
烟台海岸防护林带6树种资源利用效率比较
2015-03-07 02:06刘玉静董周焱柏新富卜庆梅
山东农业大学学报(自然科学版) 2015年4期
关键词:树种
刘玉静,董周焱,柏新富,卜庆梅
鲁东大学生命科学学院,山东烟台264025
烟台海岸防护林带6树种资源利用效率比较
刘玉静,董周焱,柏新富,卜庆梅*
鲁东大学生命科学学院,山东烟台264025
摘要:树种单一是造成我国沿海防护林生态系统脆弱、功能难以发挥的主要因素。对海岸多变环境条件下不同树种的生长发育潜力进行评估有利于沿海防护林建设的树种选择。本实验用TPS-1便携式光合系统测定烟台海岸带6个树种柽柳、黑松、龙柏、白蜡、廊坊杨和刺槐的光合性能指标,同时测定了其碳素利用效率、光合氮利用效率和灰分含量等,并用隶属函数法计算了6个树种的综合资源利用效率,以评定其生长发育潜力。结果表明,6个树种的饱和光强都较高,均适宜在较强光照条件下生长,但柽柳、黑松和龙柏的表观光量子效率、光合速率和光能利用效率均显著低于白蜡、廊坊杨和刺槐,其中尤以龙柏最低。同时,柽柳、黑松和龙柏的光合氮利用效率和叶片碳利用效率也显著低于白蜡、廊坊杨和刺槐,但是其水分利用效率则普遍高于后者;矿质元素利用效率则是柽柳最高、黑松最低,其它4个树种居中。将6个树种对各种环境资源的利用效率与光合性能指标进行综合分析并计算资源利用效率综合指数显示,6个树种中刺槐和白蜡资源利用效率最高、廊坊杨和柽柳次之、黑松再次、龙柏最低。
关键词:光合性能;资源利用效率;海岸防护林;生长潜力;树种
植物的资源利用效率(Resource use efficiency, RUE)是指植物对其生长所依赖的光能、水分、营养元素等的利用效率。其中,光能利用效率(Light use efficiency, LUE)高表明其有高的生产力和快速的生长能力[1];高的水分利用效率(Water use efficiency, WUE)则有利于其在干旱条件下获得高的生物产量[2];高的碳素利用效率(Carbon use efficiency,CUE)说明植物将吸收的碳素更多地转化为生长和贮存的物质[3];高的光合氮利用效率(Photosynthetic nitrogen use efficiency, PNUE)通常与其高的生长速率相关联[4],也就是说,资源利用效率高的植物往往具有高的生长潜力[5,6]。
沿海防护林是国家防护林体系的重要组成部分,是沿海地区的绿色屏障[7],担负着防风固沙、阻挡海雾、保护农田和改善海岸生态状况等重要作用[8]。然而,由于海岸植物常常受到风、沙、旱、盐等非生物因素的交叉或多重胁迫,使得很多植物难以在海岸恶劣环境条件下很好地生长,从而形成了现有沿海防护林树种单一、结构模式简单、生态系统脆弱等诸多问题,其中树种单一是最主要因素[9]。因此,树种多样性已成为现代海岸防护林体系建设的关键。但是,有关海岸防护林树种的选择目前仅在树种对海岸环境的适应性方面有些研究[10,11],对其它树种生长潜力的研究也多见于有关树高、胸径、生物量(蓄积量)等方面的报道[12,13]。本试验通过对黄海北部海岸带主要树种的光合性能、光能利用效率、水分利用效率和碳、氮利用效率等的测定和计算,从资源利用效率的角度对海岸多变的恶劣环境条件下各树种的生长潜力进行评价,为沿海防护林体系建设选择既能适应海岸恶劣环境条件又能保持最大限度生长的树种提供参考。
1 材料与方法
1.1研究区域自然条件和试验区土壤基本状况
试验在处于烟台市区北部海岸防护林带的芝罘林场进行。烟台沿海属温带季风气候,夏季凉爽,冬季温润;年平均降水量为651.9 mm,年平均气温11.8℃,年平均相对湿度68%,太阳辐射总量年平均值5224.4兆J/m2,年平均风速4~6 m/s,平均无霜期210 d。试验区距高潮线约30 m,从高潮线到试验区依次为裸露沙滩、前沿草带、试验林区。试验区土壤为滨海沙土,基质由疏松的中、粗沙组成,土壤有机质含量为1.01%,速效氮、磷、钾分别为56.8 mg/kg、0.8 mg/kg和52.5 mg/kg,pH值为5.8。
1.2试验材料
试验选择试验林区同一林地内同期栽种的6个树种柽柳(Tamarix chinensis Lour.)、黑松(Pinus thunbergii)、龙柏(Juniperus chinensis cv. kaizuka)、白蜡(Fraxinus chinensis Roxb.)、廊坊杨(Populus langfanggensis)和刺槐(Robinia pseudoacacia)为研究对象。所有树种均为4年生植株,各树种南北成行东西向混合排列(每行1个树种),行距1.5 m、株距1 m。每个树种选择5行每行选择1株健康植株为测定对象,于2014年5月下旬、7月下旬和9月下旬进行3次测定和取样。测定和取样均以植株向阳面中部当年生枝条近顶端的成熟叶(同化枝)为材料(黑松5月下旬测定时当年生叶还没发育成熟,故取上一年的针叶)。
