
旱地小麦理想株型与生长冗余
2015-03-13 03:28王建永李朴芳程正国AsfaBatool吕广超熊友才
生态学报 2015年8期
王建永,李朴芳,程正国,Asfa Batool,吕广超,祝 英,2,熊友才,*
1 兰州大学草地农业生态系统国家重点实验室, 干旱农业生态研究所, 生命科学学院, 兰州 730000 2 甘肃省科学院生物研究所, 兰州 730000
旱地小麦理想株型与生长冗余
王建永1,李朴芳1,程正国1,Asfa Batool1,吕广超1,祝 英1,2,熊友才1,*
1 兰州大学草地农业生态系统国家重点实验室, 干旱农业生态研究所, 生命科学学院, 兰州 730000 2 甘肃省科学院生物研究所, 兰州 730000
自1968年Donald提出作物理想株型(ideotype)以来,众多学者在如何减少生长冗余、塑造理想株型方面做了大量努力,在旱地小麦育种策略和栽培管理模式创新方面取得了一定进展。而该方面的进展有限,不同领域的研究者对理想株型和生长冗余的认识存在严重分歧。综述了近年来旱地小麦理想株型研究进展,以过去20年黄土高原雨养农业区的观测数据为主进行了集成分析,将产量分别与根系生物量、地上生物量和株高等指标进行回归分析,勾画了旱地小麦在根系、茎秆和分蘖等器官中生长冗余的演变趋势,对生长冗余产生的生态学机制展开了分析,并对理想株型与生长冗余的互作关系进行了讨论。已有的研究进展表明,旱地小麦理想株型演变是一个不断消减生长冗余,但又无法完全消除冗余的复杂过程,一定程度的冗余存在是理想株型发生的物质基础。旱地小麦理想株型育种必须以基因型和环境互作关系为基础,通过减少个体间的竞争强度和个体大小不整齐性,促进物质和能量更多地向籽粒迁移,最终提高种群产量。综上所述,旱地小麦理想株型选择需建立在生长冗余理论基础上,根据生态学基本原理对基因型和表现型进行耦合分析与选择权衡。
旱地小麦; 理想株型; 生长冗余; 互作关系; 研究进展
旱地小麦理想株型选择一直是生态学、作物学和生物学的热点问题。过去五十多年来,相关领域的学者开展了一系列提高旱地小麦产量及抗旱性的理论与实践研究[1-5],但诸多努力多集中在提高小麦的耐旱性上,尤以作物个体耐旱能力、抗旱基因定位和表达等方面为侧重点[6]。传统观点认为,抗旱性越强的品种其根系越庞大,因此大根系(指具有较多的根系生物量,较大的根系分布范围,意味着植株有相对较高的根冠比)单株产量高的个体通常为抗旱育种选择的目标[7]。而个体耐旱能力的提高是以提高个体竞争力为基础的,这必将导致作物群体产量的下降[8-10]。在以高产高效为主要目标的现代农业体系中,以大根系为选择标准的思想并不能给雨养农业区小麦产量的提高带来任何实质性的改变[11-12]。抗旱育种成效不显著,其关键原因是不同领域的研究者对干旱胁迫下的“抗旱”性状理解有很大差异。微观生物学如生物化学和分子生物学领域的学者认为抗旱性即个体细胞或细胞团组织对水分胁迫的存活能力或耐受性;而作物栽培和作物育种学领域的学者认为抗旱性是水分亏缺环境下作物或品种产量稳定性的表现;从种群生态学和进化生态学的角度讲,抗旱性为:与水分充分供给时作物群体最高产量相比在水分亏缺时减产最小的生态适应机制。正是由于对抗旱性的认识不同,造成了干旱半干旱区育种策略上的误区[10]。如何在干旱环境下通过权衡个体竞争力和群体产量之间的比重,最终提高作物单位产量成为农学界和生物学界一个公开的论战[13-14]。
1968年Donald理想株型(ideotype)概念的提出,在农学界、生态学和生物学界引起了强烈的反响,以株型塑造为手段的理想株型育种模式得以广泛开展[15-16]。作物的株型决定其受光水平、吸收水肥能力、抗逆能力等,进而决定其生物产量和经济产量,因此不少学者从源库关系[17]、表型可塑性[18]、水分利用效率[19-20]、竞争能力[21]、资源分配格局[22]及作物的生理性状[23-24]等方面出发,对作物的株型与产量之间的关系做了研究,探讨高产的途径,取得一定成效。然而,生长冗余现象的存在很大程度上限制了作物产量的提高[25-26]。
所谓生长冗余,盛承发认为是生物适应波动环境的一种生态对策[25]。当环境改善或经人类支持与保护后,作物这种固有的冗余特性变成了一种浪费和负担,对高产不利,通过栽培或育种手段减少冗余便可高产。目前大量的生长冗余仍存在于作物的营养器官中,理想株型还没有真正实现,且不同地区的研究,对理想株型的认识依然存在很大分歧,因此不少学者对理想株型育种前途提出质疑。旱地小麦如何实现高产目标?旱地小麦的育种出路何在?对于这些困惑,理解小麦株型的演变趋势及与生长冗余的互作关系等理论问题就显得更为重要。本文对旱地小麦理想株型研究进展进行了回顾,同时又以过去20年黄土高原雨养农业区的观测数据为主进行集成分析,将产量分别与根系生物量、地上生物量和株高等指标进行回归分析,对旱地小麦生长冗余大小的演变进行了讨论。本研究在综述前人已有研究进展的基础上,通过集成分析,为旱地小麦理想株型演变、个体竞争能力、大小不整齐性、物质和能量分配及籽粒产量形成等之间的关系提供更多的认识。
1 旱地小麦理想株型研究进展
