fruit Sci,2008,25(3):353-357.
[10] 孙志栋,柴春燕,陈惠云,等.果用杨梅慈溪若干地方品种亲源关系初析[J].分子植物育种,2008,6(6):1123-1126. SUN Zhidong,CHAI Chunyan,CHEN Huiyun,et al.Primary analysis of genetic relationship on some fruit waxberry landraces in Cixi City[J].Mol Plant Breeding,2008,6(6):1123-1126.
[11] 钱剑林,俞文生,王化坤,等.江浙地区杨梅主要品种的ISSR[J].植物资源与环境学报,2006,15(3):17-20. QIAN Jianlin,YU Wensheng,WANG Huakun,et al.Analysis on ISSR marker of primary cultivars of waxberry in Jiangsu and Zhejiang[J].J Plant Resour Environ,2006,15(3):17-20.
[12] 谢小波,求盈盈,戚行江,等.杨梅雌、雄种质遗传关系的RAPD和ISSR分析[J].果树学报,2008,25(2):198-202. XIE Xiaobo,QIU Yingying,QI Xingjiang,et al.Analysis of genetic relationship between male and female plants in Myrica rubra by RAPD and ISSR[J].J Fruit Sci,2008,25(2):198-202.
[13] 陈方永,王引,倪海枝,等.浙东南杨梅叶片气孔观察与相似性研究[J].植物遗传资源学报,2012,13(4):626-631. CHEN Fangyong,WANG Yin,NI Haizhi,et al.Stomata similarity of some Zhejiang Southeast bayberry leaves [J].J Plant Gen Resour,2012,13(4):626-631.
[14] 张喜焕,刘宁.4种类型矮杨梅花粉形态扫描电镜观察[J].贵州师范大学报,2005,23(2):6-9. ZHANG Xihuan, LIU Ning.SEM observation on the pollen morphology of four types of Myrica nana Cheval[J].J Guizhou Norm Univ Nat Sci,2005,23(2):6-9.
[15] 戚行江,郑锡良,梁森苗,等.不同分布地杨梅雄株花序特性及花粉扫描电镜观察[J].果树学报,2005,22 (2):175-178. QI Xingjiang,ZHENG Xiliang,LIANG Sengmiao,et al.Observation on the inflorescence characteristics and pollen of bayberry male plant grown in different regions of bayberry by scanning electron microscope(SEM)[J].J Fruit Sci,2005,22(2):175-178.
[16] 李三玉,戴善忠.杨梅花芽分化的初步研究[J].园艺学报,1980,7(1):9-16. LI Sanyu,DAI Shanzhong.Preliminary observations on flower bud differentition of Myrica rubra S.et Z.[J].Acta Hortic Sin,1980,7(1):9-16.
[17] SOGO A,TOBE H.Mode of pollen-tube growth in pistils of Myrica rubra(Myricaceae):a comparison with related families[J].Ann Bot,2006,97(1):71-77.
[18] 李正理.植物组织制片[M].北京:北京大学出版社,1996:15-50.
[19] SATO S,KATOH N,IWAI S,et al.Establishment of reliable methods of in vitro pollen germination and pollen preservation of Brassica rapa(syn.B.campestris)[J].Euphytica,1998,103:29-33.
[20] 夏国华.山核桃生殖生物学研究[D].南京:南京林业大学,2006:40-41. XIA Guohua.Reproductive Biology of Cayra eathyaensis Sarg.[D].Nanjing:Nanjing Forestry University,2006:40 -41.
[21] 方炎明.植物学[M].北京:中国林业出版社,2009:127-128.
[22] 林辰壹,韩文娟,祁晨霞,等.新疆高葶韭雄蕊开花动态特征与花粉育性研究[J].草业科学,2012,29(4):561-566. LIN Chenyi,HAN Wenjuan,QI Chenxia,et al.Studies on stamens blossom dynamic characteristics and pollen fertility of(Allium obliquum)[J].Pratac Sci,2012,29(4):561-566.
[23] 柴春燕,焦云,徐绍清,等.杨梅雄株花序特性与授粉研究[J].浙江林业科技,2011,31(5):1-6. CHAI Chunyan,JIAO Yun,XU Shaoqing,et al.Studies on characteristics of androphyte inflorescence and pollination of Myrica rubra[J].J Zhejiang For Sci Technol,2011,31(5):1-6.
