
浙江天童冬青属植物的种群结构及更新类型
2015-03-23 09:21杨同辉章建红夏晨诚蒋笑丽达良俊
浙江农林大学学报 2015年1期
曹 菁,杨同辉,章建红,夏晨诚,蒋笑丽,达良俊
浙江天童冬青属植物的种群结构及更新类型
曹 菁1,杨同辉2,章建红2,夏晨诚1,蒋笑丽2,达良俊1
(1.华东师范大学 生态与环境科学学院,上海 200241;2.浙江省宁波市农业科学研究院,浙江 宁波315040)
为加强冬青属Ilex植物资源的保护和利用,采用标准样方法调查分析了浙江宁波天童4种冬青种群结构及更新类型。结果表明:①所调查群落共出现木本植物61种,除草本层外,林木层由下至上可分为灌木层和乔木层,其中在乔木层之上存在分别由小果冬青Ilex micrococca,马尾松Pinus massoniana,天童锐角槭Acer acutum构成的不连续的超高层。②所调查冬青属植物的年龄结构可分为4种类型:增长型(皱柄冬青Ilex kengii),稳定型(铁冬青Ilex rotunda),成熟型(冬青Ilex chinensis)和衰退型(小果冬青Ilex micrococca)。③冬青属植物总体生长良好,但有些种如小果冬青属于衰退型,无幼苗,无萌枝,种子更新困难,冬青幼苗难以通过环境筛导致幼树严重匮乏,需要采取适当人工育种以及栽培技术来加以繁殖保护,并对优良种质加以选育和利用。 图2表2参30
森林生态学;冬青属;胸径级频率;种群结构;更新类型;保护措施
冬青属Ilex植物大都为常绿阔叶树种,是典型的亚热带地带性植被,喜生于山坡、谷地、溪边杂木林或灌丛中,喜光照,适应性强,耐阴耐寒耐低温,喜温暖湿润的气候,适应性较强,全世界已发现的冬青属植物有500~600种,中国有204种,浙江省分布37种2变种[1],是浙江省常绿阔叶林的主要树种之一[2]。冬青属植物在中医药材、园林绿化方面发挥着重要作用,国内对其研究大都集中于遗传特性[3-4]、化学成分[5]、生物学特性[6-7]、繁殖技术[8-9]等方面,但对其自然种群结构的研究不多。天童地处中亚热带北缘地区,该地区保存了相对较好的地带性常绿阔叶林,植物资源丰富,是不可多得的 “浙江植物基因宝库”[10]。区域植物资源的发掘对种质资源保护和利用具有重要的研究和应用价值。本研究主要通过天童4个冬青属植物的群落学调查,分析其年龄结构及更新类型,探讨其种群结构的形成原因及更新方式,为冬青属植物的保护和利用提供参考。
1 研究区域和方法
1.1 研究地区概况
天童国家森林公园位于浙江省宁波市东南部,距宁波市28.0 km,地处29°48′N,121°47′E,面积为349.0 hm2。该地森林植被保存良好,是浙江省东部地带性植被类型的代表性地段。气候为温暖潮湿的亚热带季风气候,年平均气温为16.2℃,最热月为7月,平均温度为28.1℃;最冷月为1月,平均气温为4.2℃;大于10.0℃的年积温为5 166.2℃,吉良(Kira)的温暖指数(WI)是135.0℃·月,寒冷指数(CI)是-0.8℃·月。年降水量为1 374.7 mm,多集中在夏季。年平均相对湿度为82%,变率不大。年蒸发量为1 320.1 mm,小于降水量。森林公园内土壤主要为山地黄红壤,成土母质主要是中生代的沉积岩及部分酸性火成岩和花岗岩残积风化物[11]。
