
线蚓科分类学研究进展
2015-03-13 03:28蒋万祥谢志才
生态学报 2015年8期
陈 静,蒋万祥,沈 琦,谢志才,*
1 枣庄学院生命科学学院, 枣庄 277160 2 中国科学院水生生物多样性与保护重点实验室,水生生物研究所, 武汉 430072
线蚓科分类学研究进展
陈 静1,蒋万祥1,沈 琦2,谢志才2,*
1 枣庄学院生命科学学院, 枣庄 277160 2 中国科学院水生生物多样性与保护重点实验室,水生生物研究所, 武汉 430072
线蚓科隶属于环节动物门环带纲,迄今共记录32属650余种,是该纲的第二大科。它们广泛分布于土壤、海洋、淡水、河口和冰川等。其中,约2/3的线蚓科物种(近500种)为陆生种类,100余种仅分布在海洋中。尽管经历了200多年的探索,线蚓科仍然是认知最少的类群之一。尝试回顾人类对线蚓科环带动物分类学和系统发育学方面的认知历程和积累的知识体系,描述线蚓科关键的形态学分类特征以及线蚓科分类研究遇到的主要问题和障碍,展望了线蚓科分类学未来的研究方向。线蚓科的分类研究尚处于α分类阶段,体现在以物种探索为主和大量的已描述的物种需要验证等。而基于生物学物种概念(生殖隔离)的线蚓科物种探索,虽然有一些合理的逻辑解释,但缺乏严格的科学验证。线蚓科内多数属为复系,表明已建立的线蚓科分类系统仍然不能很好的反映线蚓科的自然进化历史。线蚓科分类面临的主要问题和障碍是未描述种类亟待发掘、已描述的物种需要验证、属/种的厘定以及现代属级概念的建立、DNA分类在线蚓科的应用和线蚓科内的系统发育关系研究亟待开展,以及物种探索的不平衡、经费和研究人才匮乏以及网络分类的缺失等。将分子学数据和系统发育物种概念纳入线蚓科的分类学研究,应该是线蚓科分类的一个方向。通过解读保守基因的信息,可以揭示线蚓科的祖先与它们生活的古环境长期斗争的历史,以及将优良的性状遗传给后代的过程和驱动力。而系统发育物种概念认为物种是拥有共同祖先的,物种仅能通过生殖隔离与系统发育重建一起加以验证。基于系统发育物种概念而构建的线蚓科分类系统,必将能真实的反映线蚓科内各分类单元的亲缘关系和进化轨迹。而将最新的线蚓科分类学知识传播于分类学知识的终端使用者,是线蚓分类学家的职责。这些知识将有助于提高人们对线蚓类在生态系统中功能的了解,如土壤有机质分解、养分矿化和健康评价以及评估气候变化等。
线蚓; 分类; 系统发育; 物种鉴别; 分类学障碍; 展望
线蚓科(Enchytraeidae)隶属于环节动物门环带纲颤蚓目,由Vejdovsky于1879年以线蚓属(Enchytraeus)为模式属而建立,是环带纲的第二大科,目前全世界已知者32属约650余种[1-6], 主要分布于陆地(近500种),淡水和海洋(100余种),河口和冰川等亦有分布[3-4,7]。
生活于陆地的线蚓是土壤生物群落的重要组成部分,而在沼泽地、北方针叶林和苔原带它们的物种多样性尤为丰富。它们主要聚集在土壤表层,以微生物和腐殖质碎屑为食,能加速有机质分解和养分矿化等生态过程被称为“生态系统工程师”[8]。
线蚓科的研究虽然经历了200多年的探索,但有关它的分类学和系统学认识仍然十分肤浅,也是环带动物认知最少的类群之一。本文尝试回顾人类对线蚓科环带动物分类学和系统发育学的认知历程和积累的知识体系,简述线蚓科关键的形态学分类特征以及线蚓科分类研究遇到的主要问题和障碍,然后展望线蚓科未来的研究方向。
1 线蚓科的分类地位
1.1 环节动物门内的系统发育关系
经典的分类学将环节动物门分为多毛纲(Polychaeta)、寡毛纲(Oligochaeta)和蛭纲(Hirudinea)。对于多毛类和寡毛类系统发育关系来说,寡毛类早期被认为起源于类似多毛类的祖先[9]。之后的研究认为,所有的寡毛类和多毛类均起源于类似蚯蚓状(earthworm-like)的共同祖先,该祖先具有多毛类的生殖系统,无独立的生殖腺,无特化的生殖管,只有体腔管,体壁简单,横肌层和纵肌层几乎完整,隔膜完整,循环系统富血管弧,刚毛简单。这个环节动物的祖先向两个方向发展,一类向爬行和洞穴生活进化,成为寡毛类,另一类向游泳生活发展,成为多毛类[10]。也有证据显示,寡毛类和多毛类均以原环虫类(Archiannelida)为共同祖先[11]。
传统观点认为,寡毛纲和蛭纲组成环带总纲(Clitellata),即环节动物中所有具有环带的类群,它们具有雌雄同体,复杂的生殖系统等特征。其中蛭纲包含类蛭(Acanthobdelida, Branchiobdelida)和真蛭亚纲(Euhirudinea)[12-14]。但目前分子研究表明,寡毛纲等同于环带总纲,是蛭纲的祖先。两种之间存在过渡类群,带丝蚓科被认为与蛭类的共同祖先相似,蛭类是寡毛纲后代的一支,但并未列入寡毛纲。20世纪70年代后,分支系统学的发展和分子技术的完善,在大尺度上开展环节动物系统发育的研究十分活跃[15-18]。一系列研究结果均表明环带纲是单系类群。虽然Oligochaeta提出较早,但因前者称呼的广泛接受性以及传统意义上Clitellata包含范围更大,大家仍习惯于称其为Clitellata[19],严格意义上应称其为Oligochaeta。目前已知环带纲动物约有5000余种,占环节动物门(约15000种)的三分之一[20]。
