
鼠脊髓缺血再灌注损伤外周Th17/Treg细胞的变化与意义
2015-03-10 09:02:50姚平波蒋福生刘贵香
中风与神经疾病杂志 2015年2期
欧 军,姚平波,蒋福生,胡 聂,刘贵香,张 平
脊髓缺血再灌注损伤是一类脊髓神经元组织循环得到改善后乃出现的延迟性神经功能障碍或不可逆性迟发性神经元损伤[1]。目前认为参与脊髓缺血再灌注损伤发生的机制主要有免疫机制、血管机制、自由基损伤学说、钙介导机制、炎性反应及细胞凋亡等复杂网络演化机制调控其神经元细胞的自身活化及极化状态,也调控神经再生和修复功能[2]。辅助性T 细胞17(T-helper17 cells,Th17)是近年来发现的一类新型Th 细胞亚群,其介导炎性反应,参与机体的免疫应答。调节性T 细胞(regulatory T cells,Treg)是能够抑制免疫应答的重要细胞,具有抑制炎症反应的作用,能够下调免疫反应[3]。Treg细胞和Th17 细胞通过细胞因子的作用相互转换,从而调节机体的免疫抑制与免疫效应并使之处于精细而复杂的动态平衡中[4]。Th17/Treg 保持平衡,有利于机体免疫稳态的维持,如果Th17/Treg 失平衡就会导致疾病发生发展乃至恶化[5~7]。但目前国内外有关Th17/Treg 平衡在大鼠脊髓缺血再灌注损伤中的变化、意义及其调控机制的研究未见文献报道。为此,笔者观察大鼠脊髓缺血再灌注损伤Thl7/Treg的特异性转录因子及相关细胞因子的变化,为阐明大鼠脊髓缺血再灌注损伤免疫紊乱的细胞与分子机制提供新思路及线索。
1 材料和方法
1.1 实验动物及分组 健康雄性Wistar 大鼠40 只,体质量(400 ±20)g(由南华大学比较医学科提供)。购进大鼠后进行1 w 的适应性饲养,温度控制在20 ℃~28 ℃。随机分为对照组(10 只)和脊髓缺血再灌注组(40 只)。
1.2 脊髓缺血再灌注损伤模型制备 采用1%氯胺酮100 mg/kg 腹腔注射麻醉,固定于实验台上。按昆明医学院附属昆华医院李艳华教授首创的大鼠肾下腹主动脉结扎并切断肾上腹主动脉发出的椎动脉,制备脊髓缺血再灌注损伤模型[8]。术后静脉缓慢推注青霉素预防感染。大鼠完全清醒后放入单笼饲养。所有大鼠于缺血再灌注后重新麻醉,进行取材并-70 ℃冰箱保存。脊髓缺血再灌注损伤成功模型制作的标准:大鼠苏醒后痉挛性摆动尾巴能正常进食,前肢功能正常,双下肢瘫痪伴不同程度的大小便功能障碍。
1.3 转录因子RORγt 和Foxp3 mRNA 的表达自外周血分离淋巴细胞,Trizol 一步法提取总RNA,按照试剂盒说明书步骤反转录cDNA 及行荧光RT-PCR 检测Foxp3 mRNA 和RORγt mRNA 水平表达。各基因引物序列及扩增产物长度(见表1)。采用标准曲线法计算mRNA 表达量,并计算其相对表达量(β-actin 校正值)。引物均由英潍捷基(上海)贸易有限公司合成。