1.2 调查研究方法
1.2.1 样方设置与调查内容 于天童国家森林公园玲珑岩330.0 m处沿主游步道向下,按种群出现顺序分别设置了冬青Ilex chinensis,铁冬青Ilex rotunda,皱柄冬青Ilex kengii,小果冬青Ilex micrococca等4 个20.0 m×20.0 m样方,样方的坡度分别为38.0°,40.0°,35.0°,20.0°,铁冬青样方位于沟谷左侧,冬青样方位于山体沟谷上方,皱柄冬青样方位于沟谷右侧,小果冬青样方位于沟谷下方堆积地。对样方内所有高度超过 1.5 m的木本植物进行每木调查。鉴别每株植物种类,测定记录其胸径(DBH,cm),高度(H,m),对植株中高度超过1.5 m的萌枝也同时测定记录胸径(DBH,cm),草本层多盖度的综合级的计量采用Braun-Blanquet推测的目测估计;分别记录每株乔木幼树的个体高度(H,m)。
1.2.2 种群年龄级划分标准 由于个体年龄常难于确定,所以在实际工作中一般采用空间代替时间的方法,即以胸径级代替年龄级进行分析[12]。根据冬青属植物的生长特点,其大小级采用以下方法划分[11,13-16]:胸径<2.5 cm的幼树幼苗按胸径划分为2级:高度<0.33 m,为第1级,高度≥0.33 m,胸径<2.5 cm为第2级。胸径≥2.5 cm的个体,按胸径每增加5.0 cm为1级,胸径≥52.5 cm的为1级,共13级。在此基础上将冬青属植物种群按5级的划分标准进行处理:Ⅰ级幼苗(高度<0.33 m)包括第1级,Ⅱ级幼树(高度≥0.33 m,胸径<2.5 cm)包括第2级,Ⅲ级小树(2.5 cm≤胸径<7.5 cm)包括第3级,Ⅳ级中树(7.5 cm≤胸径<22.5 cm)包括第4级、第5级和第6级,Ⅴ级大树(胸径≥22.5 cm)包括第7级、第8级、第9级、第10级、第11级、第12级和第13级。
1.2.3 数据处理方法 群落的优势种数量由优势度分析法确定:为优势种的实际百分比与理想百分比的偏差(deviation),xi为前位树种(top species,T)的相对胸高断面积(RBA,relative basal area,%),x为以优势种(dominant species)数量确定的优势种理想百分比(ideal percentageshare),xj为剩余种(remaining species)的百分比。N为总种数。如果群落只有1个优势种,则优势种的理想百分比为100.0%。如果有2个优势种,则它们的理想百分比为50.0%,如果有3个优势种,则理想百分比为33.3%,依此类推。优势种的数量由d最小的来决定,假如x为100.0%时d最小,则优势种数量为1,假如x为50.0%时d最小,则优势种数量为2,依此类推[17]。