对作物的人工选择自一万年前就已经开始了[27-28]。在长期的人工选择过程中,人们总是有意或无意地改变作物的株型以获得较高的产量。我国早在1400年前,《齐民要术》一书就有关于“早熟者苗短而收多, 晚熟者苗长而收少”的记载[29]。在20世纪20年代,Doughty[30]提出的“最佳合成体”可以看作是理想株型概念的雏形,他认为通过适当杂交的方法和产量因素的最佳组合,把各种高产性状聚集在一起,从而得到高产。旱地小麦从野生近缘种向现代品种进化的漫长过程中,野生一粒小麦(T.boeoticum)、野生二粒小麦(T.dicoccoides)、拟斯卑尔脱小麦(T.speltoides)、粗山羊草(Ae.tauschii)和阿拉拉特小麦(T.araraticum)等经过自然轮回杂交和驯化,逐渐演变成了目前广为人类栽培利用的多倍体小麦[31]。其株型也经历了从“高根冠比、低收获指数、高冠幅/株高比、小穗和种子扁长”等“杂草型”向“低根冠比、高收获指数和高粒叶比及小冠幅/株高、大穗多粒”的作物型的演变[32-33]。
株型既是品种特性,又受栽培措施的影响,由于不同生态区的环境条件不同,获得高产的生育模式就可能不同。在生产实践中,理想株型是相对的,因此有人提出“动态模型”的概念,假定一种作物只有一种理想株型未免太过局限[34-35],理想株型应随着生育期进程及环境的改变而不断发生变化,这对理想株型有了更进一步的认识。目前对小麦理想株型的构建,多侧重从叶型、节间性状、根型和穗型等方面进行研究,主要研究冠层的形态结构和根系生长的时空变异性[36],通过对这些性状的改造以构建出小麦的理想株型。刘兆晔认为理想株型应有良好的透光性、遮光性和抗旱保水性,能适应小麦生长的各个生育阶段[37]。张文宇在分析株型指标变化趋势的基础上,通过冠层切割的方法模拟了叶面积指数的分层动态变化,并通过对形态指标的归类分析,构建了综合性株型构成指数,综合体现了叶型和茎型的动态变化[38]。
表型可塑性的研究为理想株型的研究提供了强有力的理论支持,表型可塑性被认为是植物在可变环境中最大适合度的功能反应[34]。从形态可塑性上来研究植物的理想株型塑造及生长策略具有重要的意义[35,39-41]。可塑性研究包括与资源获取直接相关的地上和地下部分的特征,如株高、叶面积大小、分蘖的有无及根长度、根的空间分布等[42-45]。植物表型可塑性能够使植物把资源最佳地分配到不同器官,以避免或降低相邻植物的竞争[46-48]。特定性状的可塑性本身可以遗传,也可以接受选择而发生进化[49]。故作物表型可塑性的研究,推进了理想株型研究的进展。
旱地谷类作物理想株型的塑造过程,是一个不断剔除生长冗余的过程,尤其是剔除根系的冗余。在干旱半干旱地区,旱地春小麦具有根系生物量小、根条数少、根冠比低的根系构型,这些特点使现代春小麦在早期营养生长过程中能节约用水而保留一定的土壤水分,以满足花期和灌浆期水分需求,从而提高作物产量[50]。因作物与环境互作关系的复杂性,以及作物间竞争存在的必然性,作物的“理想株型”到目前还未实现。“理想株型”的概念虽然定义的是个体植株,但强调的是基于群体生产的思想来塑造个体植株。作物的生产是一个群体过程[15],今后对旱地小麦理想株型的选择应围绕群体产量最优这一重要标准,“理想株型”的实质应是“群体株型”的最优化状态。
2 基于生长冗余理论的旱地小麦株型塑造
20世纪80年代对全国小麦调查结果表明[51],全国各麦区小麦品种主要性状变化有:1)植株由高变矮;2)籽粒由小变大;3)成熟期越来越早。小麦株型变得越来越合理。在黄土高原雨养农业区,农业生产的主导限制因子是水,个体间竞争主要表现为根系生物量大小的竞争,因此提高小麦产量首先要解决的问题是减少根系的资源消耗,提高水分利用效率。随着集水农业的迅猛发展,田间水分条件得到极大改善,过去一直占优势的大根系古老品种如和尚头和金包银逐渐被新一代品种所取代。目前在黄土高原培育出许多小根系品种,适应当地的生态环境,获得了较高的产量。由此看出人们已有意识地减少株型上的冗余,促进了黄土高原旱地小麦理想株型的塑造。本文以近十几年来40余篇旱地小麦研究方向的参考文献为基础,分析了相关形态指标与产量的关系,以期为旱地小麦理想株型塑造提供理论依据。
2.1 根系生长冗余研究进展
半干旱雨养农业区,植物往往产生庞大的根系,这是自然选择的结果,也是进化上稳定的生长对策(ESS)[11]。人们往往认为深入、广布且多分枝的根系是抗旱作物所必需的,故大根系被农学家和育种学家所偏爱,因此半干旱雨养农业区早期作物具有庞大的根系,不仅是自然选择的结果,也有人工选择的干预。但大量实践证明,大根系作物并没有带来高产,相反,一些小根系作物在半干旱区通常具有较高的产量。作物生产的群体观念促使人们追求群体产量最优。作物群体生活中的个体,其最大的根系有两层含义:一是使群体产量达到最大,即农业生产上的最佳;二是使个体竞争力达到最大,即进化上的最佳[11]。而人们在很长一段时间内都未意识到两者之间的差异,这也正是旱区抗旱育种表现为抗旱有余,丰产不足的原因所在[6]。
Huang[52]在研究旱地小麦从野生近缘种到六倍体小麦的进化中,发现根系生物量有先增大后减小的演变趋势。从二倍体到四倍体进化中,以自然选择为主导,以提高个体竞争力达到个体最优,趋向达到进化上的最优。这表现为植株的根系增大,根冠比增加,以人类利益而言则出现了根系冗余。从四倍体到六倍体,人工选择为主导,注重的是群体效应,成为弱竞争者的植株达到产量上的最优。这表现为小根系,根冠比减小,减少了根系冗余。也有人研究发现人工育种过程中小麦根系一直呈减小趋势[23]。尽管演变过程中根系大小有着复杂的变化趋势,但是总体趋于减小。小麦进化中根系虽减小了,而水分利用效率、对营养元素的吸收效率却显著提高[53-54]。