1.4 血清细胞因子的测定 解冻-70 ℃保存的脊髓组织,裂解组织(裂解液4 ml/g),冰浴超声匀浆,于4 ℃、3000 r/min 离心15 min 后收集上清液,按照ELISA 试剂盒说明书检测白细胞介素-17 (interleukin-17,IL-17)、IL-6 及IL-10 检测步骤,使用全自动酶标仪读取波长450 nm 处的A 值。通过绘制标准曲线求出标本中的IL-10、IL-6 和IL-17 的浓度。
1.5 统计学分析 采用SPSS 19.0 统计软件,数据以均数± 标准差(χ ± s)表示,采用One-Way ANOVA 检验,P <0.05 或P <0.01 为差异有统计学意义。
2 结果
2.1 转录因子Foxp3 mRNA 及RORγt mRNA的表达情况 脊髓缺血再灌注后大鼠组织中的Foxp3 mRNA 的表达急剧下降,在第3 天时表达量降到最低,之后随着时间的延长出现再升的趋态;RORγt mRNA 表达量在第3 天时升至最高,之后随着时间的延长出现再降的趋态(见表2)。
2.2 各组细胞因子之间的比较 脊髓缺血再灌注后各组IL-10 含量与对照组比较均明显升高(P<0.01),在第3 天为最高。脊髓缺血再灌注后第1天其IL-6 及IL-17 含量均高于对照组(P <0.01),但IL-10 在脊髓缺血再灌注后其动态演变呈双相变化趋势,即在肠缺血再灌注后第3 天和第5 天分别出现两个波谷。而脊髓缺血再灌注后第3 天和第7天其IL-6 及IL-17 含量处于较高水平,与第1 天比较有显著性差异(P <0.05,见表3)。
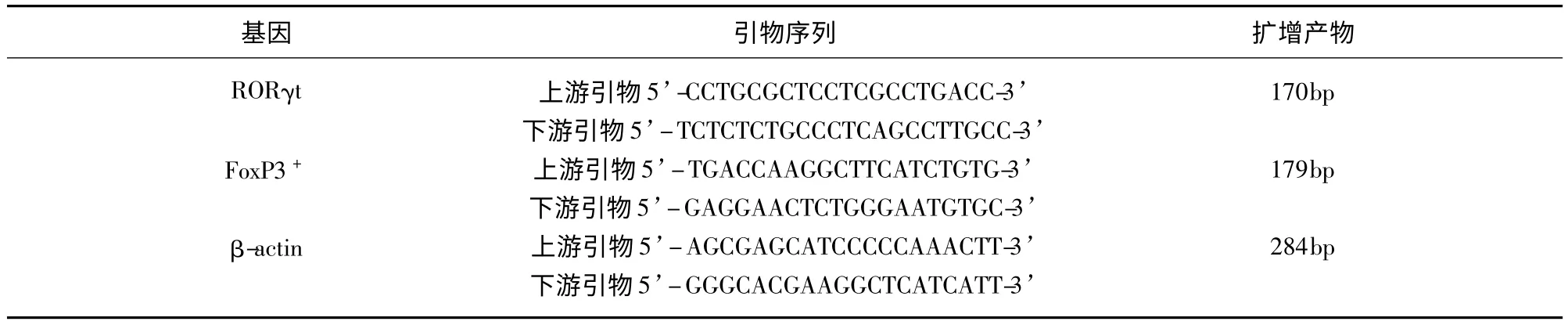
表1 各基因引物序列及扩增产物长度
表2 转录因子Foxp3 mRNA 及RORγt mRNA 的表达()
表3 各组之间细胞因子的比较(,ng/ml)
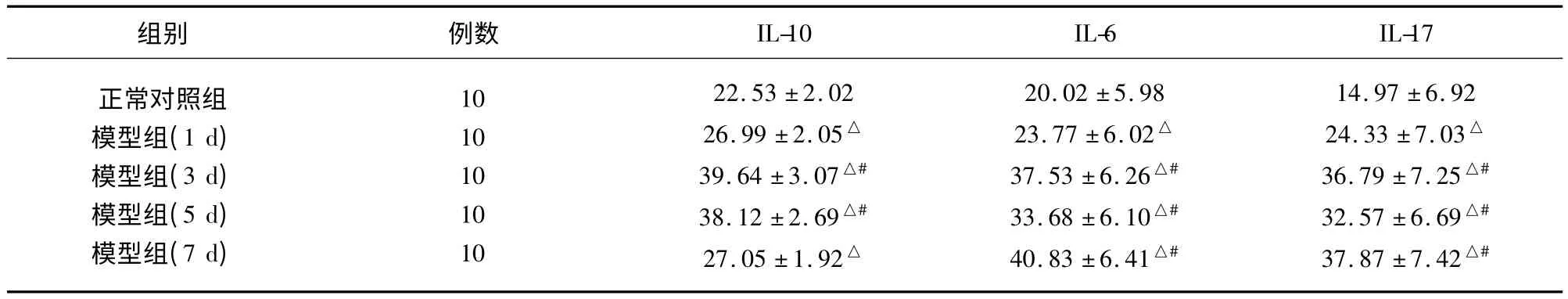
表3 各组之间细胞因子的比较(,ng/ml)
3 讨论
Treg 细胞是一种具有调节机体免疫反应和诱导免疫耐受的T 细胞亚群。Treg 细胞的免疫抑制性是其最重要特性之一,能抑制效应性CD4+T 细胞增生与活化[9,10]。Th17 细胞介导炎性反应,参与机体的免疫应答,Treg 细胞与Th17 细胞保持平衡有利于机体免疫稳态的维持。Th17 细胞与Treg 细胞之间稳态的破坏是很多炎症反应与自身免疫性疾病发生的关键因素[11,12]。