1.3试验方法
1.3.1光合作用指标和水分利用效率、叶片碳素利用效率的测定与计算光合作用相关指标的测定在上午8:30~11:30进行。使用TPS-1便携式光合系统(PP System公司,美国)测定记录不同光照强度(光合有效辐射PAR,以LED光源控制)下净光合速率(Pn)、蒸腾速率(Tr)、呼吸速率(R)等指标(使用通用叶室,测定龙柏、黑松和柽柳时为防止叶室漏气用凡士林进行密封,其叶面积测定按肖强等[14]的方法拍照并用Photoshop软件分析计算)。每个树种每次测定5个单株,每个单株测定1次。参照Ye[15]的直角双曲线修正模型计算最大净光合速率(Pmax)、光饱和点(Light saturation point, LSP)和光补偿点(Light compensation point, LCP),并利用曲线初始斜率计算表观光量子效率(Apparent quantum yield,AQY);参照王珊珊等[16]的方法计算光能利用效率[Light use efficiency, LUE(molCO2·mol-1proton)=Pn/PAR]、水分利用效率[Water use efficiency, WUE(mmol·mol-1)=Pn/Tr];参照Frantz等[17]的方法用净光合速率与总光合速率(净光合速率Pn+呼吸速率R)的比值表示叶片碳利用效率[Carbon use efficiency, CUE(mol·mol-1)=Pn/(Pn+R)]。
1.3.2光合氮利用效率测定与计算分别采集每个树种的5个单株的成熟叶片(同化枝)带回实验室(每个单株单独取样和测定),用烘干称重法测定计算比叶重LMA(g/m2)=叶片干重/叶面积(白蜡、廊坊杨和刺槐的叶面积用纸称重法测定,龙柏、黑松和柽柳的叶面积用拍照和Photoshop软件计算[14])。用Vario Micro cube小进样量元素分析仪(Elementar公司,德国)测定叶片全氮含量(以单位质量的叶氮含量表示,Nmass)。光合氮利用效率(Photosynthetic nitrogen-use efficiency,PNUE)参照苗艳明等[18]的方法计算,即PNUE(μmolCO2·mol-1·s-1)=Pmax/(1/14 Nmass×LMA)。式中,乘以1/14计算氮摩尔分子量,Nmass—叶片含氮量(g/g样品重量),LMA—比叶重(g/m2)。
1.3.3灰分含量和矿质元素利用效率测定利用马福炉干灰化法测定。分别采集每个树种的5个单株的成熟叶片(同化枝)带回实验室(每个单株单独取样和测定),在65~70℃烘干72 h,磨粉后精确称取1 g左右样品,用马福炉在700℃下灰化7 h,计算灰分含量(Ash content,AC),AC(mg/g)=灰分质量×1000/样品质量。灰分物质主要是植物直接从土壤吸收的一些矿质元素,本实验直接用灰分含量表示植物矿质元素利用效率。
1.4资源利用效率的综合指数的计算
参照隶属函数法[19]求资源利用效率综合指数(Comprehensive indices of resource use efficiency, CRI)。其中某一指标的资源利用效率指数(Index of one resource use efficiency)。xi为某一指标的各测定值,xmax为该指标各测定值的最大者,xmin为该指标各测
定值的最小者。
1.5数据分析方法
本试验中每个树种的每项指标均在3个时期各测定5次(每个时期每个树种重复测定5个单株),结果以“平均值±标准差”计,用SPSS17.0的“General Linear Model—Univariate和Duncan”以树种和测定时期做两因素方差分析和相互之间的差异显著性检验。
2 结果与分析
2.1光合性能指标的比较
6个树种光合性能指标及其方差分析结果列于表1和表2。从表1可见6个树种中鳞针叶树种(柽柳、黑松和龙柏)的表观光量子效率(AQY)和最大净光合速率(Pmax)显著低于其它阔叶树种,而其光饱和点(LSP)和光补偿点(LCP)则显著高于后者。其中,龙柏的光合效能(AQY和Pmax)最低,廊坊杨和刺槐的光合效能较高;柽柳、黑松和廊坊杨利用强光的能力较强,而白蜡则适应弱光的能力相对较高。说明在6个树种中,龙柏光能转化效率较低,柽柳则是利用弱光的能力较差。另从表2可见,不同测定时期相互之间光合性能指标(AQY、Pmax、LSP和LCP)差异不显著(P=0.079~0.844>0.05)。