早期的寡毛类环带动物各科系统发育关系研究主要基于先验论。这些研究者先主观设定一个假想的祖先或过度类群,然后根据他们对研究类群知识的掌握程度,应用少量的形态学特征(主要是生殖系统)来进行研究。突出的如Benham提出的大蚓类(Megedrili)和小蚓类(Microdrili)[21]。而Beddard则将大蚓类分为7科,小蚓类为5科[22]。其后近一个世纪的多数研究也是基于这些思路和思想[9-10,23-27]。这些研究的突出缺点是研究结果不能进行验证。
Brinkhurst[11,28-29]认为寡毛类的祖先拥有8对生殖腺,而现存的类群则是在进化过程中这些生殖腺逐渐退化的结果(图1)。他的研究具有合理的逻辑解释和可验证性而得到广泛认可[20]。
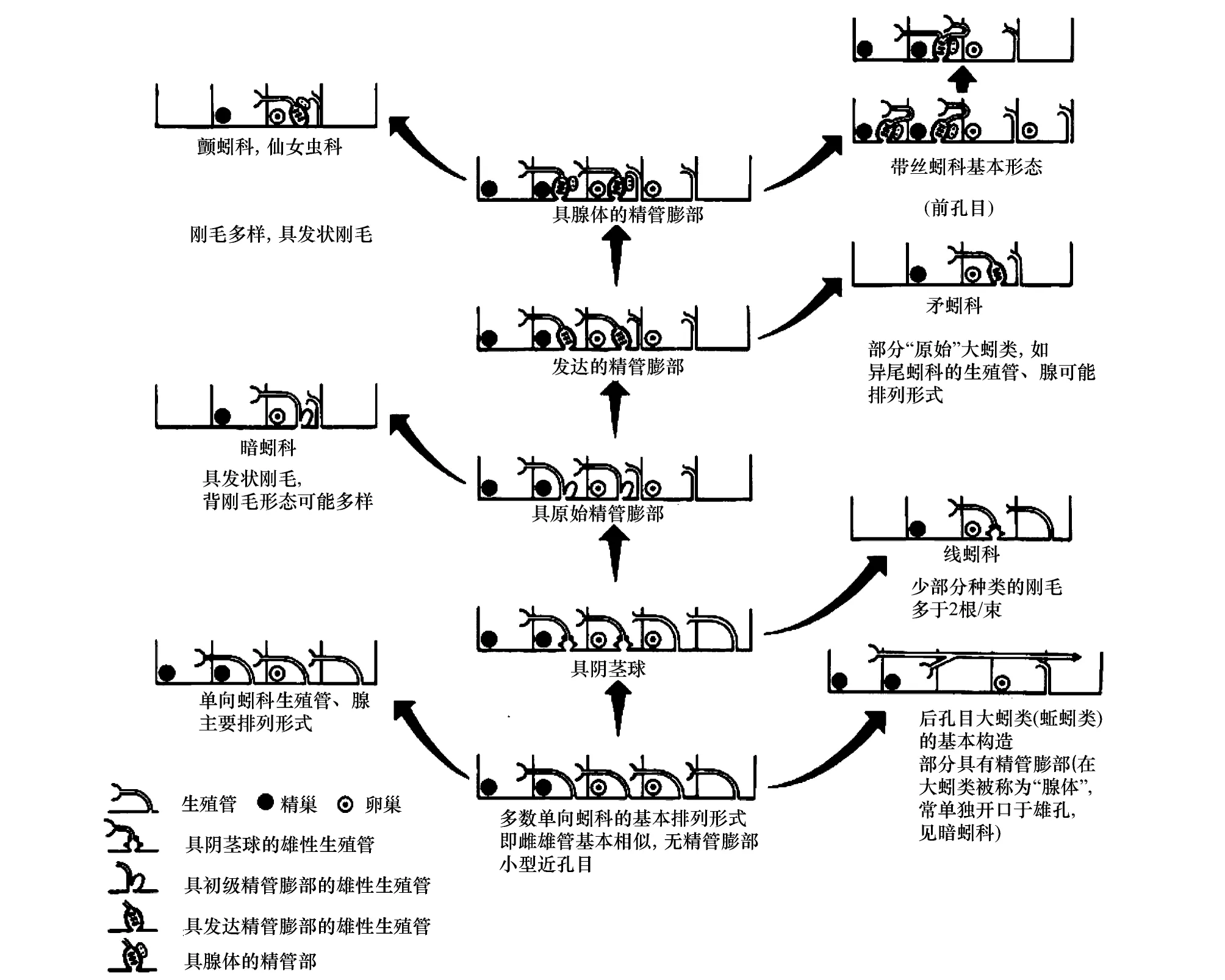
图1 寡毛纲各科的进化路线图,主要基于寡毛类的生殖腺和生殖管的分布格局(改自[28])Fig.1 Evolutionary scheme of oligochaete families, based on patterns of gonads and gonoducts (modified from [28])
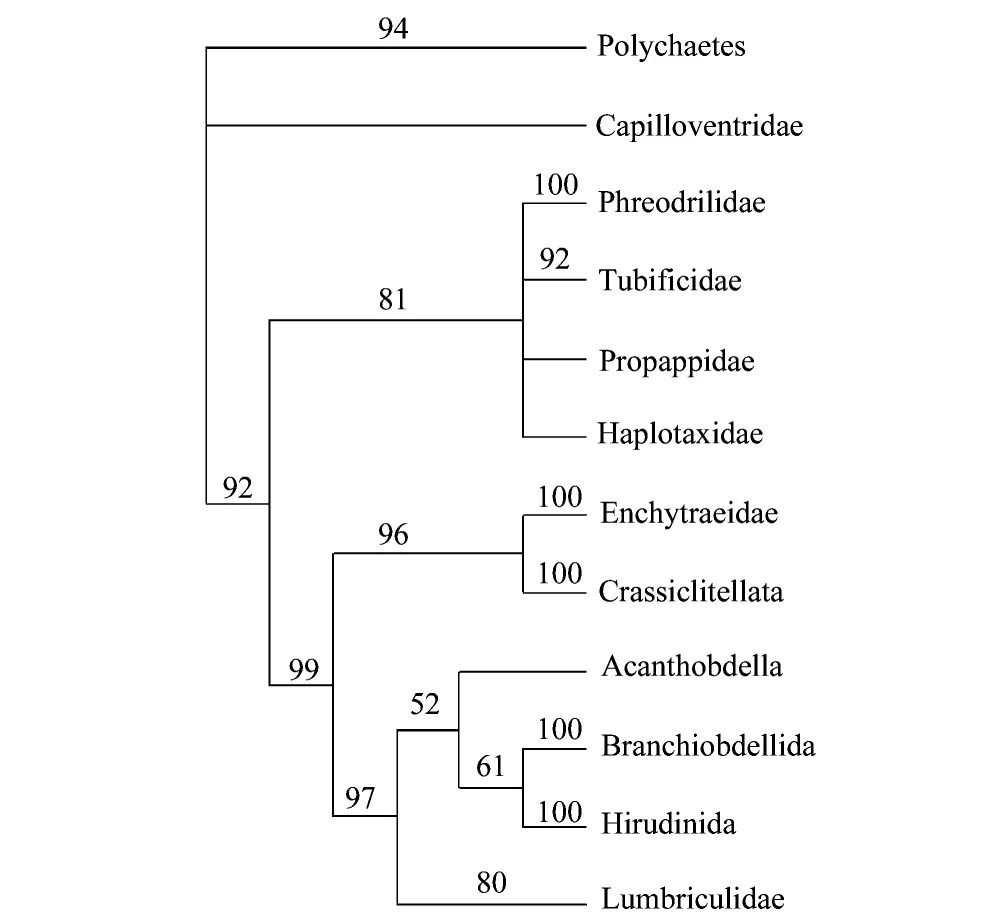
图2 基于18S rDNA 序列的环带纲各类群的系统发育关系[34]Fig.2 Phylogenetic tree of Clitellata based on an analysis of 18S rDNA gene sequences of 39 ingroup clitellates and 12 outgroup polychaetes, using an aligned dataset in which gaps were coded as a fifth character state[34]
1.2 线蚓科的分类地位
线蚓科是环带纲中较大的一类,但其分类地位和科间进化过程至今都缺乏了解。Coates于1987[30-31]和1989年[32],依据形态性状对线蚓进行了系统发育关系研究,将原隶属线蚓科的Propappus属提升为Propappidae科,并将其作为线蚓科的外类群。与此相反,分支系统学研究表明Enchytraeidae和Crassiclitellata互为姐妹群。如果将Propappidae科也包括在内的话,Propappidae和Enchytraeidae似乎不互为姐妹群。2008年,Marotta[33]等人将分子和形态学特征结合分析发现,Crassiclitellata从属于lumbriculid-hirudinean分支,此类群与Enchytraeidae 和 Propappidae组成的类群互为姐妹群 (图2)[34]。
2 线蚓科的分类特征
线蚓科的分类研究属世界性难题。其分类特征极为分散,涉及外部的许多特征,如体色、皮肤腺(epidermal glands)的形状和颜色、环带、刚毛的形状和排列方式、头孔、背孔等,和内部性状如生殖、消化、循环、神经、排泄等组织和器官。而且,许多重要分类性状如食道附属物(oesophageal appendages)、体腔球、排泄肾管等固定后,常易收缩变形而无法观察,分类鉴定必须借助活体观察。故其研究明显滞后于环带动物的其它类群[35]。
线蚓科寡毛类具有一般环节动物的形态特征:头部由口前叶和围口节组成,同律分节,除第一节和最后一节,一般每体节背腹各有2束刚毛;两侧对称、真体腔、雌雄同体等。一般线蚓科性成熟个体体长2—30 mm,体宽0.1—1 mm,体节数很少小于20或大于70[35]。体壁由角质膜、上皮、环肌层、纵肌层和体腔上皮构成。一般环带前皮肤腺较环带后多,性成熟个体XII、XIII节皮肤腺细胞增厚形成环带,XII节刚毛脱落。由脑、围咽神经节、腹神经索构成神经系统。消化道一般由口、咽、食道、肠道和肛门组成。血液循环系统为闭管式。线蚓一般行有性生殖,即雌雄同体,异体受精,以卵茧(cocoon)的方式排出体外。部分种类可进行断裂生殖(fragmentation)[36]。
线蚓科分类的主要依据是:体长与体节数;皮肤腺形态及分布;刚毛形态结构与分布;脑的形态结构;消化道及其附属物;消化肾管和肾管的分布与结构特征;体腔球的形态;生殖腺及生殖器官的形态结构等(图3)[35]。
3 线蚓科分类学研究历史和现状
自1774年Müller发现第1个线蚓Lumbricilluslineatus以来,有关线蚓的研究即引起寡毛类分类学家的广泛关注,相关研究相继开展(图4)。早期的分类学家基于线蚓外部形态性状和对生殖器官等内部形态特征的比较对种和亚种进行直观地归类和整理。由于线蚓其形态等方面有极高的多样性,变异相当复杂,且没有统一的鉴定特征以描述物种,对其进行分类和系统学研究难度较高,故同物异名和异物同名现象非常普遍。
19世纪晚期,欧洲各国研究人员对线蚓科环带动物开展了大量的研究工作。Grube于1851年建立了寡毛纲(Oligochaeta)。1879年,Vejdovsky发表 “Monographie der Enchytraeiden”, 这是关于该科最早的一部专著。1884年,Vejdovsky 发表 “System und Morphologie der Oligochaeten”从解剖学方面对寡毛类作了详尽的描述,是关于寡毛类最早的综述,将寡毛目划分为10科并对其进行了描述,即Aphanoenura,Nadomorpha,Chaetogastridae,Discordrilidae,Enchytraeidae,Tubificidae,Phreoryctidae,Lumbriculidae,Criodrilidae 及Lumbricidae,但对线蚓科并未做详细介绍。Beddard 于1900年发表 “Monograph of the order Oligochaeta”。