[24] 戴攀峰,谭敦炎.雪莲的开花生物学特性及其生态适应意义[J].植物生态学报,2011,35(1):56-65. DAI Panfeng,TAN Dunyan.Floral biological characteristics of Saussurea involucrata in relation to ecological adaptation[J].Chin J Plant Ecol,2011,35(1):56-65.
[25] 金银根.植物学[M].北京:科学出版社,2010:209-210.
[26] 徐川梅,高欣,汤定钦.利用激光共聚焦显微镜研究孝顺竹花粉粒发育[J].林业科学,2010,46(11):158-162. XU Chuanmei,GAO Xin,TANG Dingqin.Observation of pollen development in Bambusa multiplex with laser scan confocal microscope[J].Sci Silv Sin,2010,46(11):158-162.
[27] 廖菊够,康洪梅,代瑾然,等.烟草大小孢子发生和雌雄配子体发育研究[J].西北植物学报,2013,33(1):11-16. LIAO Jugou,KANG Hongmei,DAI Jinran,et al.Megasporogenesis,microsporogenesis and development of female and male gametophyte of Nicotiana debneyi[J].Acta Bot Boreal-Occident Sin,2013,33(1):11-16.
[28] ENDRESS P K.Angiosperm ovules:diversity,development,evolution[J].Ann Bot,2011,107(9):1465-1489.
[29] ZHANG Di,WANG Ling,ZHUO Lihuan.Embryology of Iris mandshurica Maxim.(Iridaceae)and its systematic relationships[J].Plant Systematics and Evol,2011,293(1/4):43-52.
[30] 王自芬,任毅.被子植物胚珠研究进展[J].植物学通报,2007,24(1):49-59. WANG Zifen,REN Yi.Research progress of the angiosperm ovule[J].Chin Bull Bot,2007,24(1):49-59.
Biological characteristics of the Myrica rubra flower
SHEN Lifen,XIANG Weibo,FAN Caiting,JIN Peng,ZHOU Mingbing,XU Chuanmei
(The Nurturing Station for the State Key Laboratory of Subtropical Silviculture,Zhejiang A&F University,Lin’an 311300,Zhejiang,China)
The purpose of this study was to determine the biological characteristics of the Myrica rubra flower. The external structure of the anemophilous M.rubra flower was observed with a dissecting microscope;then pollen viability and development of the internal structure were determined by the fluorochrome reaction and eosin staining.Results showed that the male flower had 8 to 20 anthers and every anther had four pollen sacs. Most microspore tetrads had a tetrahedral type configuration with only a few having a successive configuration. Most mature pollen grains had two nuclei and three germinal apertures;pollen vitality was more than 90%.The female inflorescence consisted mainly of the stigma,style,and ovary.The superior ovary had a single carpel and one room.Also,the orthotropous ovule had a single integument and a thick nucellus.[ch,4 fig.30 ref.]Key words:cash forestry;Myrica rubra;anther;ovary;biological characteristics
S667.6;Q944.4
A
2095-0756(2015)02-0278-07
浙 江 农 林 大 学 学 报,2015,32(2):278-284
Journal of Zhejiang A&F University
10.11833/j.issn.2095-0756.2015.02.016
2014-03-07;
2014-04-21
浙江省自然科学基金资助项目(Y13C160036)
沈利芬,从事森林可持续性经营理论与技术研究。E-mail:shenlf0715@163.com。通信作者:徐川梅,从事植物细胞生物学研究。E-mail:xuchuanmei1979@126.com
猜你喜欢
——植物学教材质疑(六)
生命科学研究(2022年1期)2022-03-14 07:50:40
铜仁学院学报(2018年7期)2018-09-08 08:50:40
现代园艺(2017年22期)2018-01-19 05:06:57
南风(2017年34期)2018-01-16 07:29:09
中国稻米(2017年6期)2017-12-28 02:13:01
科学种养(2017年6期)2017-06-13 08:42:04
科学种养(2017年5期)2017-05-17 10:18:50
广西植物(2016年10期)2016-11-11 06:51:39
西南农业学报(2016年4期)2016-05-17 05:41:45
作物研究(2015年4期)2015-03-24 00:37:07