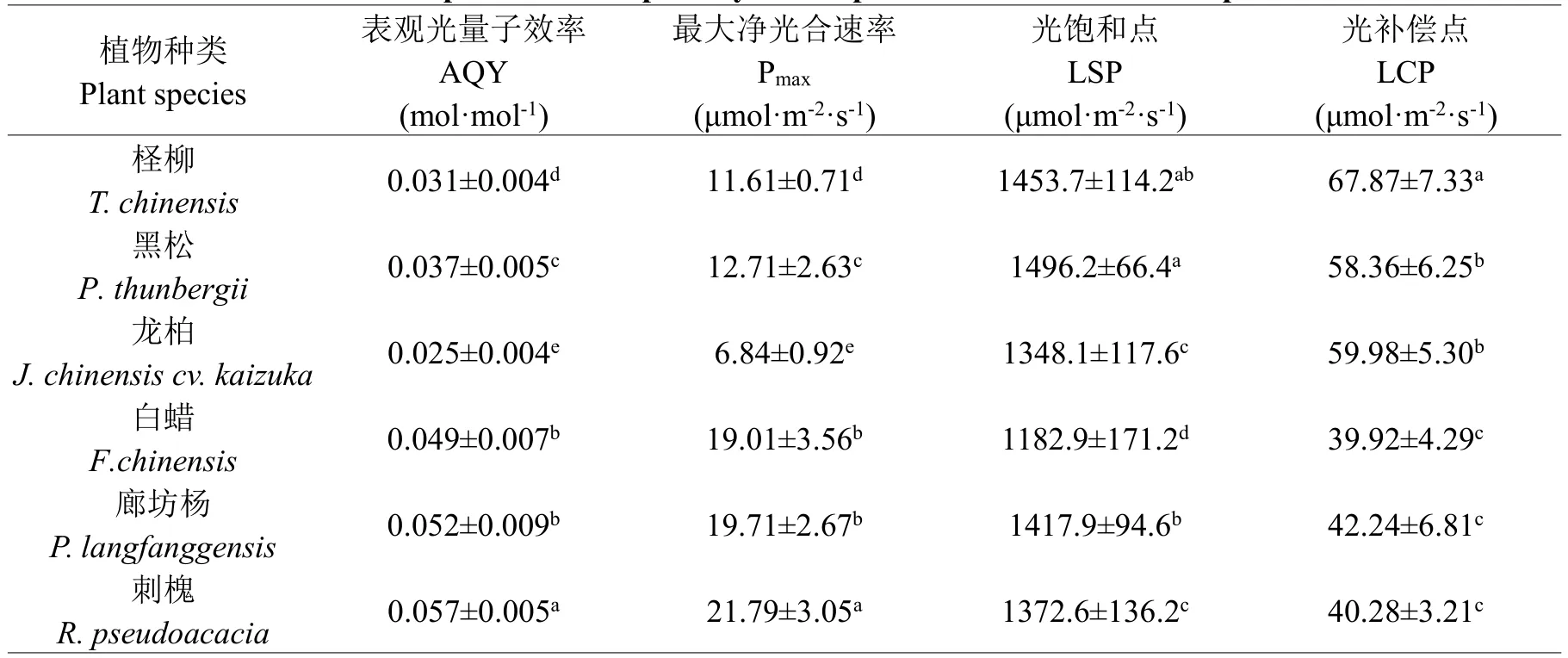
表1 6个树种的光合性能指标Table 1 Comparison of the photosynthetic performance in the 6 tree species

表2 不同树种不同测定时期资源利用效率指标的两因素方差分析表Table 2 Two-way ANOVAon the resource use efficiency of the tree species in different period of determination
2.2光能利用效率和水分利用效率
对光能利用效率和水分利用效率的分析、比较显示,6个树种的光能利用效率(LUE)差异显著(P=0.032<0.05,表2),其中:廊坊杨和刺槐的光能利用效率最高、白蜡次之、3个鳞针叶树种较低(表3),尤以龙柏最低,仅为最高者的30.65%。水分利用效率在不同树种之间差异不显著(P=0.701>0.05,表2),其中,柽柳和龙柏的水分利用效率相对较高,黑松、白蜡和刺槐稍低,廊坊杨最低(表3)。
2.3碳、氮和矿质元素利用效率
经方差分析和多重比较显示,本试验中6个树种的叶片碳利用效率(CUE)、光合氮利用效率(PNUE)和矿质元素利用效率(MUE)在不同树种间均有极显著差异(表2,P=0.000~0.006<0.01)。叶片碳利用效率总体表现为鳞针叶树种显著低于阔叶树种(表3),其中阔叶树种中白蜡最高、鳞针叶树种中龙柏最低,白蜡比龙柏高14.6%;光合氮利用效率也表现为鳞针叶树种显著低于阔叶树种(表3),其中阔叶树种中刺槐最高、鳞针叶树种中龙柏最低,龙柏仅为刺槐的5.5%,两者相差约18倍;矿质元素利用效率则表现为柽柳远高于其它树种,为黑松的约4倍、其它树种的近2倍,除柽柳和黑松外,其它树种相互之间差异较小(表3)。
另外,统计分析还显示(表2):不同测定时期的LUE、CUE、PNUE差异不显著(P=0.095~0.614),而MUE和WUE在不同时期则有显著差异和极显著差异(P=0.015和0)。其中,水分利用效率5月下旬最高、9月下旬次之、7月下旬最低;叶片碳利用效率也是5月下旬最高、9月下旬和7月下旬相对较低。

表3 6个树种的资源利用效率比较Table 3 Comparison of the resource use efficiency in the 6 tree species
2.4资源利用效率的综合分析
植物对环境资源的利用过程复杂,每种植物对各种环境资源的利用都有不同的倾向,某一种参数或指标所表达的仅是这一因素的信息,因此,任何单一指标或参数都不可能很好地评定植物的资源利用效率。为此,本实验应用隶属函数法对6个树种的各种环境资源利用效率进行综合分析(表4),结果表明,6个树种的资源利用效率综合指数可分为4个层次:刺槐和白蜡最高、廊坊杨和柽柳第二、黑松第三、龙柏最低。

表4 6个树种的资源利用效率指数RIi值及资源利用效率综合指数CRI值Table 4 The RIiand the CRI in the 6 trees species
3 讨论
植物的生长发育涉及对光能、水分、氮素、碳素、矿质元素等环境资源的综合利用过程[20],资源利用效率的高低在一定程度上影响其生存、生长和繁育能力,决定其对环境的适应能力和竞争力[21,22]。Funk等[6]将植物的资源利用效率定义为植物生长发育过程中利用每单位资源所同化的碳素量或积累的生物量。植物的碳素同化是通过光合作用来实现的,因此,光合作用特性对植物的资源利用效率有很大的影响。本试验对柽柳、黑松、龙柏、白蜡、廊坊杨和刺槐等6个树种光合性能的比较发现,它们的饱和光强都较高,均适宜在较强光照条件下生长,但3个鳞针叶树种的光能转化效率和光合速率均显著低于其它阔叶树种,而其光补偿点则显著高于后者,说明鳞针叶树种的光合能力不及阔叶树种,其中尤以龙柏最低。