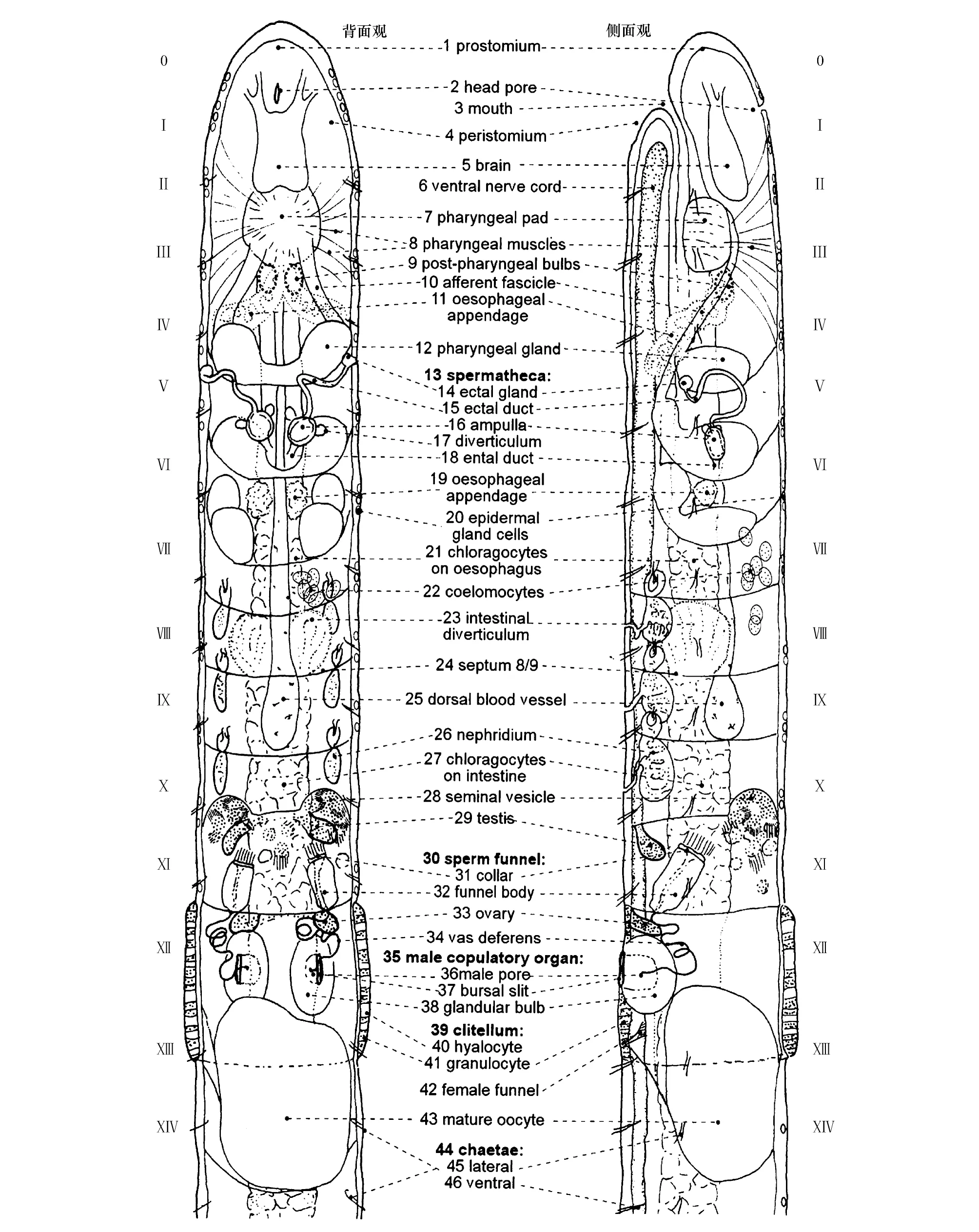
图3 线蚓前14节背面观和侧面观,示主要的形态学分类特征[35]Fig.3 Dorsal and lateral views of enchytraeid within anterior 14 segments showing the key morphological traits of identification[35] 1—46分类特征分别为口前叶、头孔、口、围口节、脑节、腹神经束、咽板、咽肌肉束、咽后球、咽后肌肉束、咽后附属物、食道腺、受精囊、受精囊外管腺体、受精囊外管、受精囊体、受精囊体分支、受精囊内管、食道附属物、皮肤腺、食道外壁黄色细胞、体腔球、肠道分支、隔膜、背血管、肾管、肠道外壁黄色细胞、储精囊、精巢、精漏斗、漏斗领、漏斗体、卵巢、输精管、雄性交配器官、雄孔、雄孔外裂、交配球、环带、透明细胞、颗粒细胞、雌性漏斗、成熟卵、刚毛、侧刚毛和腹刚毛
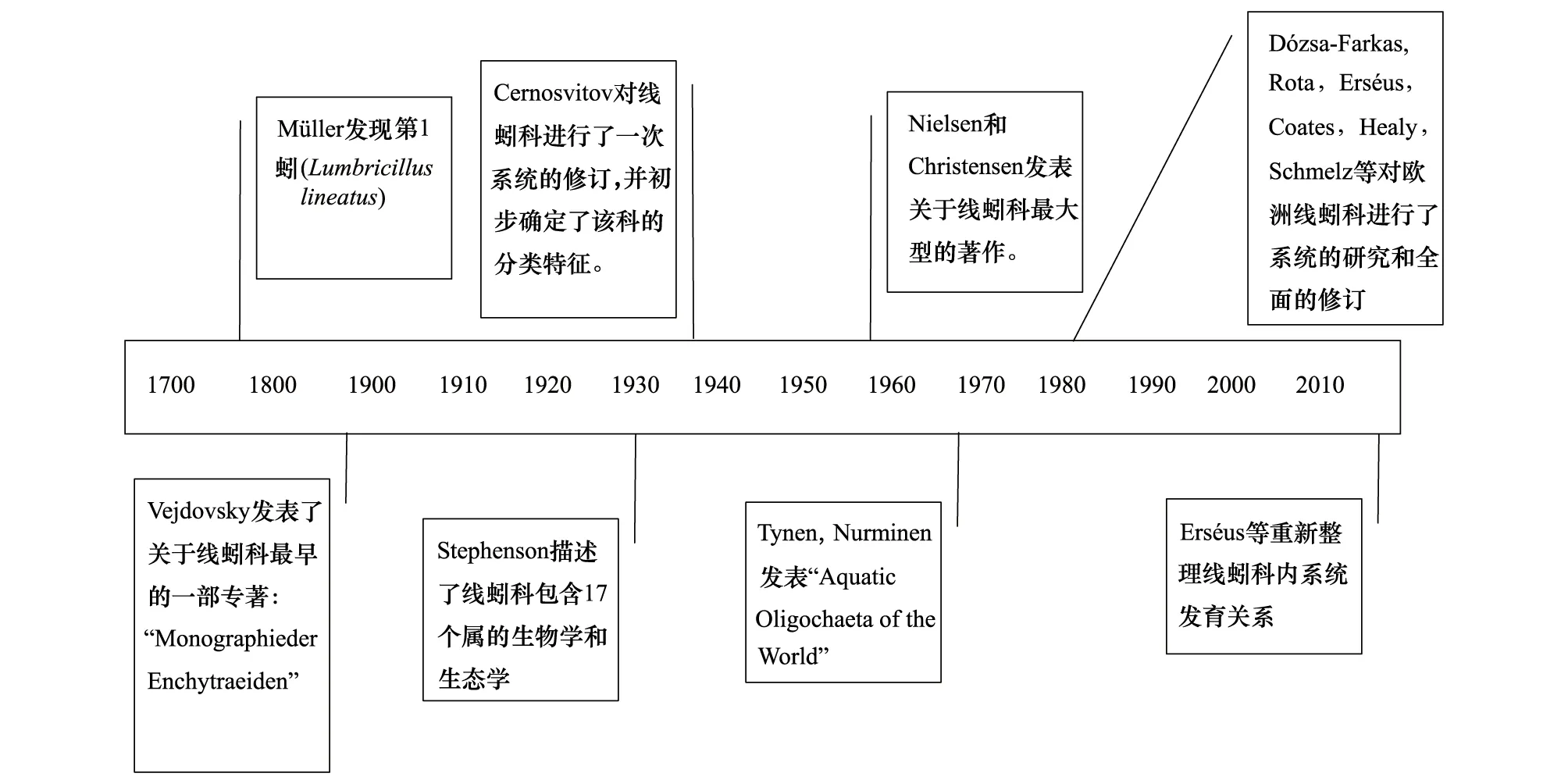
图4 线蚓科分类学研究大事记Fig.4 The chronicle of events of enchytraeid classification and taxonomy
19世纪末,20世纪初,大量物种被发现。英国学者Stephenson于1930年出版了寡毛类分类著作《The Oligochaeta》,详细描写了寡毛类的生物学和生态学等特征,其中线蚓科包含17个属,但其系统位置没有解决。Cernosvitov于1937年发表 “Systema der Enchytraeiden” 一文,该文对线蚓科进行了一次系统的修订,除了对线蚓科系统发育进行讨论外,还描述了他认为的可以接受的303个种和69个可疑种,并初步确定了该科动物的分类特征。
随后,整个20世纪,对线蚓科的记载不仅包括了淡水、海洋、陆地等不同生态环境,而且涉及热带、温带、寒带(极地)等各种大陆气候区。其中Stephenson、Cernosvitov、Nielsen、Christensen、Coats、Dózsa-Farkas等对欧洲各国以及南、北美洲、澳大利亚等地线蚓的研究工作做出了突出贡献[3]。
1959—1963年,Nielsen和Christensen系统整理并描述了欧洲各国线蚓,发表“the Enchytraeidae, critical revision and taxonomy of European species”系列专著[1,37-38],这也是到目前为止关于线蚓科最大型的著作。该文是在Cernosvitov的基础上,较系统的对欧洲种类进行重新整理,并尝试用染色体数目作为种类鉴定特征。虽然该文没有完整的对非欧洲种类进行完整的描述,而且并没有完全遵照国际动物命名法规,但他们对该科的划分依然被后人沿用,奠定了线蚓科现代分类学基础。
随后Tynen和Nurminen等对欧洲和北美的线蚓进行了系统的整理,撰写和发表一系列文章:1969年合作完成了“A key to the European littoral Enchytraeidae (Oligochaeta) ”一文;1975 年Tynen发表 “A Checklist and Bibliography of the North American Enchytraeidae (Annelida: Oligochaeta)”。“Aquatic Oligochaeta of the World”[39]一书提及线蚓科21属。上述的工作主要集中在北半球[3]。
60年代末Bell,Righi以及他的学生Christofensen等开始对南美洲特别是巴西等地线蚓进行调查,经过30多年的研究,迄今已发现线蚓科24个新种[40]。关于南非线蚓的研究也有零星报道。
随着描述的物种增多,自20世纪中叶开始,人们尝试着寻找可靠的分类特征。1959年,Nielsen 和 Christensen在“the Enchytraeidae, critical revision and taxonomy of European species” 一文中尝试利用染色体数目对线蚓进行分类。1996年,Schmelz利用形态学特征与蛋白质条带相结合的方法对线蚓进行物种鉴定[41];1998年,Schirmacher用RAPD-PCR方法鉴定两个相似陆栖线蚓[42]。
4 线蚓科科内系统发育研究现状