六倍体春小麦随年代推进逐渐呈现根条数减少、根系下扎深、根活性吸收面积比(根系活性吸收面积/根系总吸收面积)增加、种子根中央大导管直径减小的趋势[50]。这些特点均利于现代春小麦在生育后期利用土壤储水,保证了作物产量。早在1983年苗果园对黄土高原旱地冬小麦根系生长规律研究后指出,下层根量对旱地小麦后期充分利用深层水十分重要[31]。因Donald对理想株型根系的描述是小根系,使人们对根系冗余的认识有了本质的改变,更多的人工选择倾向于小根系品种的培育,小根系品种也为旱农区的作物产量提高带来更大收益。张荣[55]在黄土高原连续两年的实验结果显示,在极端干旱年份(1995年),古老品种和与现代品种在产量上没有显著差异(P>0.05),而在正常降雨年份(1996年),古老品种产量显著低于现代品种(P<0.05)。对各品种进行回归分析:根系大小与籽粒产量、生物量均呈显著负相关(R=0.9065*,R=0.9860**)。古老品种在根系上形成冗余,现代品种减少了根系冗余,故在产量上差异突显。孙国钧[10]对小根系品种在干旱半干旱区具有高产潜力做了如下解释:其一,减少的根系生物量转移到地上部分,提高了小麦的繁殖分配;其二,减少了根系冗余,降低了作物个体间的竞争能力,提高了群体的水分利用效率,将有限的土壤水分充分利用。

图1 旱地小麦根系生物量与产量的散点分布图Fig.1 Scatters distribution of the root biomass and the yield of dryland wheat
图1为各试验处理下旱地小麦根系生物量与产量的散点分布图。从图中可看出在黄土高原干旱半干旱区,产量并未随根系生物量的增加而一直增加。在根系生物量较大时,产量反而有下降趋势,这与Zhang[9]认为旱地作物中普遍存在根系上的冗余结论一致。为了验证根系冗余的存在以及减少冗余可以增加产量,大量的去根试验也广泛开展[56-59]。试验证明减少根系的冗余可以显著提高作物产量。因自然选择压力的存在,长期的人工选择仍不能完全消除根系的冗余,理想株型与生长冗余之间不是简单的此消彼长的关系。旱地小麦育种过程中根系的演变是一个逐渐消除冗余但又无法完全消除的过程,人工选择的干预,促进了这一演变过程。
2.2 旱地小麦株高性状演变
矮杆小麦的出现使小麦产量大幅度提高,曾被誉为一场绿色革命[15]。人们培育矮杆小麦的初衷是为了抗倒伏,却意想不到的获得了高产。有研究指出在70a时间内,英国小麦品种,株高从130cm降到70cm左右,收获指数由34%提高到50%,这个数值可能还会继续提高到62%[60]。墨西哥的硬质小麦从1969年到1984年间其收获指数从0.25上升到0.45,株高有较大的降低[61]。中国60年代小麦平均株高比50年代下降了3.3 cm,70年代比60年代下降了7.5 cm,80年代比70年代又下降了8.3 cm[51]。在当前高精度的农田生产条件下,产量的提高主要因为资源分配格局发生变化,株高降低释放的生物量用于籽粒的建设而增加产量。进化过程中,种群中高秆小麦对光资源的竞争较其周围矮杆小麦有着绝对优势,因此高秆小麦会被逐渐选择下来,而根据生活史对策理论,有限资源向某一功能器官分配增多,就必然使分配到其它功能器官的资源相应减少。当小麦获得的收益(向茎秆做更多投资而获得较高竞争力)不足以弥补对繁殖分配造成的损害时,小麦株高便不再增加,最终达到进化上的稳定(ESS)。而这种策略下的株高,从农业生产的角度来讲是往往冗余的。
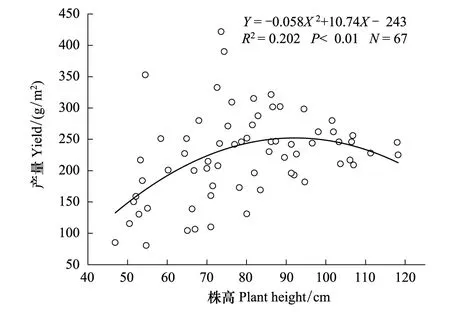
图2 旱地小麦株高与产量的散点分布图Fig.2 Scatters distribution of the height and the yield of dryland wheat
在有限的资源环境下,较大的营养体可能对水分、养分、光照和CO2的吸收和同化利用能力较高,但营养生长越大,相应的蒸腾耗水会越多,甚至引起倒伏[62]。人工培育旱地小麦理想株型过程中,倾向于矮秆品种以减少株高的冗余。从六倍体的古老品种到现代品种,株高呈逐渐降低趋势。但应当指出小麦株高并不是越矮越好,过矮的小麦株型,冠层低而密集,通风透光不良,往往后期生育和成熟不良,籽粒饱满度差[51]。现在育种过程中过多降低株高以增加繁殖分配的做法并不可取,因为经过无数代的人工育种选择,株高已达到一个最优的高度[63-64]。傅兆麟[65]研究认为理想株型株高平均值为(79.1±6.1) cm,变幅范围为68.0—91.0 cm。徐沛然[66]研究指出超高产基因型株高不能超过85cm,以80 cm较好。李朴芳[33]总结前人的研究认为旱地六倍体小麦株高最好控制在80cm—100 cm之间。可以看出理想株型中的株高并不是固定不变的,而是在某一最优的范围(如80—90 cm)内有所变动,这可能与研究区生态环境的不同有关。