群落的优势种由物种重要值来确定[18]:Iv=(Dr+Pr+Fr)/3;Dr=D(某个种的株数)/∑D(全部种的总株数);Pr=P(某个种的胸高断面积)/∑P(全部种的胸高断面积);Fr=F(某个种的频度)/∑F(全部种的总频度)。Iv(importance value)为物种重要值,Dr(relative density)为物种相对密度,Pr(relative prominence)为物种相对优势度,Fr(relative frequency)为物种相对频度。木本层用相对胸高断面积(RBA,%)表示其相对优势度,草本层相对优势度用相对体积当量(RVEV,relative volume equivalent-value,%)表示[2,19],公式如下:其中:Hi为第 i种的最大高度(m),Ci为第 i种的盖度(%)。草本层调查时采用Braun-Blanquet推测的目测估计法,调查了最大高度与盖度,未采集株数信息,草本层中物种在群落中的地位与作用仅以盖度和相对优势度来表示。
2 结果与分析
2.1 群落垂直结构
植株的高度级频率分布反映了树种在森林中的各种特征以及群落的分层结构[19]。所调查的群落(总盖度依次为90.0%,95.0%,98.0%和88.0%)除草本层外,林木层可以分为2层,由下至上分别为灌木层和乔木层。灌木层个体高度级的顶点在6.0~8.0 m,盖度分别达到65.0%,45.0%,55.0%和35.0%,主要由山茶科Theaceae的窄基红褐柃Eurya rubiginosa,连蕊茶Camellia fraterna,杨桐Adinandra millettii等,樟科Lauraceae的浙江新木姜子Neolitsea aurata,黄丹木姜子Litsea elongata,红楠Machilus thunbergii等,壳斗科Fagaceae的栲树Castanopsis fargesii,苦槠Castanopsis sclerophylla,白栎Quercus fabri等,山矾科 Symplocaceae的薄叶山矾 Symplocos anomala,山矾 Symplocos sumuntia,四川山矾 Symplocos setchuensis等,金缕梅科Hamamelidaceae的杨梅叶蚊母树Distylium myricoides,蔷薇科Rosaceae的光叶石楠Photinia glabra,木犀科Oleaceae的华东木犀Osmanthus cooperi等及群落优势种如铁冬青,木荷Schima superba,交让木Daphniphyllum macropodum等的幼树构成;乔木层个体高度级的顶点为16.0~ 18.0 m,盖度依次为80.0%,85.0%,90.0%和75.0%,主要由群落优势树种铁冬青,马尾松Pinus massoniana,青冈Cyclobalanopsis glauca,木荷,樟树Cinnamomum camphora,交让木构成。其中有3个样方在乔木层之上存在分别由小果冬青、马尾松和天童锐角槭Acer acutum构成的不连续的超高层(盖度分别为25.0%,5.0%和35.0%)[20](图1)。
2.2 群落种类组成