线蚓科内水平系统发育关系研究发展缓慢。Eisen最初把线蚓科分为4个亚科16属:中线蚓亚科Mesenchytraeinae (中线蚓属),线蚓亚科Enchytraeinae (线蚓属、Michaelsena属)、裸线蚓亚科Achaetinae (裸线蚓属)和丝线蚓亚科Lumbricillinae (包含其余所有12属)。Cejka把线蚓科分为白线蚓亚科Fridericiinae,中线蚓亚科Mesenchytraeinae 和亨利蚓亚科Henleinae[43]。Michaelsen 认为线蚓科寡毛类与仙女虫科和螵体虫科具有直接的亲缘关系[44-45],Stephenson持不同意见。Stephenson提出了寡毛类各类群间亲缘关系,而线蚓科可能起源于Phreodrilidae,即现今Haplotaxidae。线蚓科各类群则根据刚毛形状、数目、背血管起始等形态性状分为几个类群:1)以Henlea为中心的Buchholzia,Bryodrilus,Aspiododrilus,Fridericia和Distichopus;2)以Enchytraeus为中心的Marionina,Lumbricillus和Michaelsena;3)Achaeta,Mesenchytraeus,Stercutus,Litorea和Chirodrilus各属独立。其中Henlea属依据其是否有消化道盲囊分为HenleaFriend、HenleanellaFriend 和HepatogasterCejka 3个亚属[9]。Cernosvitov认为线蚓科应分为6个亚科[46]。Chekanovskaya认为线蚓科和仙女虫科具有共同的祖先[47]。
到20世纪70年代后期,随着系统学研究方法的进展和对线蚓科寡毛类研究的重视和认识的深入,分类学家尝试在系统发育研究的基础上来构建线蚓科级分类阶元和属级分类阶元的系统关系。Coates依据形态性状对线蚓进行了系统发育关系研究,将原隶线蚓科的Propappus属提升为Propappidae科,并将其作为线蚓科的外类群,对线蚓科11属103种的129个形态学性状在属级水平上对其亲缘关系进行分析[31-32]。研究结果显示,裸线蚓亚科(Achaetinae)为非单系群,仅裸线蚓(Achaeta)、白线蚓(Fridericia)、Randidrilus和Enchytronia为单系群,其余7属为复系类群,而玛利安蚓(Marionina)最为紊乱,需进行重新修订[48-49]。并推测线蚓科的现存种应起源于南美洲、非洲和印度。最新的研究支持半线蚓(Hemienchytraeus)和裸线蚓的单系性,而多数属为复系类群[50]。典型的如苔丝蚓(Bryodrilus)、欧氏蚓(Oconnorella)和亨利蚓(Henlea)互为姊妹群,柯氏蚓(Cognettia)、斯特科特蚓(Stercutus)、楔氏蚓(Cernosvitoviella)和中线蚓(Mesenchytraeus),以及拥有两种体腔球的半白线蚓(Hemifridericia)、巴氏蚓(Buchholzia)和白线蚓各组成一个单系群。丝线蚓(Lumbricillus)和玛利安蚓最为紊乱,需进行重新厘定[4,51]。De Wit等[52]利用分子数据系统开展海洋针线蚓的系统发育与其关键形态学特征的进化轨迹,显示该属的许多形态性状高度保守,而空间距离是影响其物种亲缘关系的关键驱动要素。总之,线蚓科类群复杂,种类丰富,地理分布较广泛。线蚓属/种间分类性状分散、系统发育研究匮乏,因此线蚓科的系统发育关系研究亟待开展。
5 我国线蚓科分类学研究现状
受Stephenson提出的即认为线蚓在东洋区分布甚少这一观点的影响, 我国线蚓科的研究重视不够,实际开展线蚓科的研究则始于20世纪90年代。早期研究力量十分薄弱,主要是一些零星的新种描述。1907年,Stewart从我国西藏采得标本,经Stephenson鉴定为腹式亨利蚓(H.ventriculosa)[53]。这是关于我国线蚓最早的记述。陈义记载了2种。梁彦龄等[54]报道西藏冰川华线蚓 (SinenchytraeusglacialisLiang and Hsü, 1979) 1新属新种,许智芳等[55]报道岷县等毛蚓(IsosetosaminxianensisXu et al, 1989) 1新属新种, 受到国际同行的关注。近20年来,我国对线蚓科的分类学研究有了长足的进步。1992年梁彦龄和谢志才记载了浙江天目山线蚓科4属8种。1999年以后,谢志才、王洪铸、陈静、沈琦等报道长江流域、珠江流域、吉林长白山和陕西省华山等地共计12属60种,包括新种31种,新记录属8属[56-73]。海洋方面,Erséus和梁彦龄等[74-79]先后开展胶州湾和香港的海洋线蚓科种类,共计5属11种。目前我国经记载的线蚓科种类达17属94种[55,70-73],约占世界经记录线蚓种类的1/8,改变了中国线蚓科寡毛类种类贫乏的传统观点。
6 线蚓科分类学研究存在的问题
6.1 未描述种类亟待发掘
线蚓科的物种多样性极为丰富,特有物种众多,大量的新物种亟待发掘。但世界各大洲线蚓的物种探索极不平衡。欧洲是线蚓科物种发掘最为深入的地区,至今已记录500余种,而南美洲77种,北美洲37种,非洲34种,大洋洲15种,亚洲约100种。其次,线蚓科物种探索集中与土壤和海洋系统,而在淡水、冰川和河口等的研究较少。以中国线蚓科的分类为例,虽然我国是线蚓科物种记录最多的亚洲国家(17属94种),但13属70余种为土壤分布的种类,而吉林省的长白山地区研究的最为深入(10属51种),包括新种16种,而我国海洋的线蚓科种类仅有零星报道,淡水、河口和冰川的记录几乎为空白。
6.2 已描述的物种需要验证
首先,早期记录物种的原始描述过于简单,使用的形态分类特征过于单一,缺乏对种内/种间形态变异的整体理解,以及对个体发育变异、异速生长和不同居群之间的地理差异的整体把握。其次,现代线蚓科的分类要求标本必须在活体状态下观测,许多关键分类特征的信息(如体色、皮肤腺、食道附属物、体腔球等)只有在活体下才能准确获取。再次,研究者在进行分类整理的过程中,多未检视模式标本,甚至没有与地模标本进行比对,导致广布种的普遍出现,所列出的同物异名表需要重新确认。因此,线蚓科的物种分类有必要从新的视角进行审视。