脊髓缺血再灌注损伤机制尚不十分明确。随着不断深入的研究,人们逐渐关注IL-10/IL-17 或IL-6/IL-17 轴在其发生及发展中的重要意义。本实验研究表明,脊髓缺血再灌注损伤不同时间点的RORγt mRNA、Foxp3 mRNA 及细胞因子的表达与对照组比较,差异具有统计学意义(P <0.05 或P <0.01)。这说明在脊髓缺血再灌注损伤组织中Th17细胞/Treg 细胞比例失衡,Treg 细胞下调,免疫调节功能下降,导致效应性CD4+T 细胞大量活化,产生大量致炎因子,导致免疫功能紊乱,Th17 细胞数也随之减少。Th17 细胞表达水平过低,从而导致机体抵抗能力下降,但Th17 细胞过度表达可能引发炎性反应和自身免疫应答。故适当提高缺血再灌注损伤组织Treg 的水平,将有助于改善其免疫功能。
此外随着脊髓缺血再灌注损伤的发生及发展,其机体细胞因子及转录因子表达的动态演变如何,也是本实验深入研究的方向和内容,以期全面系统地观察Th17、Treg 及其上游调控因子与脊髓缺血再灌注损伤病程的相关性。在本实验中,通过qRTPCR 技术检测RORγt mRNA 和Foxp3 mRNA 的表达及采用ELISA 方法,检测IL-10、IL-17、IL-6 的含量来反映脊髓缺血性再灌注损伤Th17/Treg 特异性转录因子动态演变及相关细胞因子的变化趋势。在观察的时间点内,第3 天时Foxp3 mRNA 和RORγt mRNA的表达出现一个不对称峰值时脊髓缺血再灌注损伤后细胞因子IL-10、IL-17、IL-6 出现在高峰期说明机体通过表达Foxp3 诱导T 淋巴细胞分化Treg 细胞并促进IL-10 的分泌发挥抗炎及免疫抑制的作用;促炎因子IL-17 是Th17 细胞标志性的细胞因子,启动炎症的发生,促进炎性介质IL-6 的释放加剧炎性反应。在实验中还观察到第7 天呈Foxp3 mRNA 呈上升趋势,与此同时RORγt mRNA 的表达在7 d 开始下降,而IL-6、IL-17 及IL-10 含量乃处于较高水平,这可能与脊髓缺血再灌注损伤的机制的复杂化有关,且随着时间的延长,其他的相关因素参与到脊髓缺血再灌注损伤发生发展环节中来,导致一系列机体病理生理及内环境稳态的变化,从而导致器官的损伤和全身炎性反应。
综上所述,Treg 细胞与Th17 细胞发展的不平衡性在脊髓缺血再灌注损伤的发生发展中起关键性作用。这种稳态的破坏既使免疫系统激活,又使免疫效应不足,这种双向变化,进一步加剧了炎症反应,加剧了炎性反应状态的发展。这提示:调控Th17 细胞的活化并增强Treg 细胞的活性使其处于稳态之中可以调节机体的免疫状态和抑制炎症反应状态的进展,从而为脊髓缺血再灌注损伤的靶向治疗提供了新的思路和线索。
[1]黄 飞,李亚男,尹 飞,等.脊髓缺血再灌注损伤对大鼠水通道蛋白4 表达的影响[J].中国免疫学杂志,2013,29(5):495 -498.