光能利用效率是决定植物生产力高低的最重要因素[1],它反映植物对光的利用能力。Durand等[23]认为植物对光能的捕获和利用效率高低是其能否适应环境并很好地生存下去的决定性因素。水分利用效率作为叶片光合与水分生理过程的耦合因子,既是联系碳循环与水循环的重要变量,亦是研究和评价植物生长和适应性的综合生理生态依据之一[24,25]。光合氮素利用效率则反映植物利用氮营养和合理分配氮的能力,是氮对植物光合生产力乃至生长产生影响的重要指标,光合氮利用率较高的植物生长较快[25,26]。本试验中,3个阔叶树种刺槐、白蜡和廊坊杨的光能利用效率和光合氮利用效率显著高于3个鳞针叶树种柽柳、龙柏和黑松,而水分利用效率则是鳞针叶树种显著高于阔叶树种。相关分析显示,6个树种的光能利用效率与光合氮利用效率有显著正相关(相关系数r=0.855, P<0.05)、与水分利用效率则具有显著的负相关(r=﹣0.910, P<0.05)、光合氮利用效率与水分利用效率也具有一定的负相关性(r=﹣0.691, P=0.13)。有研究表明,蒸腾耗水能力越强,植物对水分利用效率就越低,但光能利用效率和光合氮利用效率则越高[27,28]。本试验结果与此相符,也就是说,高的光能利用效率和光合氮利用效率是以消耗较多的水分为代价的。
植物碳利用效率表示植物碳的收支情况,反映总光合产物的分配格局[29]。Bradford等[3]认为高的碳素利用效率表明植物将吸收的碳素更多地转化为生长和贮存的营养物质,有利于植物的生长。而灰分含量(矿质元素利用效率)的高低可反映不同植物对矿质元素选择吸收与积累的特点,灰分含量的增加代表植物的代谢较为旺盛、生长较为迅速、竞争力增强[30,31]。宋莉英等[32]的研究结果也显示,较高的灰分含量的植物其生物量建成成本较低,竞争力强。本试验中,鳞针叶树种的叶片碳素利用效率显著低于阔叶树种(P<0.05),其中龙柏最低;矿质元素利用效率则是柽柳远高于其它树种、黑松远低于其它树种,龙柏、白蜡、廊坊杨和刺槐相互之间差异较小。这一结果表明3个阔叶树种贮存、转化碳素的能力和生长势强于3个鳞针叶树种,但柽柳能够尽可能多地吸收矿质元素以低成本策略在海岸环境下生长,黑松和龙柏则生长、竞争力较弱。
由于某一资源利用效率所表达的仅是植物对这一因子利用的倾向,为了准确评价不同种类植物的生长潜力,本文引入了资源利用效率综合指数(CRI)作为衡量指标。通过对资源利用效率综合指数的计算发现,6个树种的资源利用效率综合指数由高到低排序为:刺槐和白蜡、廊坊杨和柽柳、黑松、龙柏。也就是说,在黄海北部沙质海岸环境条件下,本试验研究的6个树种中刺槐和白蜡生长潜力最高、廊坊杨和柽柳次之、黑松再次、龙柏最低。当然,在海岸防护林建设树种选择时,除了生长潜力,还要参考各树种的抗盐性、抗风能力等,以便科学合理地规划和配置各树种在海岸防护林体系中的适宜层次和位置。
参考文献
[1] Binkley D, Laclau JP, Sterba H. Why one tree grows faster than another: Patterns of light use and light use efficiency at the scale of individual trees and stands[J]. Forest ecology and management, 2013,288:1-4
[2]梅旭荣,钟秀丽,刘晓英.探讨品种间差异改良作物水分利用效率[J].作物学报,2013,39(5):761-766
[3] Bradford MA, Crowther TW. Carbon use efficiency and storage in terrestrial ecosystems[J]. New Phytologist, 2013,199(1):7-9
[4] Hikosaka K. Interspecific difference in the photosynthesis-nitrogen relationship: patterns, physiological causes, and ecological importance[J]. Journal of Plant Research, 2004,117(6):481-494
[5] Binkley D. Understanding the role of resource use efficiency in determining the growth of trees and forests[C]//Schlichter T, Montes L. Forests in Development: A Vital Balance. Netherlands: Springer, 2012:13-26
[6] Funk JL, Vitousek PM. Resource-use efficiency and plant invasion in low-resource systems[J]. Nature, 2007,446:1079-1081