6.3 属/种的厘定及其现代属级概念的建立
鉴于线蚓科的主要类群多为复系群,线蚓学家们均在开展各属的修订和整理工作[51,80]。突出的如,Schmelz[81]全面整理了白线蚓属,提出了副神经腺体等一系列该属新的形态分类性状,创建了白线蚓属的全新的分类体系。在经描述的197种中,仅89种被认为是有效种,这是线蚓科单属系统整理最为细致的工作。Xie 和 Rota[67]发现头孔的位置、刚毛的形状和排列方式、背血管前段的起始位置、隔膜腺的形状、脑节的形状等是鉴定玛利安蚓属物种的重要特征。陈静等[64]对欧氏蚓属进行重新修订,提出了食道附属物的有无、背血管的起始位置和消化肾管的组成等5个关键属征。最近,我们在研究中线蚓属时,发现精荚(spermatozeugmata)的有无、形状和排列方式可能是鉴定该属的重要的性状[70-71]。各属间的属征明晰、属内物种间亲缘关系的研究,新的分类特征的发掘、评估和应用,以及建立现代属级的统一的分类系统,是线蚓科研究迫切需要解决的问题。
6.4 DNA分类在线蚓科的应用
自Tautz等[82-83]提出DNA分类(DNA taxonomy)的概念以来,就迅速被分类学家所接受,并广泛应用于各类生物的分类学研究[84-86]。把DNA序列作为生物分类系统的主要平台,即利用保守的分子序列,来定义和判断与传统意义上的物种有相似范围的分子操作分类单元(molecular operational taxonomic unit, MOTU)[87]。由于细胞色素c氧化酶I基因(COI) 的演化速率较慢,在分类上具有更大的物种识别能力,一般将其作为目的基因来进行DNA分类[88]。虽然有些线蚓学家尝试应用DNA序列来鉴别一些外形相似的物种和隐存种(cryptic species)[89-91],但DNA分类在线蚓科实际尚未大规模展开。相信随着DNA分类的广泛应用,并结合经典的形态学分类,能极大地促进线蚓科的分类学研究。
6.5 线蚓科内的系统发育关系研究亟待开展
基于形态学和分子学数据的研究证实线蚓科为单系类群,但线蚓科内多数属为复系类群[4,50]。现有的线蚓科分类体系并不能很好的反映该科各属的亲缘关系。而人们对线蚓科的物种分化样式、物种进化历史等的认识几乎为空白。形态学分类特征的高度多样化表明线蚓科在进化过程中可能经历多次的性状的趋同/趋异进化(如刚毛的获得与丢失)。而基于分子学数据的线蚓科内系统发育进化树的构建应是复原这些过程的最好的途径。
7 总结与展望
虽然经历了200多年的研究,人们积累的对线蚓科的分类学知识仍然极其有限,建立的线蚓科分类系统仍然不能很好的反映线蚓科的自然进化历史。而基于生物学物种概念(生殖隔离)的线蚓科物种探索,虽然有一些合理的逻辑解释,但缺乏严格的科学验证。从科学的可验证性上来说,现今已建立的线蚓科分类体系还只是对线蚓科认识的经验积累和初级阶段(α分类阶段)。而线蚓科分类学特征的多样性和生殖隔离的不可验证性表明,仅根据形态特征对线蚓科分类很难揭示线蚓科的自然进化关系。而将分子学数据和系统发育物种概念纳入线蚓科的分类学研究,应该是线蚓科分类的一个方向。通过利用保守的基因片段(如COI)的信息,可以解读线蚓科的祖先与它们生活的古环境长期斗争的历史,以及将这些优良的性状遗传给后代的过程和驱动力。而系统发育物种概念认为物种是拥有共同祖先的,是在系统发育类别中最小的种群聚集体。从实际操作上来讲,相比生物学物种概念而言,系统发育物种概念相对更严谨,它认为物种仅能通过生殖隔离与系统发育重建一起加以验证。
制约线蚓科分类的关键要素包括:(1)物种探索的不平衡(见6.1)。(2)研究经费的缺乏和网络分类(e taxonomy)。传统的分类研究很难争取到研究经费。而相关的线蚓科分类文献资料零散,应开展线蚓科的网络分类研究工作,让研究者能共享这些研究信息。(3) 分类学障碍(taxonomic impediment)。目前专职研究线蚓科分类的学者全世界不超过10人,这些人主要分布在欧洲。线蚓科分类学人才极度匮乏。
线蚓科由于其对栖息地环境的适应性辐射进化,是生态学和进化研究的理想类群。线蚓科广泛分布于陆地生态系统中,是土壤生态系统的主要类群,线蚓主要以微生物和腐殖质碎屑为食,通过取食、消化、排泄(粪便)、分泌(粘液)和掘穴等活动改善土质,加速土壤的物质循环和能量传递[92-93]。它们对人类干扰(尤其是土壤杀虫剂、各种重金属等)的反应极为敏感,是评估土壤生态系统健康与否的理想指示类群,也是评价全球气候变化(如气温的升高、CO2浓度的波动和降雨量的减少引起的土壤干旱等)的理想生物[94]。另外,线蚓科不仅物种丰富,而且特有种分化显著,是生物区系分化问题探讨以及生物地理学研究的理想材料。但是目前尚无有关世界线蚓科的分类学专著。仅有的线蚓科动物志仅限于欧洲。而这些专著不能满足相关学者和大众的需求。而编撰世界的线蚓科物种名录、地理分布和检索表,将最新的线蚓科分类学知识传播于分类学知识的终端使用者,是线蚓分类学家的职责。
[1〗 Nielsen C O, Christensen B. The Enchytraeidae Critical revision and taxonomy of European species. Natura Jutlandica, 1959, 8-9: 1-160.