所调查的群落中共出现木本植物61种,从生活型区分来看,常绿针叶树2种,常绿阔叶树41种,落叶阔叶树18种。针叶树为阳性的马尾松和杉木Cunninghamia lanceolata;常绿阔叶树种以壳斗科(米槠Castanopsis carlesii,栲树,青冈等),山茶科(连蕊茶,木荷,杨桐等),樟科(红楠,黄丹木姜子,浙江新木姜子等),冬青科Aquifoliaceae(冬青,铁冬青,皱柄冬青等),山矾科(山矾,四川山矾,薄叶山矾等)的树种为主,占总数的67.2%;落叶阔叶树种占总种数的29.5%,主要有冬青科小果冬青、樟科的檫木Sassafras tzumu,钓樟L.umbellata,漆树科Anacardiaceae南酸枣Choerospondias axillaris,金缕梅科枫香Liquidambar formosana等。
在调查的4个样方中分别出现28,23,27和25个树种,铁冬青为群落的优势种,在群落中重要值达到9.8%,小果冬青虽为群落伴生种,但其重要值9.4%与铁冬青相当,其最大高度和最大胸径更是分别达到20.6 m和38.9 cm,分别与木荷、苦槠、南酸枣和青冈等共同构成其群落主要组成种。冬青和皱柄冬青分别为其群落的伴生种,重要值仅为6.2%和9.9%,其个体最大高度仅有8.5 m和10.5 m,个体最大胸径则达到16.8 cm和25.7 cm;冬青的萌枝率在4种冬青属植物中最高(38.9%),其次为铁冬青(25.0%)和皱柄冬青(17.9%),而小果冬青无萌枝。4个样方中仅皱柄冬青和冬青在群落草本层中有幼苗出现,优势度分别为3.6%和0.2%(表1,表2)。
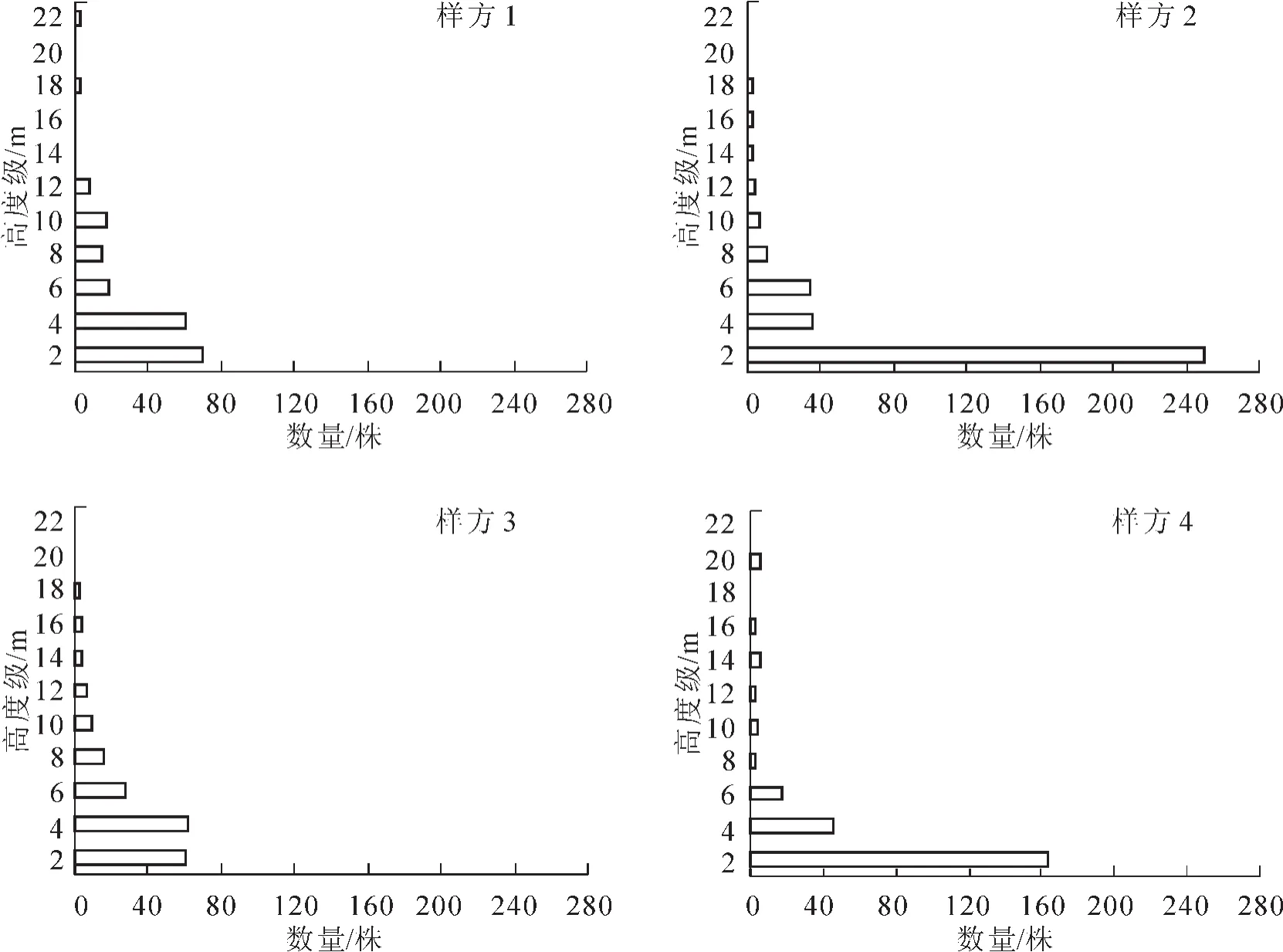
图1 群落高度级频率分布Figure 1 Height classes frequency distribution in the communities
2.3 冬青属植物的年龄结构
种群的年龄结构反映了种群过去的更新机会和未来的演化趋势[21],研究种群的年龄结构对深入分析种群的动态和进行预测有重要意义[22-25]。所调查冬青属植物的年龄结构可分为以下几种类型(图2):①增长型(皱柄冬青)。幼龄级个体较多(64.1%),明显多于中龄(5.1%)及大龄(5.1%)植株,幼苗库资源丰富(25.7%),反映其天然更新较好。②稳定型(铁冬青)。各大小级都有个体分布并具连续性,拥有较多的幼苗、幼树和少量的大树,种群处于稳定发展阶段。③成熟型(冬青)。种群个体在各年龄级分布不规则,但存在一定延续性,幼苗(55.5%)多于小树(11.1%)和中树(27.8%),幼树(5.6%)较少,这种结构介于稳定型与衰退型之间,但此时群落外貌、结构已分化成熟,故称为成熟型种群[26]。④衰退型(小果冬青)。种群大小结构在不同年龄级有缺失现象,幼树(50.0%)与大树(50.0%)比例相当,幼苗资源库匮乏,更新不良。
3 结论与讨论
3.1 冬青属植物种群结构的形成原因
冬青是群落伴生种,对环境的要求并不严格,适应性强[27]。在所调查样方中萌枝率较高,幼苗较多但幼树却很少,说明在过渡阶段必然存在一道环境筛,许多个体难以通过环境筛而大量死亡。又由于群落总盖度达到90.0%,乔木层盖度也达到80.0%,因而林内光量相对较少,再加上台风等自然干扰,马尾松、枫香等阳性树种的竞争,导致现阶段冬青呈现幼苗明显多于幼树,而缺乏大树的年龄结构;皱柄冬青也为群落的伴生种,幼苗耐阴大树喜光[22],由于优势种青冈、木荷等大树的存在导致该群落的郁闭度高(98.0%),林下光照较少。皱柄冬青在群落中幼树及小树所占比例较大且连续性更新,而中树和大树所占比例较少,其更新具有机会性和波动性[2],一旦有林窗形成,皱柄冬青将会快速生长。