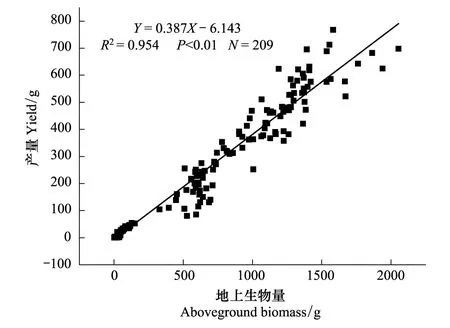
图3 旱地小麦地上生物量与产量的散点分布图Fig.3 Scatters distribution of the above-biomass and the yield of dryland wheat
从图2看出,产量并不是随株高的增加而一直增加,在株高超过95 cm后,随着株高的增加产量有下降的趋势。对旱地小麦株高样本(N=67)统计分析,株高低于80 cm的占50.75%,80—100 cm的占32.84%,而株高大于100 cm的样本占总样本的16.42%,说明旱地小麦的株高还没有达到理想状态。干旱环境限制了小麦的株高,在改良环境以促进株高方面还有提高产量的发展空间。如图3所示,旱地小麦地上生物量与产量呈显著正相关(R2=0.954,P<0.01),现阶段旱地小麦在产量提高上还有很大的发展潜力,提高地上生物量以提高产量的做法是可取的,这与张荣[12]研究结果一致。
种群个体大小不整齐度代表作物在胁迫条件下的竞争状况,是种群对干旱环境的一种适应对策,但是以降低产量为代价的。随着干旱胁迫的增大,种群个体大小不整齐性增加,加剧了个体间对水分的竞争[67],导致生长冗余的发生。提高作物群体株高大小整齐度,也将是今后旱地小麦理想株型演变的方向[24,68]。
2.3 旱地小麦分蘖性状及籽粒大小演变
在理想株型演变中,分蘖数一直是人们研究的热点。盛承发[25]认为小麦在分蘖上的冗余是长期进化过程中形成的一种生态对策,可以减少外界环境不利变化对其繁殖的影响,其中无效分蘖在正常情况下也许对产量无甚贡献,但在遭受严重伤害等异常情况下,可减轻灾害损伤,减小绝种风险。在干旱地区低分蘖的小麦品种,由于减少了分蘖上的冗余,更多能量供给主茎,提高了收获指数[69]。
旱地小麦从六倍体古老品种到现代品种的选育过程中,分蘖数有减少的趋势。分蘖数的减少,可以使主茎接受更多的资源。然而目前小麦仍有一定数量的分蘖存在,且高达50%—70%的分蘖是无效分蘖[25],这对于产量的提高是一个很大的阻碍[70]。分蘖与主茎存在资源竞争关系,过多的分蘖意味着有更多的库容量,若总库容量过大,势必造成部分分蘖不能成功发育,对资源形成浪费,出现生长冗余。马守臣[20]研究认为小麦在分蘖上存在对水分利用不利的生长冗余,去除无效分蘖对产量没有显著影响,而减少这些冗余则可以节约用水、提高作物的水分利用效率。黄土高原旱作小麦种植区,因作物生育期与丰雨季节存在严重的时空错位,作物生长主要依赖上一生长季的土壤储水。如果在有效分蘖终止期,采用人工措施或化学调控减少无效分蘖的生长,从而减少其对土壤水分的消耗,以保留更多水分供后期使用,这对提高作物群体产量具有重要意义。这也说明旱地小麦的理想株型应具有分蘖数少的特性。

图4 旱地小麦千粒重和收获指数之间的回归关系Fig.4 Scatters distribution of the thousand kernel weight and the harvest index of dryland wheat
小麦的产量构成因素包括单位面积穗数、每穗粒数和粒重。70年代以来,中国小麦育种工作者广泛开展了小麦大粒育种研究。随着时间推进,小麦的千粒重有明显提高,小麦千粒重增加的百分比50年代为最高,增幅达12.42%,60—80年代增幅依次为9.3%、4.9%和1.7%,千粒重的增幅逐年代有所减慢[51]。图4为旱地小麦千粒重和收获指数之间的散点分布图,近年来干旱地区的实验结果表明,千粒重和收获指数呈显著正相关(P<0.01),千粒重的提高可以明显地提高收获指数,这意味着更多的能量转移至籽粒,资源分配格局发生了改变。
3 旱地小麦株型演变与生长冗余的互作关系
早在19世纪,达尔文就指出作物从野生种到现代品种进化过程中,人工选择和自然选择目标具有异向性[71]。自然选择侧重个体竞争力的提高,人工选择的目的则是改善作物的群体产量。根据达尔文进化学说,若个体采取某一种策略,若由此获得的收益大于付出的代价,那么优化竞争能力的资源分配策略是能够被自然选择下来的。所以长期的竞争将会使作物保留一些从农业生产角度来看完全是冗余的性状。然而农学工作者在很长一段时间内都没有意识到这种竞争对农作物产量带来的负面影响。就作物的高产高效而言,冗余的产生是不利的,而从作物本身的自然属性出发,生长冗余的特性是长期进化过程中形成的适应环境波动、提高竞争能力的生态对策,是长期自然选择和进化的结果[20,26]。由于自然选择和人工选择的矛盾性及环境的多变性,作物株型演变与生长冗余的互作关系变得更加复杂。在育种过程中,就需权衡人类的利益(弱竞争,无冗余,高繁殖分配)和作物生存的利益(强竞争,有冗余,低繁殖分配),以达到群体产量的最大化。张大勇从进化的角度认为理想株型与生长冗余之间存在着博弈关系。他基于一系列假设,论证了生长冗余广泛存在于作物中(图5)。
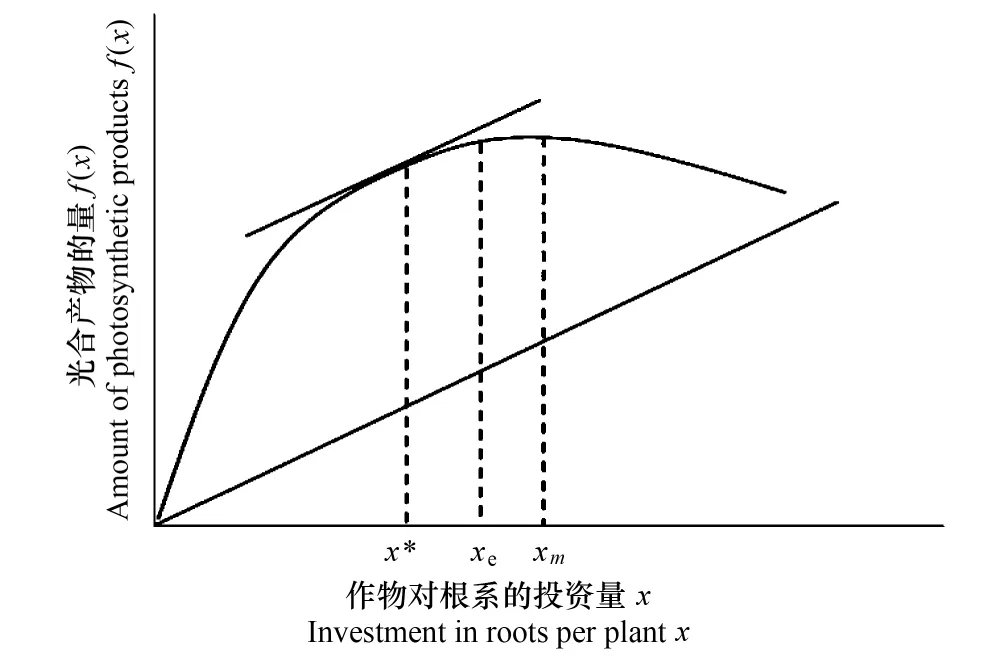
图5 具有相同基因型的作物在水资源限制环境下光合产物与根系投资量的关系Fig.5 The relationship between the total amount of photosynthetic products and the root biomass under the water limited conditions曲线为光合产物f(x),直线为根系的投资量x。光合产物的量在xm处最大,总产量在x*处最大; xe 为实际生产中的状态,xe与x*之间的差值即为根系的生长冗余(根据Zhang D Y, 1999, 稍作修改[9]