[2] Nakamura Y. Checklist of enchytraeids (Oligochaeta: Enchytraeidae) of the world. Miscellaneous Publication of the Tohoku National Agricultural Experiment station, 2000, 24: 29-104.
[3] Martin P, Martinez-Ansemil E, Pinder A, Timm T, Wetzel M J. Global diversity of oligochaetous clitellates (“Oligochaeta”: Clitellata) in freshwater. Hydrobiologia, 2008, 595(1): 117-127.
[4] Erséus C, Rota E, Matamoros L, De Wit P. Molecular phylogeny of Enchytraeidae (Annelida, Clitellata). Molecular Phylogenetics and Evolution, 2010, 57(2): 849-858.
[5] Schmelz R M, Collado R. An updated checklist of currently accepted species of Enchytraeidae (Oligochaeta, Annelida). Landbauforschung -vTI Agriculture Forestry and Research, 2012, 357: 67-87.
[6] Christensen B, Dózsa-Farkas K. A new genusGlobulidrilusand three new enchytraeid species (Oligochaeta: Enchytraeidae) from Seoraksan National Park (Korea). Journal of Natural History, 2012, 46(45/46): 2769-2785.
[7] Hartzell P, Nghiem J V, Richio K J, Shain D H. Distribution and phylogeny of glacier ice worms (MesenchytraeussolifugusandMesenchytraeussolifugusrainierensis). Canadian Journal of Zoology, 2005, 83(9): 1206-1213.
[8] Lavelle P, Bignell D, Lepage M, Wolters V, Roger P, Ineson P, Heal OW, Dhillion S. Soil function in a changing world: the role of invertebrate ecosystem engineers. European Journal Soil Biology, 1997, 33: 159-193.
[9] Stephenson J. The Oligochaeta. Oxford: Clarendon Press, 1930: 1-978.
[10] Brinkhurst R O, Jamieson B G M. Aquatic Oligochaeta of the World. Edinburgh: Oliver & Boyd, 1971: 1-860.
[11] Brinkhurst, R O. Evolution in the Annelida. Canadian Journal of Zoology, 1982, 60(5): 1043-1059.
[12] Sawyer R T. Leech Biology and Behavior. Oxford: Clarendon Press, 1986.
[13] Brusca R C, Brusca G J. Invertebrates. Sinauer Associates Sunderland, 2002, i-xviii: 1-936.
[14] Rouse G W, Fauchald K. The articulation of annelids. Zoologica Scripta, 1995, 24(4): 269-301.
[15] McHugh D. Molecular systematics of polychaetes (Annelida). Hydrobiologia, 2005, 535-536(1): 309-318.
[16] Rousset F, Roze D. Constraints on the origin and maintenance of genetic kin recognition. Evolution, 2007, 61(10): 2320-2330.
[17] Purschke G, Hessling R, Westheide W. The phylogenetic position of the Clitellata and the Echiura-on the problematic assessment of absent characters. Journal of Zoological Systematics and Evolutionary Research, 2000, 38(3): 165-173.
[18] Borda E, Siddall M. Review of the evolution of life history strategies and phylogeny of the Hirudinida (Annelida: Oligochaeta). Lauterbornia, 2004, 52: 5-25.
[19] Martin P. On the origin of the Hirudinea and the demise of the Oligochaeta. Proceedings of the Royal Society of London Series B-Biological Sciences, 268, 1471: 1089-1098.
[20] Erséus C. Phylogeny of oligochaetous Clitellata. Hydrobiologia, 2005, 179: 357-372.
[21] Benham W B. An attempt to classify earthworms. Quarterly Journal of Microscopical Science (New Series), 1890, 31: 201-315.
[22] Beddard F E. A Monograph of the Order of Oligochaeta. Oxford: Clarendon Press, 1895: 769-769.
[23] Michaelsen W. Zur Stammgeschichte der Oligochaeten. Zeitschrift für wissenschaftliche Zoologie, 1929, 134: 694-716.
[24] Timm T. On the origin and evolution of aquatic Oligochaeta. Eesti NSV Teaduste Akadeemia Toimetised Bioloogia, 1981, 30: 174-181.
[25] Omodeo P. History of Clitellata. Italian Journal of Zoology, 1998, 65(1): 51-73.
[26] Omodeo P. Evolution and biogeography of megadriles (Annelida, Clitellata). Italian Journal of Zoology, 2000, 67(2):179-201.
[27] Erséus C. Aspects of the phylogeny of the marine Tubificidae. Hydrobiologia, 1984, 115(1): 37-44.
[28] Brinkhurst R O. Comments on the evolution of the Annelida. Hydrobiologia, 1984, 109(2): 189-191.
[29] Brinkhurst R O. The position of the Haplotaxidae in the evolution of oligochaete annelids. Hydrobiologia, 1984, 115(1): 25-36.
[30] Coates K A. Phylogenetic analysis of sorne Enchytraeidae (Annelida:Oligochaeta) parsimony analysis of structural characters[D]. Victoria: University of Victoria, 1987.
[31] Coates K A. Phylogenetics of some Enchytraeidae (Annelida: Oligochaeta): a preliminary investigation of relationships to the Haplotaxidae. Hydrobiology, 1987, 40: 91-106.
[32] Coates K A. Phylogeny and origins of Enchytraeidae. Hydrobiology, 1989, 180(1): 17-33.
[33] Marotta R, Ferraguti M, Erséus C, Gustavsson L M. Combined-data phylogenetics and character evolution of Clitellata (Annelida) using 18S rDNA and morphology. Zoological Journal of the Linnean Society, 2008, 154(1): 1-26.
[34] Erséus C, Källersjö M. 18S rDNA phylogeny of Clitellata (Annelida). Zoologica Scripta, 2004, 33: 187-196.
[35] Schmelz R M, Collado R. A guide to European terrestrial and freshwater species of Enchytraeidae (Oligochaeta). Soil Organisms, 2010, 82(1): 1-176.
[36] Nakamura Y. A new fragmenting enchytraeid species, Enchytraeus japonensis from a cropped Kuroboku soil in Fukushima, Northern Japan (enchytraeids in Japan 5), Edaphologia, 1993, 50: 37-39.
[37] Nielsen C O, Christensen B. The Enchytraeidae. Critical revision and taxonomy of European species. Natura Jutlandica, 1961, 10 (S1): 1-23.
[38] Nielsen C O, Christensen B. The Enchytraeidae. Critical revision and taxonomy of European species. Natura Jutlandica, 1963, 10 (S2): 1-19.
[39] Brinkhurst R O, Jamieson B G M. Aquatic Oligochaeta of the World. Toronto: University of Toronto Press, 1971: 1-860.
[40] Römbke J. The role of Gilberto Righi in the development of tropical microdrile taxonomy. The 7th international symposium on earthworm. ecology Cardiff Wales 2002. Pedobiologia, 2003, 47(5/6): 405-412.
[41] Schmelz R M. Additional morphological traits facilitate enchytraeid determination. Newsletter on Enchytraeidae, 1996, 5: 45-46
[42] Schirmacher A, Schmidt H, Westheide W. RAPD-PCR investigations on sibling species of terrestrailEnchytraeus(Annelida: Oligochaeta). Biochemical Systematics and Ecology, 1998, 26(1): 35-44.
[43] Cejka B. Die Oligochaeten der Russischen in den Jahren 1900-1903 unternommenen Nordpolarexpedition. I. Über eine neue Gattung der Enchytraeiden (Hepatogaster). Mémoires de l′Academie Impériale des Sciences de St.-Pétersbourg 8e série, Classe Physico-Mathematrque, 1910, 29(6): 1-29.