铁冬青为耐阴树种,是其地形顶级的优势种,尤其是幼苗耐阴性极强,喜生于温暖湿润气候[1],所调查样方中铁冬青幼龄级个体最多(约52.6%),萌枝率也较高(25.0%)。由于栲树、木荷等大树的枯死形成的林窗,中树和大树将逐渐发展,随着年龄增长,铁冬青对光的需求量增大。由于种内竞争而产生自疏现象,故中、大个体数量较少。小果冬青系落叶大乔木,为阳性树种,是群落伴生种,小果冬青样方位于沟谷下方的堆积地,群落郁闭度高达88.0%。正因此样方中小果冬青无幼苗,无萌枝,幼树较少,而仅有的几株大树亦存在于超高层中,这与王良衍等[8]调查的天童3个立地类型群丛中小果冬青数量较少,在林内呈单株分布,多处在乔木层的超高层的研究一致。
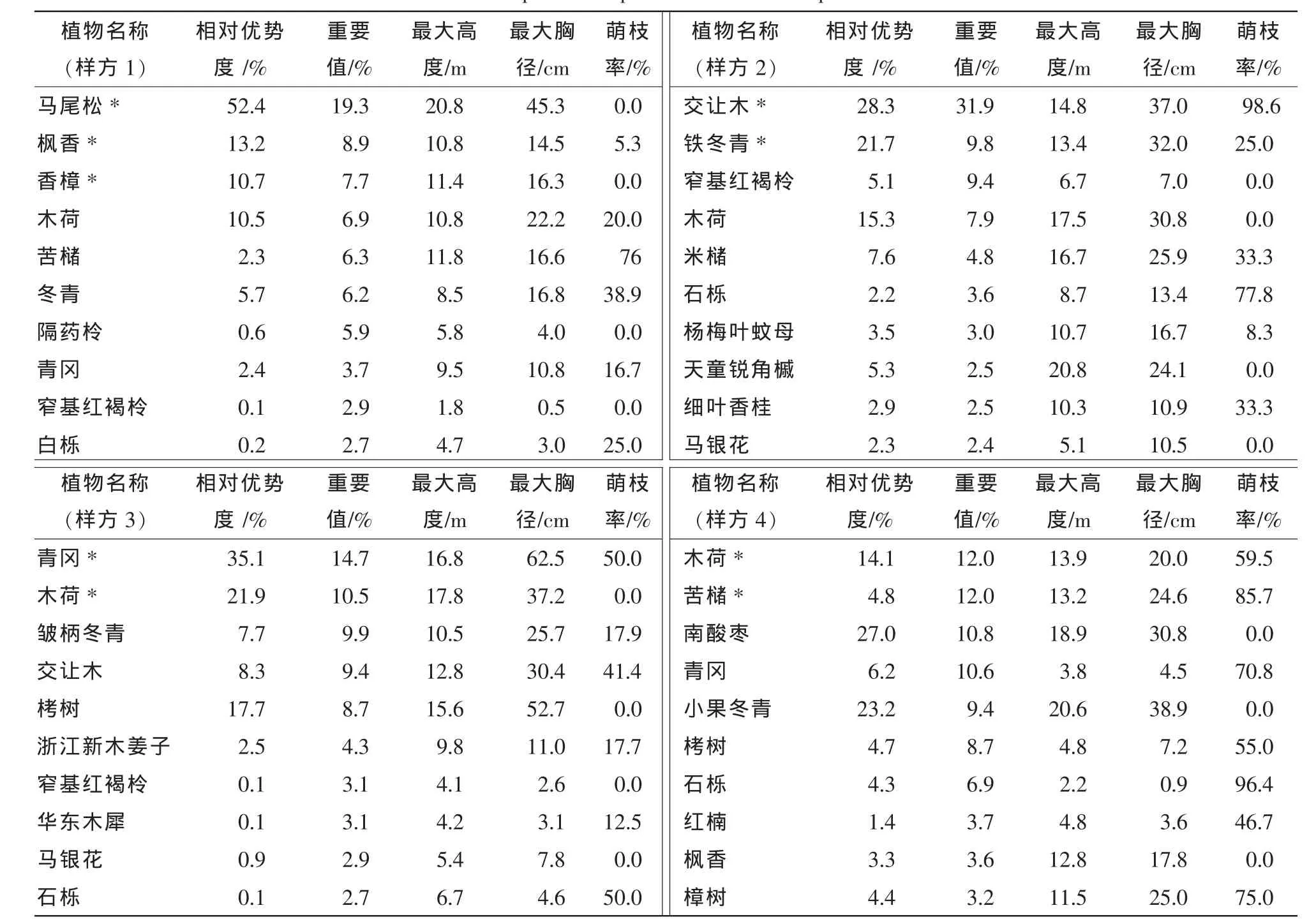
表1 样方主要组成树种及相关参数Table 1 Main species composition and related parameters
3.2 冬青属植物的更新类型
冬青缺乏大树,其年龄结构为稳定型,小树较少,但幼苗资源丰富,更新能力强,铁冬青幼树、小树、中树和大树连续分布,随着年龄的增长,数量逐渐减少,种群处于稳定发展阶段。样方1和样方2中都有枯立木形成的林窗,林窗更新是森林中一种非常重要的更新机制[28-29],冬青的更新以萌枝与幼苗2种方式,铁冬青则以萌枝更新为主。与此同时由于林窗形成导致林内光量增加,冬青和铁冬青大树的比例也将增加。由冬青种群结构的特征可以看出,许多幼苗个体难以通过环境筛而使得幼树比例明显减少,这种现象很可能导致种群丰富度下降[30]。皱柄冬青的更新也有萌枝与实生苗2种方式,但萌枝率较低,主要依靠种子更新,其幼龄级个体数量明显多与中树和大树,属于典型的金字塔增长型。小果冬青属于衰退型,无幼苗,严重缺乏小树和中树,幼树和大树所占比例相当,更新具有不连续性,相关研究[8]显示小果冬青种子细小坚硬、发芽率低,导致种群更新困难,必须采取适当人工措施促进其更新。
3.3 冬青属植物的保护和利用
冬青属植物树姿优美、果实艳丽,具有较高的观赏价值,近年来作为乡土树种逐渐应用到绿化建设中。中国对冬青属植物的应用研究尚不完善,其品种较多、分布范围较广,资源量少,特别需要加强自然种质资源的保护,尤其是冬青属植物自然种群分布和更新发育情况。根据本研究可以看出:冬青属植物总体生长良好,铁冬青处于稳定发展阶段,皱柄冬青呈增长趋势,但个别种在自然更新方面有困难,如小果冬青,种群正朝衰退方向发展,其种子发芽率低[8],而且无萌枝。再如冬青,年龄结构介于稳定型与衰退型之间,幼苗难以通过环境筛而导致幼树资源严重匮乏。因此,需要采取适当人工育种以及栽培技术加以繁殖保护,可通过无性繁殖方式对优良的种质加以选育,以期更多的优良品种得到实际应用。冬青属植物种群结构及其更新类型的研究有助于进一步了解冬青属植物的自然生长情况以及为园林应用提供理论依据。