由图5可以看出x*为人工育种的最终目的,代表的是理想株型状态,xm是自然选择的结果,代表作物具有较好的竞争能力。xe为实际生产中作物对根系的投资量。因作物基因型的不同、环境的复杂程度及种内竞争的存在,xe与x*之间的差异始终不能消除(即xe不能达到x*状态),理想株型演变的过程就是xe在与两者(xm与x*)的博弈中,以求得产量最大化。
在长期的进化过程中,单位面积作物的生物量已没有太大变化[55,72],产量的提高主要靠地上生物量和收获指数的提高,这说明能量从其他器官转移到籽粒上来,营养器官中的生长冗余减少,资源分配格局发生改变。而旱地小麦株型演化过程中,株型的演变趋势并不是不变的,如旱地小麦的株高并不是越矮越好,根系生物量也不是越小越好,株型上冗余的能量不能完全释放出来用于籽粒的建设。可见,一定程度的冗余存在是塑造理想株型的基础。
针对作物中冗余的存在,农业生产中有人提出了冗余度的概念,认为各种冗余都有一定的限度,其大小可以用冗余度表示[73]。最佳生长冗余度[74]是指在一定的栽培管理条件下,农作物存在着一个与最高产量相对应的生长冗余度。生产中如通过各种栽培管理措施使农作物的生长冗余度达到最佳值,就能获得高产。但究竟多大程度的冗余存在能够使产量达到最优?过多的生长冗余造成了资源浪费,而通过人工调节过度消除生长冗余可能会影响作物的生长发育而不能获得高产。同时,旱地小麦小根系品种在产量上的优势也是相对于中度干旱情况而言的,大根系品种在水分充足条件下,也突出了自身的优势。因此作物最佳冗余度的确定,不仅取决于作物间的竞争程度、人类利益与作物生存利益之间的权衡,还必须考虑复杂的环境背景、作物的物候等因素。不同的水分状况、光照条件及物候期,所对应的最佳冗余度都可能存在差异,理想株型的塑造标准也不同,这也是理想株型与生长冗余互作关系复杂的一面。
4 旱地小麦理想株型演变和生长冗余研究的发展方向
小麦在中国已有4000多年的种植历史,各地所形成的品种类型、株型不仅适应当地的气候、土壤等自然条件,也与当地的耕作制度相适应。干旱半干旱地区严酷的生态环境,一直制约着小麦产量的提高。旱地小麦产量的提高不仅取决于所有产量因素的协调发展[65],还取决于当地的环境因素。环境差异是引起我国旱地春小麦产量差异的主要原因,环境间、基因型与环境互作引起的变异远远大于基因型间的变异,因此应重视外界环境的改良作用[62]。
基于株型改良提高产量的措施被广泛应用到生产实践中。对株型的改良,主要去改良自然选择对其个体有利,却有害于群体产量提高的性状(如大根系、高茎秆);而那些在任何情况下的改良对作物个体和群体都有利的性状(如单位叶面积的光合作用速率),是不需要过多注意的,这是因为长期的自然进化已使这些性状表现的很完美[9,75]。理想株型理论的提出及育种实践,使小麦产量的提高跃上了一个新的台阶。
关于旱地小麦株型演变和生长冗余研究发展的方向,需要有两点认识:“理想株型”不能真正实现,生长冗余无法完全消除;且一定程度生长冗余的存在是理想株型发生的基础。在干旱区以株型塑造为主提高作物产量的过程中应注意:(1)由弱竞争者组成的种群,即个体将更多资源投入到繁殖器官而非更多地投入到竞争器官,将有助于收获指数的提高。因此,育种过程应是降低个体竞争能力的过程。(2)籽实产量是收获指数与生物量的乘积。要实现单位面积较高的籽实产量,应同时提高收获指数与地上生物量(图6)。理想株型育种过程中虽然无法完全消除生长冗余,但可以无限制的接近理想株型状态,只要意识到生长冗余存在的必然性和减少冗余可以获得高产的重要性[76],就一定会找到各种行之有效的生产措施以提高产量。理想株型的塑造是一个漫长的进化过程,它需要人工选择的不断干预。目前旱地农业处于从低产向中产发展阶段,提高产量的潜力仍然很大[77],株型育种的潜力还没有完全挖掘出来,基于株型改良方式以提高旱地小麦产量的思想,具有广阔的发展前景。
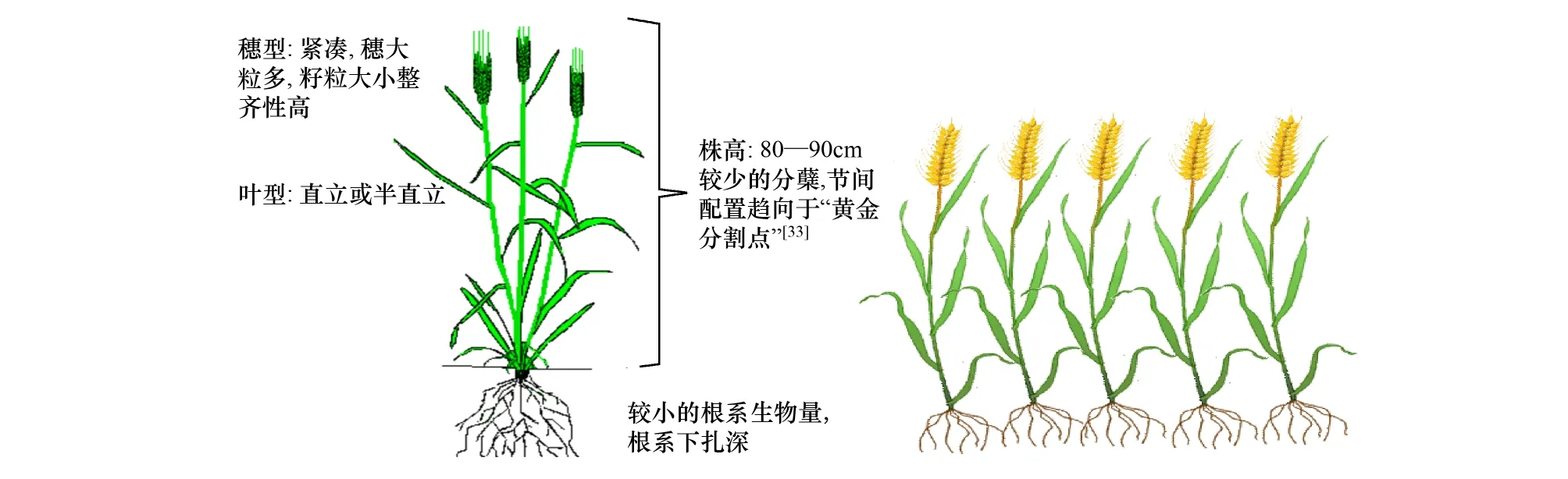
图6 旱地小麦理想株型示意图及群体效果Fig.6 The weak competitor ideotype for dryland wheat in a crop community
致谢:感谢兰州大学生命科学学院张荣博士、西澳大利亚大学农学院Neil C. Turner教授对本文写作的帮助。
[1] Sivapalan S, O′Brien L, Ortiz-Ferrara G, Hollamby G J, Barclay I, Martin, P J. Yield performance and adaptation of some Australian and CIMMYT/ICARDA developed wheat genotypes in the West Asia North Africa(WANA) region. Crop and Pasture Science, 2001, 52(6): 661-670.