[44] Michaelsen W. Zur Stammgeschichte und Systematik der Oligochäten, insbesondere der Lumbriculiden. Archiv für Naturgeschichte, Berlin, 1921, 86(8): 130-141.
[45] Michaelsen W. Schmarotzende Oligochäten nebst Erörterungen über verwandtschaftliche Beziehungen der Archioligochäten. Mitteilungen aus dem Staatsinstitut und Zoologischen Museum in Hamburg, 1926, 42: 91-103.
[46] Cernosvitov L. System der Enchytraeiden. Bulletin de l′Association Russe pour les recherches scientifiques à Prague, 1937, 5: 263-295.
[47] Chekanovskaya O V. Aquatic Oligochaeta of the USSR. Keys to the fauna of the USSR. Moscow: Academy of Sciences of the USSR Publishers, 1981: 1-513.
[48] Rota E, Matamoros L, Erséus C. In search ofMarionina(Clitellata, Enchytraeidae): a taxonomic history of the genus and re-description of the type speciesPachydrilusgeorgianusMichaelsen, 1888. Italian Journal of Zoology, 2008, 75(4): 417-436.
[49] Schmelz R M, Collado R. A type-based redescription ofPachydrilusgeorgianusMichaelsen, 1888, the type species ofMarioninaMichaelsen, 1890, with comments onChristensenidrilusDózsa-Farkas & Convey, 1998 (Enchytraeidae, “Oligochaeta”, Annelida). Verhandlungen des Naturwissenschaftlichen Vereins in Hamburg, 2008, 44: 7-22.
[50] Christensen B, Glenner H. Molecular phylogeny of Enchytraeidae (Oligochaeta) indicates separate invasions of the terrestrial environment. Journal of Zoological Systematics and Evolutionary Research, 2010, 48: 208-212.
[51] Matamoros L. Systematics ofMarionina(Annelida: Clitellata: Enchytraeidae) [D]. Göteborg, Sweden: University of Gothenburg, 2011.
[52] De Wit P, Emilia R, Erséus C. Phylogeny and character evolution inGrania(Annelida, Clitellata). Zoologica Scripta, 2011, 40(1): 509-519.
[53] Stephenson J. Report on a collection of aquatic animals made in Tibet by Captain F. H. Stewart, II. Oligochaete worms, Mollusca and fish. Records of the Indian Museum, 1909, 3: 105-144.
[54] 梁彦龄,许智芳,张德宁. 西藏冰川上线蚓科一新属新种. 动物分类学报, 1979, 4 (4): 312-315.
[55] 许智芳,张德宁,姜建明. 甘肃陆栖寡毛类一新属新种(寡毛纲:近孔目:线蚓科). 动物分类学报, 1989, 14: 153-156.
[56] 陈静. 中国陆栖线蚓科环带动物分类学与生态学研究. 中国科学院水生生物研究所, 2009.
[57] Wang H Z, Xie Z C, Liang Y L. Records of Enchytraeidae (Clitellata) from the People′s Republic of China. Hydrobiologia, 1999, 406: 57-66.
[58] Xie Z C, Wang H Z, Liang Y L. Studies on the Enchytraeidae (Oligochaeta, Annelida) of China. I. On new species and new records of the genusHemienchytraeus. Acta Hydrobiologica Sinica, 1999, 23(4): 352-358.
[59] Xie Z C, Liang Y L, Wang H Z. Taxonomic study onFridericia(Enchytraeidae, Oligochaeta) along the Changjiang (Yangtze) Basin. Acta Hydrobiologica Sinica, 1999, 23(s1): 158-163.
[60] Xie Z C, Liang Y L, Wang H Z. Two new species ofFridericia(Enchytraeidae, Oligochaeta) from Changbaishan Mountain, Jilin Province, China. Species Diversity 2000, 5(1): 53-58.
[61] Xie Z C, Liang Y L, Wang H Z.Enchytraeuschaoyangensis, a new terrestrial culture species (Enchytraiedae, Oligochaeta) from north-east of China.ActaHydrobiologica, 2000, 42(1/2): 69-72.
[62] Xie Z C, Liang Y L, Wang H Z. A taxonomic study ofBryodrilus(Enchytraeidae, Oligochaeta) from Changbaishan Mountain, China. Species Diversity, 2000, 5(2): 93-101.
[63] Chen J, Xie Z C.Bryodrilusfuscistriatus, a new enchytraeid species (Oligochaeta: Enchytraeidae) from northeastern China. Proceedings of the Biological Society of Washington, 2006, 119(2): 195-201.
[64] Chen J, Xie Z C, He S P. A Taxonomic Study ofOconnorella(Enchytraeidae, Oligochaeta) from Changbaishan Mountain, China. Zoological Science, 2006, 23(10): 917-922.
[65] Chen J, Xie Z C.Fridericiadianchiensissp. nov. (Enchytraeidae, Oligochaeta) from Yunnan Province, China. Proceedings of the Biological Society of Washington, 2008, 121(2): 283-288.
[66] Chen J, Xie Z C.Fridericialiangisp. nov. (Enchytraeidae, Oligochaeta) from Changbaishan Mountain, China. Proceedings of the Biological Society of Washington, 2009, 122(4): 399-404.
[67] Xie Z C, Rota E. Four new terrestrial species ofMarionina(Clitellata, Enchytraeidae) from China and re-examination ofM.hoffbaueriMöller. Journal of Natural History, 2001, 35(10): 1417-1431.
[68] Xie Z C, Liang Y L, Wang J.Fridericiananningensis, a new terrestrial enchytraeid species (Oligochaeta) from south-western China. Proceedings of the Biological Society of Washington, 2001, 114(1): 275-279.
[69] Xie Z C.Mesenchytraeusgigachaetus, new name, a replacement name forMesenchytraeusmegachaetusShen et al., 2011. Proceedings of the Biological Society of Washington, 2012, 125(1): 18-18.
[70] Shen Q, Chen J, Xie Z C.Mesenchytraeusmegachaetus, a new enchytraeid with enlargened ventral chaetae (Annelida, Clitellata) from ChangbaishanMountain, China. Proceedings of the Biological Society of Washington, 2011, 124(2): 127-133.
[71] Shen Q, Chen J, Xie Z C.Mesenchytraeusmonodiverticulussp. nov (Annelida: Clitellata: Enchytraeidae) from Changbai Mountain, with a key to Mesenchytraeus with enlarged chaetae. Proceedings of the Biological Society of Washington, 2012, 125(3): 215-227.
[72] Shen Q, Chen J, Xie Z C.Mesenchytraeusanisodiverticulus, a new enchytraeid with enlarged ventral chaetae (Annelida, Clitellata) from north-eastern China. Italian Journal of Zoology, 2012, 79(1): 86-91.
[73] 沈琦. 长白山线蚓科(环节动物门, 环带纲)分类学研究. 中国科学院水生生物研究所, 2012.
[74] Erséus C. The marine Tubificidae (Oligochaeta) of Hong Kong and southern China. Asian Marine Biology, 1984, 1: 135-175.
[75] Erséus C. Marine Oligochaeta of Hong Kong// Morton B. ed., The Marine Flora and Fauna of Hong Kong and Southern China II. Hong Kong: Hong Kong University Press, 1990: 259-335.
[76] Erséus C. Marine Oligochaeta of Hong Kong: A supplement//Morton B. ed.. The Marine Flora and Fauna of Hong Kong and Southern China III. Hong Kong: University Press Hong Kong, 1992: 157-180.
[77] Erséus C. Oligochaeta from Hoi Ha Wan//Morton B. ed. The Marine Flora and Fauna of Hong Kong and Southern China III. Hong Kong: University Press Hong Kong, 1992: 909-917.
[78] Erséus C. Additional notes on the taxonomy of the marine Oligochaeta of Hong Kong, with a description of a new species of Tubificidae//Morton B. ed. The Marine Flora and Fauna of Hong Kong and Southern China IV, Hong Kong: University Press Hong Kong, 1997: 37-52.