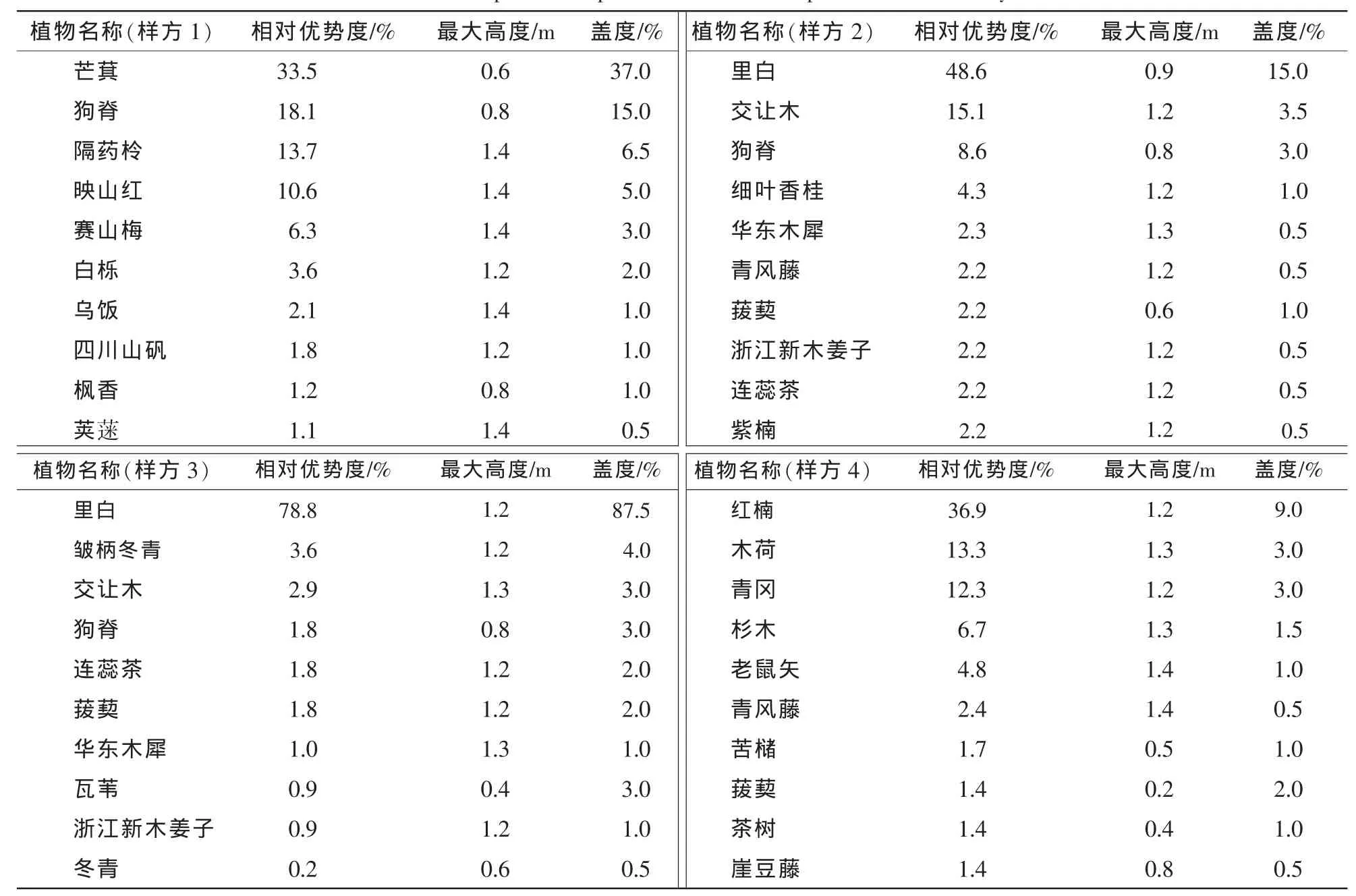
表2 草本层主要组成种及相关参数Table 2 Main species composition and related parameter in herb layer
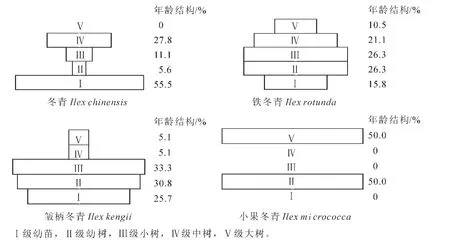
图2 4种冬青属植物的年龄结构图Figure 2 The age structure of four Ilex species
4 致谢
论文写作中得到杨晓东博士和Arshad Ali博士的帮助,在此表示诚挚的感谢。
[1] 陈书坤.中国植物志:第45卷第2分册[M].北京:科学出版社,1999.
[2] 达良俊,杨永川,宋永昌.浙江天童国家森林公园常绿阔叶林主要组成种的种群结构及更新类型[J].植物生态学报,2004,28(3):376-384. DA Liangjun,YANG Yongchuan,SONG Yongchang.Population structure and regeneration types of dominant species in an evergreen broadleaved forest in Tiantong National Forest Park,Zhejiang Province,Estern China[J].Acta Phytoecol Sin,2004,28(3):376-384.
[3] 张凤琴,刘国民,周鹏,等.用RAPD技术探讨冬青属苦丁茶的遗传差异、亲缘关系与分类地位[J].云南植物研究,2004,26(6):637-644. ZHANG Fengqian,LIU Guomin,ZHOU Peng,et al.A atudy on genetic variations,relationships and classification of several“KUDINGCHA” species in Ilex(Aquifoliaceae)based on RAPD analysis[J].Acta Bot Yunnan,2004,26 (6):637-644.
[4] 章建红,高云振,张斌,等.26种冬青属植物遗传多样性分析[J].西北植物学报,2011,31(3):504-510. ZHANG Jianhong,GAO Yunzhen,ZHANG Bin,et al.Genetic diversity of llex L.tree species[J].Acta Bot Boreal-Occident Sin,2011,31(3):504-510.
[5] 解军波,李萍.冬青属植物化学成分及药理活性研究进展[J].中草药,2002,33(1):85-88. XIE Junbo,LI Ping.Advances in research of chemistry and pharmacology of Ilex L.[J].Chin Trad Herb Drugs,2002, 33(1):85-88.