[7]张志东,毛培利,刘玉虹,等.林分结构对烟台黑松海岸防护林天然更新的影响[J].生态学报,2010,30(8):2205-2211
[8]许景伟,李传荣,王卫东,等.沿海沙质岸黑松防护林的生物量及生产力[J].东北林业大学学报,2005,33(6):29-32
[9]黄敏参,杜晓娜,廖蒙蒙,等.东南沿海潮间带防护林主要树种的光合特性及水分利用策略[J].生态学杂志,2012,31(12):2996-3002
[10]李国旗,安树青,张纪林,等.海岸带防护林4种树木的风压应力分析[J].南京林业大学学报,1999,23(4):76-80
[11]韩玉洁,孙海菁,朱春玲,等.上海沿海防护林树种适应性评价[J].南京林业大学学报(自然科学版),2010,34(4):165-168
[12]姚延,白景萍,郝志刚,等.混农杨树林生长潜力及营养元素循环[J].山西农业大学学报,1998,18(4):310-315
[13] Hodge GR, Dvorak WS. Growth potential and genetic parameters of four Mesoamerican pines planted in the Southern Hemisphere[J]. Southern Forests: a Journal of Forest Science, 2012,74(1):27-49
[14]肖强,叶文景,朱珠,等.利用数码相机和Photoshop软件非破坏性测定叶面积的简便方法[J].生态学杂志,2005,24(6):711-714
[15] Ye ZP. A new model for relationship between irradiance and the rate of photosynthesis in Oryza sativa[J]. Photosynthetica, 2007,45(4):637-640
[16]王珊珊,陈曦,王权,等.新疆古尔班通古特沙漠南缘多枝柽柳光合作用及水分利用的生态适应性[J].生态学报,2011,31(11):3082-3089
[17] Frantz JM, Bugbee B. Acclimation of plant populations to shade: photosynthesis, respiration, and carbon use efficiency[J]. Journal of the American Society for Horticultural Science, 2005,130(6):918-927
[18]苗艳明,吕金枝,毕润成,等.翅果油树叶性特征的动态变化[J].植物学报,2012,47(3):257-263
[19] Guo QQ, Zhang WH, Li HE. Comparison of photosynthesis and antioxidative protection in Sophora moorcroftiana and Caragana maximovicziana under water stress[J]. Journal of Arid Land, 2014,6(5):637-645
[20] Griffin K. Calorimetric estimates of construction cost and their use in ecological studies[J]. Functional ecology, 1994,8(5):551-562
[21] Nagel JM, Griffin KL. Construction cost and invasive potential: comparing Lythrum salicaria (Lythraceae) with co-occurring native species along pond banks[J]. American Journal of Botany, 2001,88(12):2252-2258
[22] Gyenge J, Fernández ME. Patterns of resource use efficiency in relation to intra-specific competition, size of the trees and resource availability in ponderosa pine[J]. Forest Ecology and Management, 2014,312:231-238