[2] Manschadi A M, Christopher J, deVoil P, Hammer G L. The role of root architectural traits in adaptation of wheat to water-limited environments. Functional Plant Biology, 2006, 33(9): 823-837.
[3] Richards R A. Physiological traits used in the breeding of new cultivars for water-scarce environments. Agricultural Water Management, 2006, 80(1/3): 197-211.
[4] Olivares-Villegas J J, Reynolds M P, McDonald G K. Drought-adaptive attributes in the Seri/Babax hexaploid wheat population. Functional Plant Biology, 2007, 34(3): 189-203.
[5] Manschadi A M, Hammer G L, Christopher J T, de Voil P. Genotypic variation in seedling root architectural traits and implications for drought adaptation in wheat (TriticumaestivumL.). Plant and Soil, 2008, 303(1/2): 115-129.
[6] 张娟, 谢惠民, 张正斌, 徐萍. 小麦抗旱节水生理遗传育种研究进展. 干旱地区农业研究, 2005, 23(3): 231-238.
[7] 刘洪升, 李凤民. 水分胁迫下春小麦根系吸水功能效率的研究. 西北植物学报, 2003, 23(6): 942-948.
[8] Passioura J B. Roots and drought resistance. Agricultural Water Management, 1983, 7(1/3): 265-280.
[9] Zhang D Y, Sun G J, Jiang X H. Donald′s ideotype and growth redundancy: a game theoretical analysis. Field Crops Research, 1999, 61(2): 179-187.
[10] 孙国钧, 张荣, 张大勇. 干旱、半干旱区作物抗旱性和育种. 兰州大学学报: 自然科学版, 2001, 37: 53-59.
[11] 张大勇, 姜新华, 赵松岭, 段舜山. 半干旱区作物根系生长冗余的生态学分析. 西北植物学报, 1995, 15(5): 110-114.
[12] 张荣, 孙国钧, 张大勇. 干旱、半干旱地区作物育种的困惑与出路. 西北植物学报, 2000, 20(6): 930-935.
[13] Tambussi E A, Nogués S, Ferrio P, Voltas J, Araus J L. Does higher yield potential improve barley performance in Mediterranean conditions? A case study. Field Crops Research, 2005, 91(2/3): 149-160.
[14] Fleury D, Jefferies S, Kuchel H, Langridge P. Genetic and genomic tools to improve drought tolerance in wheat. Journal of Experimental Botany, 2010, 61(12): 3211-3222.
[15] 赵松岭, 李凤民, 张大勇, 段舜山. 作物生产是一个种群过程. 生态学报, 1997, 17(1): 100-104.
[16] 王永锐. 作物高产群体生理. 北京: 科学技术文献出版社, 1991: 1-10.
[17] 傅兆麟, 刘东华. 小麦源 库关系的研究. 安徽农业科学, 2005, 33(8): 1477-1479.
[18] 朱志红, 王刚. 燕麦表型可塑性与繁殖分配的研究. 兰州大学学报: 自然科学版, 2002, 38(1): 76-83.
[19] 孙国钧, 张荣, 李凤民, 张大勇. 作物抗旱性、水分利用与籽粒产量的形成. 资源科学, 1999, 21(5): 31-36.
[20] 马守臣, 徐炳成, 李凤民, 黄占斌. 冬小麦(Triticumaestivum)分蘖冗余生态学意义以及减少冗余对水分利用效率的影响. 生态学报, 2008, 28(1): 321-326.
[21] 董珑丽, 魏茶花, 马晓娟, 张荣. 春小麦竞争能力与产量的关系. 生态学报, 2007, 27(10): 4203-4208.
[22] 郭安红, 魏虹, 李凤民, 赵松岭. 土壤水分亏缺对春小麦根系干物质累积和分配的影响. 生态学报, 1999, 19(2): 179-184.
[23] 许宝泉, 张荣, 张大勇, 孙国钧. 旱农区春小麦根系生长冗余与碳平衡关系的研究. 西北植物学报, 2003, 23(6): 958-963.
[24] 孟宪芝. 干旱对三个春小麦品种根系呼吸及产量的影响[D]. 兰州: 兰州大学, 2008.
[25] 盛承发. 生长的冗余——作物对于虫害超越补偿作用的一种解释. 应用生态学报, 1990, 1(1): 26-30.
[26] 张荣, 孙国钧, 李凤民. 冗余概念的界定与冗余产生的生态学机制. 西北植物学报, 2003, 23(5): 844-851.
[27] Tanno K, Willcox G. How fast was wild wheat domesticated? Science, 2006, 311(5769): 1886.
[28] Nevo E, Chen G X. Drought and salt tolerances in wild relatives for wheat and barley improvement. Plant, Cell and Environment, 2010, 33(4): 670-685.
[29] 魏燮中. 我国古农书关于禾谷类作物株型及其生态表现的记载. 农业考古, 1987, (1): 292-299.
[30] Doughty L R, Engledow F L. Investigations on yield in the cereals: V. A study of four wheat fields: the limiting effect of population-density on yield and an analytical comparison of yields. The Journal of Agricultural Science, 1928, 18(2): 317-345.
[31] 马元喜. 小麦的根. 北京: 中国农业出版社, 1999: 10-12.
[32] Dubcovsky J, Dvorak J. Genome plasticity a key factor in the success of polyploid wheat under domestication. Science, 2007, 316(5833): 1862-1866.
[33] 李朴芳, 程正国, 赵鸿, 张小丰, 李冀南, 王绍明, 熊友才. 旱地小麦理想株型研究进展. 生态学报, 2011, 31(9): 2631-2640.