[6] 张璐,张纪林,教忠意,等.不同光照条件下3种冬青属植物的光合特征日变化研究[J].西北植物学报,2006,26(3):490-495. ZHANG Lu,ZHANG Jilin,JIAO Zhongyi,et al.Diurnal photosynthetic changes of three Ilex species under different light circumstances[J].Acta Bot Boreal-Occident Sin,2006,26(3):490-495.
[7] 李相传,孙柏年,林志成,等.冬青属植物的叶表皮特征及其分类学意义[J].兰州大学学报:自然科学版,2010,46(4):13-29. LI Xiangchuan,SUN Bainian,LIN Zhicheng,et al.Epidermal anatomy of the genus Ilex and its taxonomic significance [J].J Lanzhou Univ Nat Sci,2010,46(4):13-29.
[8] 王良衍.小果冬青栽培技术[J].林业实用技术,2004(12):15-16. WANG Liangyan.Cultivation techniques of Ilex micrococoa Maxim[J].Pract For Techn,2004(12):15-16.
[9] 杨永川,王良衍,宋坤,等.2种优良乡土冬青的繁育及栽培技术[J].浙江林学院学报,2005,22(4):406-409. YANG Yongchuan,WANG Liangyan,SONG Kun,et al.Reproduction and forestation techniques of two native tree species:Ilex micrococca and I.buergeri[J].J Zhejiang For Coll,2005,22(4):406-409.
[10] 喻晓雁,谈献和,邹立思.浙江天童森林公园百合科药用植物资源调查[J].中医学报,2011,26(6):710-712. YU Xiaoyan,TAN Xianhe,ZOU Lisi.Investigation on resources of medicinal liliaceae in Zhejiang Tiantong Forest Park[J].China J Chin Med,2011,26(6):710-712.
[11] 宋永昌,王祥荣.浙江天童国家森林公园的植被和区系[M].上海:上海科学技术文献出版社,1995:1-130.
[12] 金则新.浙江天台山七子花种群结构与分布格局研究[J].生态学杂志,1997,16(4):15-19. JIN Zexin.A study of population structure and distribution pattern of Heptacodium miconioides in the Tiantai Mountain,Zhejiang[J].Chin J Ecol,1997,16(4):15-19.
[13] 陈小荣,李乐,夏家天,等.百山祖亮叶水青冈种群结构和分布格局[J].浙江农林大学学报,2012,29(5):647-654. CHEN Xiaorong,LI Le,XIA Jiatian,et al.Population structure and distribution of Fagus lucida in the Baishanzu forest[J].J Zhejiang A&F Univ,2012,29(5):647-654.
[14] 韩路,王海珍,彭杰,等.不同生境胡杨种群径级结构与格局动态研究[J].北京林业大学学报,2010,32 (1):7-12. HAN Lu,WANG Haizhen,PENG Jie,et al.Size-class structure and distribution pattern of Populus euphratica Oliv. in different habitats[J].J Beijing For Univ,2010,32(1):7-12.
[15] 胡小兵,于明坚.青冈常绿阔叶林中青冈种群结构与分布格局[J].浙江大学学报:理学版,2003,30(5):574-579. HU Xiaobing,YU Mingjian.Size structure and distribution pattern of Cyclobalanopsis glauca population in evergreen broad-leaved forests[J].J Zhejiang Univ Sci Ed,2003,30(5):574-579.
[16] 康华靖,陈子林,刘鹏,等.大盘山自然保护区香果树种群结构与分布格局[J].生态学报,2007,27(1):389-396. KANG Huajing,CHEN Zilin,LIU Peng,et al.The population structure and distribution pattern of Emmenopterys henryi in Dapanshan Natural Reserve of Zhejiang Province[J].Acta Ecol Sin,2007,27(1):389-396.
[17] OHSAWA M.Difference of vegetation zones and species strategies in the subalpine region of Mt.Fuji[J].Vegetatio, 1984,57(1):15-52.
[18] 宋永昌.植被生态学[M].上海:华东师范大学出版社,2001:45-46.
[19] 杨同辉,王良衍.浙江天童舟山新木姜子群落结构及其更新类型[J].浙江林学院学报,2010,27(6):877-883. YANG Tonghui,WANG Liangyan.Community structure and regeneration types of Neolitsea sericea in Tiantong, Zhejiang Province[J].J Zhejiang For Coll,2010,27(6):877-883.
[20] OHSAWA M.Anaga Cloud Forest:A Comparative Study on Evergreen Broad-leaved Forests and Trees of the Canary Islands and Japan[R].Chiba:Chiba University,1999:67-87.
[21] 张谧,熊高明,赵常明,等.神农架地区米心水青冈-曼青冈群落的结构与格局研究[J].植物生态学报,2003,27(5):603-609. ZHANG Mi,XIONG Gaoming,ZHAO Changming,et al.The structures and patterns of a Fagus engleriana-Cyclobalanopsis oxyodon community in Shennongjia area,Hubei Province[J].Acta Phytoecol Sin,2003,27(5):603-609.