[79] Erséus C, Sun D, Liang Y L, Sun B. Marine Oligochaeta of Jiaozhou Bay, Yellow Sea coast of China. Hydrobiologia, 1990, 202(1/2): 107-124.
[80] Schmelz R M, Collado R, Römbke J. Mata Atlntica enchytraeids (Paraná, Brazil): A new genus,Xetadrilusgen. nov., with three new species, and four new species ofGuaranidrilusernosvitov (Enchytraeidae, Oligochaeta). Zootaxa, 2011, 2838: 1-29.
[81] Schmelz R M. Taxonomy ofFridericia(Oligochaeta, Enchytraeidae): Revision of species with morphological and biochemical methods. Keltern-Weiler. Goecke & Evers, 2003: 1-415.
[82] Tautz D, Arctander P, Minelli A, Thomas R H, Vogler A. DNA points the way ahead in taxonomy. Nature, 2002, 418(6897): 479-479.
[83] Tautz D, Arctander P, Minelli A, Thomas R H, Vogler A P. A plea for DNA taxonomy. Trends in Ecology and Evolution, 2003, 18(2): 70-74.
[84] Hebert P D N, Penton E H, Bums J M, Janzen D H, and Hallwachs W. Ten species in one: DNA barcoding reveals cryptic species in the neotropital skipper butterflyAstraptesfulgerator. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(41): 14812-14817.
[85] Pons J, Barraclough TG, Gomez-Zurita J, Cardoso A, Duran D P, Hazell S, Kamoun S, Sumlin W D, Vogler A P. Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Systematic Biology, 2006, 55(4): 595-609.
[86] Vuataz L, Sartori1 M, Wagner A, Monaghan M T. Toward a DNA taxonomy ofalpineRhithrogena(Ephemeroptera: Heptageniidae) using a mixed yule-coalescent analysis of mitochondrial and nuclear DNA. PloS One, 2011, 6(5): e19728.
[87] Vogler A P, and Monaghan M T. Recent advances in DNA taxonomy. Journal of Zoological Systematics and Evolutionary, 2007, 45(1):1-10.
[88] Hebert P D N, Ratnasingham S, de Waard J R. Barcoding animal life: cytochrome c oxidase subunit 1 divergences among closely related species. Proceedings of the Royal Society B: Biological Science, 2003, 270 (Sl): S96-S99.
[89] de Wit P, and Erséus C. Genetic variation and phylogeny of Scandinavian species ofGrania(Annelida: Clitellata: Enchytraeidae), with the discovery of a cryptic species. Journal of Zoological Systematics and Evolutionary Research, 2010, 48(4): 285-293.
[90] Cecha G, Borosb G, Dózsa-Farkas K. Revision ofBryodrilusglandulosus(Dózsa-Farkas, 1990) andMesenchytraeuskuehneltiDózsa-Farkas, 1991 (Oligochaeta: Enchytraeidae) using morphological and molecular data. Zoologischer Anzeiger, 2012, 251 (3): 253-262.
[91] Collado R, Hass-Cordes E, Schmelz R M. Microtaxonomy of fragmentingEnchytraeusspecies using molecular markers, with a comment on species complexes in enchytraeids. Turkish Journal of Zoology, 2012, 36(1): 85-94.
[92] van Vliet P C J, Beare M H, Coleman D C, and Hendrix P F. Effects of enchytraeids (Annelida: Oligochaeta) on soil carbon and nitrogen dynamics in laboratory incubations. Applied Soil Ecology, 2004, 25(2): 147-160.
[93] Topoliantz S, Ponge J F, Viaux P. Earthworm and enchytraeid activity under different arable farming systems, as exemplified by biogenic structures. Plant and Soil, 2000, 225(1/2): 39-51.
[94] Maraldo K. Enchytraeidae (Oligochaeta) in a changing climate-ecology and ecophysiology of enchytraeids exposed to climate changes. Copenhagen: Press of National Environmental Research Institute of Aarhus University, 2009: 1-131.
Systematics of the enchytraeidae (annelida, clitellata): past, present and future
CHEN Jing1, JIANG Wanxiang1, SHEN Qi2, XIE Zhicai2,*
1CollegeofLifeSciences,ZaoZhuangUniversity,Zaozhuang277160,China2TheKeyLaboratoryofAquaticBiodiversityandConservation,InstituteofHydrobiology,ChineseAcademyofScience,Wuhan430072,China
Enchytraeidae is the second largest family of the annelid class Clitellata, with nearly 650 species in 32 genera having been recognized up to now. These small but pretty potworms are almost cosmopolitan, being distributed in all major habitats including soils, oceans, fresh water, estuaries and glaciers, but with the majority of species being terrestrial (ca. 500) or marine (ca. 100). In spite of the long duration (over 200 years) of enchytraeid taxonomic investigation, the Enchytraeidae is still one of the most poorly-understood group of clitellates. Here, we provide an overview of the state of taxonomy and phylogenetics of the Enchytraeidae, then characterize its key morphological traits for enchytraeid identification, determine its main taxonomic problems and impediments, and provide future taxonomic and systematic perspectives. Firstly, enchytraeid systematics is still at its initial stage (αtaxonomy). This stage mainly concentrates on species exploration and validation. These species identification processes have been carried out through morphological analysis and consideration of the biological species concept. However, the high interspecific and/or intraspecific diversity of these traits, most of which are only observed and recorded in live condition, adds difficulty to enchytraeid identifications. Moreover, reproduction isolation of enchytraeids has rarely been tested, thus the hypothesis of established enchytraeid species in the biological species concept can not be confirmed. Second, phylogenetic studies of Enchytraeidae based on morphological-and-molecular data suggest that most of the enchytraeid genera are polyphyletic, indicating the established taxonomic systems yet do not genuinely reflect their evolutionary history. Thirdly, enchytraeid taxonomy faces such taxonomic problems and impediments as accelerating species exploration and testifying species validity, the establishment of genera/species definitions that reflect their real phylogenetic relationships, the application of DNA taxonomy to enchytraeid identification and reconstruction of intra-family phylogenies. Other impediments also constrain the develepment of enchytraeid taxonomy, such as the species-exploration imbalance across different regions of the world, the scarcity of funds and talented experts and the deficiency of electronically available taxonomic resources. Finally, we maintain that the application of both molecular data and the phylogenetic species concept should be the future direction of the enchytraeid taxonomy and systematics. Through reading the information stored in genes (such as COI gene), we can reconstruct the evolutionary history of enchytraeid ancestor struggling with their palaeoenvironment and the key processes and driving forces that enchytraeid ancestor had inherited their merit properties to their descendants. An enchytraeid taxonomic system based on the phylogenetic species concept should be tested and verified, e.g., by testing for reproductive isolation. Moreover, it is an important duty that enchytraeid experts disseminate advanced enchytraeid taxonomic knowledge to the public that can promote such enchytraeid-related research and recognitions of their roles in the processes of soil decomposition and nutrient mineralization, assessing soil ecosystem health condition, and evaluating global climate change.
enchytraeidae; taxonomy; phylogeny; identification; taxonomic impediment; perspective
国家自然科学基金(31272270,31100388); 山东省自然科学基金(ZR2010CL004, ZR2011CQ012)
2013-06-08;
日期:2014-05-16
10.5846/stxb201306081470
*通讯作者Corresponding author.E-mail: zhcxie@ihb.ac.cn
陈静,蒋万祥,沈琦,谢志才.线蚓科分类学研究进展.生态学报,2015,35(8):2461-2472.
Chen J, Jiang W X, Shen Q, Xie Z C.Systematics of the enchytraeidae (annelida, clitellata): past, present and future.Acta Ecologica Sinica,2015,35(8):2461-2472.
猜你喜欢
课堂内外·教师版(2022年4期)2022-05-23
江苏农业科学(2019年5期)2019-09-02
生物学通报(2019年7期)2019-07-08
广东农业科学(2017年5期)2017-08-29
天文爱好者(2016年7期)2016-12-20
中央民族大学学报(自然科学版)(2016年2期)2016-06-27
传感技术学报(2016年1期)2016-03-22
环境与生活(2016年1期)2016-03-17
新疆大学学报(哲学社会科学版)(2015年6期)2015-10-12
中央民族大学学报(自然科学版)(2015年3期)2015-06-11