[2]宋庆鑫,张 帆,王 琨,等.miRNA 在急性脊髓损伤病理生理调节机制中的作用研究进展[J].中国脊柱脊髓杂志,2013,23(11):1022 -1024.
[3]Lu P,Wang M,Zheng P,et al.Th17/Treg unbalance is involved in the pathogenesis of experimental autoimmune encephalomyelitis[J].Chinese Journal of Cellular and Molecular Immunology,2014,30(10):1013 -1017.
[4]Ma L,Liang Y,Fang M,et al.The cytokines (IFN-gamma,IL-2,IL-4,IL-10,IL-17)and Treg cytokine (TGF-beta1)levels in adults with immune thrombocytopenia[J].Die Pharmazie,2014,69(9):694 -697.
[5]刘贵香,姚平波,张 平,等.急性脑梗死患者外周血Th17 细胞与调节性T 细胞的表达水平与预后的关系[J].广东医学,2014,35(7):1027 -1029.
[6]胡 聂,包 铮,王 松,等.严重胸部创伤患者外周血Th17/Treg平衡变化与预后的关系[J].广东医学,2014,35(7):1095 -1097.
[7]蒋福生,张 平,张新华,等.急性脑梗死患者免疫调节性T 淋巴细胞表达与预后的关系[J].中国动脉硬化杂志,2013,21(10):923 -926.
[8]金 华,郭光琼,李艳华,等.脊髓缺血再灌注损伤模型的改进及对大鼠神经行为学的影响[J].中风与神经疾病杂志,2012,29(10):879 -882.
[9]Dong L,Wang X,Tan J,et al.Decreased expression of microRNA-21 correlates with the imbalance of Th17 and Treg cells in patients with rheumatoid arthritis[J].J Cellular and Molecular Medicine,2014,28(10):1 -12.
[10]Nogueira LG,Santos RH,Fiorelli AI,et al.Myocardial gene expression of T-bet,GATA-3,Ror-gammat,FoxP3,and hallmark cytokines in chronic Chagas disease cardiomyopathy:an essentially unopposed TH1-type response[J].Mediators of Inflammation,2014,2014:914326.
[11]Edwards JP,Thornton AM,Shevach EM,et al.Release of active TGFbeta1 from the latent TGF-beta1/GARP complex on T regulatory cells is mediated by integrin beta8[J].J Immunol,2014,193(6):2843 -2849.
[12]Hu R,Huffaker TB,Kagele DA,et al.MicroRNA-155 confers encephalogenic potential to Th17 cells by promoting effector gene expression[J].J Immunol,2013,190(12):5972 -5980.
猜你喜欢
军事文摘(2022年8期)2022-11-03 14:22:01
大电机技术(2022年3期)2022-08-06 07:48:24
核科学与工程(2021年4期)2022-01-12 06:30:04
天津医科大学学报(2021年3期)2021-07-21 09:03:48
煤气与热力(2021年4期)2021-06-09 06:16:54
中华戏曲(2020年1期)2020-02-12 02:28:18
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
医学研究杂志(2015年9期)2015-07-01 17:28:19
中国中医药现代远程教育(2014年21期)2014-03-01 04:32:11
中国中医药现代远程教育(2014年20期)2014-03-01 04:31:33