[22] 宋萍,洪伟,吴承祯,等.珍稀濒危植物桫椤种群结构与动态研究[J].应用生态学报,2005,16(3):413-418. SONG Ping,HONG Wei,WU Chengzhen,et al.Population structure and its dynamics of rare and endangered plant Alsophila spinulosa[J].Chin J Appl Ecol,2005,16(3):413-418.
[23] 赖江山,张谧,谢宗强.三峡库区常绿阔叶林优势种群的结构和格局动态[J].生态学报,2006,26(4):1073 -1079. LAI Jiangshan,ZHANG Mi,XIE Zongqiang.Structure and distribution pattern of dominant populations in the evergreen broad-leaved forest in three gorges reservoir area[J].Acta Ecol Sin,2006,26(4):1073-1079.
[24] TANOUCHI H,YAMAMOTO S.Structure and regeneration of canopy species in an old-growth evergreen broadleaved forest in Aya district,southwestern Japan[J].Vegetatio,1995,117(1):51-60.
[25] MASAKI T,TANAKA H,TANOUCHI H,et al.Structure,dynamics and disturbance regime of temperate broadleaved forests in Japan[J].J Veg Sci,1999,10(6):805-814.
[26] 蔡飞,宋永昌.武夷山木荷种群结构和动态的研究[J].植物生态学报,1997,21(2):138-148. CAI Fei,SONG Yongchang.A study on the structure and dynamics of Schima superba population on Wuyi Mouatain [J].Acta Phytoecol Sin,1997,21(2):138-148.
[27] 蔡卓,马长乐.冬青属植物园林应用特征研究[J].现代园艺,2013(4):163-164. CAI Zhuo,MA Changle.Research on landscape application of Ilex L.[J].Xiandai Hort,2013(4):163-164.
[28] RUNKLE J R.Gap regeneration in some old-growth forests of the eastern United States[J].Ecology,1981,62(4): 1041-1051.
[29] YAMAMOTO S.Gap characteristics and gap regeneration in subalpine old-growth coniferous forests,central Japan [J].Ecol Res,1995,10(1):31-39.
[30] CHO D S,BOERNER R E J.Canopy disturbance patterns and regeneration of Quercus species in two Ohio oldgrowth forest[J].Vegetation,1991,93(1):9-18.
Population structure and regeneration types of Ilex spp.in Tiantong National Forest Park,Zhejiang Province
CAO Jing1,YANG Tonghui2,ZHANG Jianhong2,XIA Chencheng1,JIANG Xiaoli2,DA Liangjun1
(1.Department of Ecological and Environmental Sciences,East China Normal University,Shanghai 200241,China;2. Ningbo Academy of Agriculture Science,Ningbo 315040,Zhejiang,China)
The Ilex species has been recognized for potential role in landscaping.Most of the research focused on genetic,biological,chemical and propagation characteristics of the species.This study aimed to provide the ways to strengthen conservation measures and utilization for Ilex species based on age and population structure. The four sampling plots(each of 20 m×20 m size)were established in subtropical,evergreen broadleaf forests in Tiantong National Forest Park,Zhejiang Province.All woody species in the plots were identified and measured to quantify population structure and regeneration types.The complete randomized design (CRD)with sub-sampling of Ilex species in each sampling plot were used in this study.The four plots were used as replicates to represent different Ilex species in the study area.Results showed:1)61 woody species overall with a forest structure mainly consisting of a tree canopy layer and a shrub layer along with an herb layer.An emergent layer,consisting of Ilex micrococca,Pinus massoniana,and Acer acutum species within each plot,was found only in three plots.2)The age structure of Ilex species in all studied plots were categorized into four types:(a)the increasing pyramid type--Ilex kengii,(b)the stable type--Ilex rotunda,(c)the mature type--Ilex chinensis,and (d)the decreasing type--Ilex micrococca.3)Overall Ilex species grew very well,but Ilex micrococca,lacking saplings and sprouts,needed special attention along with Ilex chinensis which had very lit-tle regeneration.Thus,suitable artificial breeding and cultivation techniques should be applied for conservation of Ilex species and favorable breeding germplasm with necessary protection measures being utilized.[Ch,2 fig. 2 tab.30 ref.]
forest ecology;Ilex species;DBH class distribution;population structure;regeneration types;protective measures
S718.54
A
2095-0756(2015)01-0076-08
2014-04-22;
2014-06-18
浙江省宁波市重大(重点)科技攻关项目(2013C11034);国家自然科学基金资助项目(40971041);上海市科学技术委员会科研计划项目(08DZ1203102)
曹菁,从事植被生态学和城市生态学研究。E-mail:ErinCECNU@163.com。通信作者:达良俊,教授,博士生导师, 从事植被生态学和城市生态学等研究。E-mail:ljda@des.ecnu.edu.cn
猜你喜欢
北华大学学报(自然科学版)(2022年5期)2022-11-21
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
新农民(2020年2期)2020-12-08
中华建设(2020年5期)2020-07-24
河北果树(2020年1期)2020-02-09
小学生必读(低年级版)(2018年6期)2018-09-08
农村百事通(2016年24期)2017-01-07
中央民族大学学报(自然科学版)(2015年3期)2015-06-11