[34] Coleman J S, McConnaughay K D M, Ackerly D D. Interpreting phenotypic variation in plants. Trends in Ecology and Evolution, 1994, 9(5): 187-191.
[35] 孙儒泳. 生活史对策. 生物学通报, 1997, 32(5): 2-4.
[36] 张晓萍, 杨慎骄, 张笑培, 白芳芳, 王和洲. 不同播期冬小麦株型构建及其生育特征. 应用生态学报, 2013, 24(4): 915-920.
[37] 刘兆烨, 于经川, 姜鸿明, 辛庆国, 刘克宁, 赵明. 小麦理想株型的探讨. 中国农学通报, 2010, 26(8): 137-141.
[38] 张文宇, 汤亮, 姚鑫锋, 杨月, 曹卫星, 朱艳. 基于过程的小麦株型指标动态模拟. 中国农业科学, 2012, 45(12): 2364-2374.
[39] Sultan S E. Phenotypic plasticity and plant adaptation. Acta Botanica Neerlandica, 1995, 44: 363-383.
[40] Sultan S E. Phenotypic plasticity for plant development, function and life history. Trends in Plant Science, 2000, 5(12): 537-542.
[41] Sultan S E. Promising directions in plant phenotypic plasticity. Perspectives in Plant Ecology, Evolution and Systematics, 2004, 6(4): 227-233.
[42] Caradus J R, Hay M J M, Mackay A D, Thomas V J, Dunlop J, Lambert M G, Hart A L, Van Den Bosch J, Wewala S. Variation within white clover (TrifoliumrepensL.) for phenotypic plasticity of morphological and yield related characters, induced by phosphorus supply. New Phytologist, 1993, 123(1): 175-184.
[43] Wool D, Hales D F. Phenotypic plasticity in Australian cotton aphid (Homoptera: Aphididae): Host plant effects on morphological variation. Annals of the Entomological Society of America, 1997, 90(3): 316-328.
[44] Cordell S, Goldstein G, Mueller-Dombois D, Webb D, Vitousek P M. Physiological and morphological variation in Metrosideros polymorpha, a dominant Hawaiian tree species, along an altitudinal gradient: the role of phenotypic plasticity. Oecologia, 1998, 113(2): 188-196.
[45] 王天慧. 植物表型可塑性及生活史对策研究 [D]. 吉林: 东北师范大学, 2006.
[46] Brewer J S. Short-term effects of fire and competition on growth and plasticity of the yellow pitcher plant,Sarraceniaalata(Sarraceniaceae). American Journal of Botany, 1999, 86(9): 1264-1271.
[47] Via S, Hawthorne D J. The genetic architecture of ecological specialization: correlated gene effects on host use and habitat choice in pea aphids. The American Naturalist, 2002, 159(S3): S76-S88.
[48] Weiner J. Allocation, plasticity and allometry in plants. Perspectives in Plant Ecology Evolution and Systematics, 2004, 6(4): 207-215.
[49] 陆霞梅, 周长芳, 安树青, 方超, 赵晖, 杨茜, 颜超. 植物的表型可塑性、异速生长及其入侵能力. 生态学杂志, 2007, 26(9): 1438-1444.
[50] 李话, 张大勇. 半干旱地区春小麦根系形态特征与生长冗余的初步研究. 应用生态学报, 1999, 10(1): 26-30.
[51] 董玉琛, 郑殿生. 中国小麦遗传资源. 北京: 中国农业出版社, 1999: 58-112.
[52] Huang M L, Deng X P, Zhao Y Z, Zhou S L, Inanaga S, Yamada S, Tanaka K. Water and nutrient use efficiency in diploid, tetraploid and hexaploid wheats. Journal of Integrative Plant Biology, 2007, 49(5): 706-715.
[53] Zhang S Q, Shan L, Deng X P. Change of water use efficiency and its relation with root system growth in wheat evolution. Chinese Science Bulletin, 2002, 47(22): 1879-1883.
[54] 李秧秧, 张岁岐, 邵明安. 小麦进化材料水分利用效率与氮利用效率间相互关系. 应用生态学报, 2003, 14(9): 1478-1480.
[55] 张荣, 张大勇. 半干旱区春小麦不同年代品种根系生长冗余的比较实验研究. 植物生态学报, 2000, 24(3): 298-303.
[56] 王振宇, 吕金印, 李凤民, 徐炳成. 断根对冬小麦竞争能力与产量的影响. 植物生态学报, 2007, 31(2): 300-304.
[57] 石岩, 位东斌, 于振文, 于松烈. 深耘断根对旱地高产小麦花后旗叶衰老及产量的影响. 麦类作物学报, 2000, 20(3): 67-69.
[58] 侯慧芝, 黄高宝, 郭清毅, 王利立. 干旱灌区冬小麦根系的生长冗余. 生态学杂志, 2007, 26(9): 1407-1411.
[59] Ma S C, Li F M, Xu B C, Huang Z B. Effect of lowering the root/shoot ratio by pruning roots on water use efficiency and grain yield of winter wheat. Field Crops Research, 2010, 115(2): 158-164.
[60] Austin R B, Bingham J, Blackwell R D, Evans L T, Ford M A, Morgan C L, Taylor M. Genetic improvements in winter wheat yields since 1900 and associated physiological changes. The Journal of Agricultural Science, 1980, 94(3): 675-689.
[61] Waddington S R., Osmanzai M, Yoshida M, Ransom J K. The yield of durum wheats released in Mexico between 1960 and 1984. Journal of Agricultural Science, 1987, 108(2): 469-477.
[62] 常磊, 柴守玺, 杨德龙, 鲁清林. 我国旱地春小麦产量及主要农艺指标的变异分析. 应用生态学报, 2010, 21(11): 2821-2829.
[63] Fischer R A, Quail K J. The effect of major dwarfing genes on yield potential in spring wheats. Euphytica, 1990, 46(1): 51-56.
[64] Miralles D J, Slafer G A. Individual grain weight responses to genetic reduction in culm length in wheat as affected by source-sink manipulations. Field Crops Research, 1995, 43(2/3): 55-66.
[65] 傅兆麟. 小麦超高产基因型的株型结构问题. 云南农业大学学报, 2007, 22(1): 17-22.
[66] 徐沛然, 牟春生. 小麦超高产育种几个问题的探讨 // 何中虎, 张爱民. 中国小麦育种研究进展. 北京: 中国科学技术出版社, 2002: 119-125.
[67] Pan X Y, Wang G X, Yang H M, Wei X P. Effect of water deficits on within-plot variability in growth and grain yield of spring wheat in northwest China. Field Crops Research, 2003, 80(3): 195-205.
[68] Song L, Li F M, Fang X W, Xiong Y C, Wang W Q, Wu X B, Turner N C. Soil water availability and plant competition affect the yield of spring wheat. European Journal of Agronomy, 2009, 31(1): 51-60.
[69] Elhani S, Martos V, Rharrabti Y, Royo C, García del Moral L F. Contribution of main stem and tillers to durum wheat (TriticumturgidumL. var.durum) grain yield and its components grown in Mediterranean environments. Field Crops Research, 2007, 103(1): 25-35.
[70] Donald C M. The breeding of crop ideotype. Euphytica, 1968, 17(3): 385-403.
[71] Darwin C. The Variation of Animals and Plants under Domestication. London: John Murray, 1868.
[72] 董珑丽. 半干旱区春小麦竞争能力与产量、密度关系研究 [D]. 兰州: 兰州大学, 2007.
[73] 党承林, 王崇云. 生态系统的能量冗余与热力学第二定律. 生态学杂志, 1999, 18(1): 53-58.
[74] 韩明春, 吴建军, 王芬. 冗余理论及其在农业生态系统管理中的应用. 应用生态学报, 2005, 16(2): 375-378.
[75] Passioura J B. The interaction between the physiology and the breeding of wheat // Evans L T, Peacock W W J, Frankel O H, eds. Wheat Science-Today and Tomorrow. Cambridge: Cambridge University Press, 1981: 191-201.
[76] 张大勇, 姜新华, 赵松岭. 再论生长的冗余. 草业学报, 1995, 4(3): 17-22.
[77] 山仑. 植物抗旱生理研究与发展半旱地农业. 干旱地区农业研究, 2007, 25(1): 1-5.
Plant ideotype and growth redundancy in dryland wheats
WANG Jianyong1, LI Pufang1, CHENG Zhengguo1, ASFA Batool1, LÜ Guangchao1, ZHU Ying1,2, XIONG Youcai1,*
1StateKeyLaboratoryofGrasslandandAgro-Ecosystems,InstituteofAridAgroecology,SchoolofLifeSciences,LanzhouUniversity,Lanzhou730000,China2InstituteofBiology,GansuAcademyofSciences,Lanzhou730000,China
Since Donald proposed ideotype breeding in 1968, agronomists and breeders have endeavoured to develop ideotype-oriented breeding methods to improve the dryland wheat production. Several scholars have discussed methods to reduce growth redundancy and improve the ideotype of crops, and have made some progress on dryland wheat breeding strategies and model innovation for cultivation management. Nevertheless, the evaluation of crop ideotype is not the same in different ecological zones, and progress on a specific crop ideotype for a particular ecological zone is not universally accepted. In this paper, the recent progress in ideotype research for the dryland wheat is reviewed and the evolutionary trend in growth redundancy in roots, stems and tiller number studied. At the same time, based on the data that has been collected under rainfed agricultural systems on Loess Plateau over the past two decades, we conducted regression analysis between root biomass, above-ground biomass, height and yield. On the Loess Plateau, spring wheat still has a high root/shoot ratio and removal of some root biomass can improve seed production, implying that there is a high incidence of growth redundancy in modern crop cultivars. The excessive growth of individual organs results in decreased yields. Further ecological mechanism of growth redundancy and interaction between ideotype and growth redundancy were also discussed. Research has shown that growth redundancy can be gradually eliminated in ideotype succession process, but can′t be completely eliminated. A certain extent of the growth redundancy is the basic material for the ideotype. Over the long course of evolution from wild diploid and tetraploid, to modern hexaploid genotypes, spring wheat has been evolving from the high root/shoot ratio to the low root/shoot ratio, and this crop type could contribute to the increased yield of hexaploid wheat. We point out that the direction of dryland wheat ideotype breeding in the future should focus on the interaction between genotype of the wheat and its environment, reducing the intensity of competition among individuals and size inequality, promoting photosynthesis products that are deposited in the grain, and finally improving the population production. In summary, the interaction between a dryland wheat ideotype and growth redundancy is very complex. Selecting a dryland wheat ideotype should be based on the growth redundancy theory. Future breeding should be based on the basic principles of ecology, coupling the genotype and phenotype of dryland wheat, and then trade-off selection. The research focuses should move from research dealing with drought-resistance physiological genetics to the current ones dealing with crops ideotype. We believe that the theoretical and applied perspectives for the improvement of wheat production in the future will encompass the following: (1) The combination of the various morphological-physiological traits into one plant type; (2) The increase in the above-ground biomass and hence harvest index by selecting for small root systems and optimum resource allocation patterns; and (3) Selection and maintenance of weak competitors in population. Finally, the cooperative strategy that maximizes the population′s grain production, although clearly desirable in agriculture, is not evolutionarily stable.
dryland wheat; plant ideotype; growth redundancy; interaction; review
国家自然科学基金(31070372); 国家科技支撑计划(2012BAD14B10); 甘肃省科学院开发与应用基金项目(2012JK-03); 科技部国际合作项目(2013DFA30950); 国家星火计划项目(2012GA860003); 国家公益性行业(气象)科研专项(GYHY201106029-2); 中央高校基本科研业务费专项(lzujbky-2012-220)
2013-06-08;
日期:2014-05-16
10.5846/stxb201306081469
*通讯作者Corresponding author.E-mail: xiongyc@lzu.edu.cn
王建永,李朴芳,程正国,Asfa Batool,吕广超,祝英,熊友才.旱地小麦理想株型与生长冗余.生态学报,2015,35(8):2428-2437.
Wang J Y, Li P F, Cheng Z G, Asfa Batool, Lü G C, Zhu Y, Xiong Y C.Plant ideotype and growth redundancy in dryland wheats.Acta Ecologica Sinica,2015,35(8):2428-2437.
猜你喜欢
今日农业(2022年14期)2022-09-15
作物学报(2022年7期)2022-05-11
今日农业(2020年19期)2020-12-14
冰雪运动(2020年6期)2020-07-21
冰雪运动(2019年5期)2019-08-24
中国农业信息(2017年12期)2017-09-03
上海农业学报(2017年4期)2017-04-10
养生保健指南(2016年12期)2017-01-06
中国科技信息(2015年2期)2015-11-16
植物营养与肥料学报(2014年1期)2014-03-11