[23] Durand LZ, Goldstein G. Photosynthesis, photoinhibition, and nitrogen use efficiency in native and invasive tree ferns in Hawaii[J]. Oecologia, 2001,126(3):345-354
[24] Richards RA, Rebetzke GJ, Condon AG, et al. Breeding opportunities for increasing the efficiency of water use and crop yield in temperate cereals[J]. Crop Science, 2002,42(1):111-121
[25]何彦芳,汤璐瑛,陈雅涵,等.北京及周边区域植物叶片6种生理属性研究[J].中国农业大学学报,2012,17(3):81-87
[26]郑淑霞,上官周平.不同功能型植物光合特性及其与叶氮含量、比叶重的关系[J].生态学报,2007,27(1):171-181
[27]张国芳,王北洪,孟林,等.四种偃麦草光合特性日变化分析[J].草地学报,2005,13(4):344-348
[28] Livingston NJ, Guy RD, Sun ZJ, et al. The effects of nitrogen stress on the stable carbon isotope composition, productivity and water use efficiency of white spruce (Picea glauca (Moench) Voss) seedlings[J]. Plant, Cell & Environment, 1999,22(3):281-289
[29] Dillaway DN, Kruger EL. Trends in seedling growth and carbon-use efficiency vary among broadleaf tree species along a latitudinal transect in eastern North America[J]. Global Change Biology, 2014,20:908-922
[30]郭水良,黄华,晁柯,等.金华市郊10种杂草的热值和灰分含量及其适应意义[J].植物研究,2005,25(4):460-464
[31] Villar R, Merino J. Comparison of leaf construction costs in woody species with differing leaf life‐spans in contrasting ecosystems[J]. New Phytologist, 2001,151(1):213-226
[32]宋莉英,彭长连,彭少麟.华南地区3种入侵植物与本地植物叶片建成成本的比较[J].生物多样性,2009,17(4):378-384
Comparison among Resource Use Efficiencies of 6 Tree Species in the Coastal Forest Shelterbelt of Yantai
LIU Yu-jing, DONG Zhou-yan, BAI Xin-fu, BU Qing-mei*
College of Life Science/Ludong University, Yantai 264025, China
Abstract:The scarcity of tree species is the main factor that results in the vulnerability and malfunction of coastal protection forest as an ecosystem. Assessing the growth potential of the various woody species under the harsh coastal environment will be beneficial to the selection of tree species for coastal shelterbelt construction. In this study, the photosynthetic performance indicators of 6 trees (namely Tamarix chinensis, Pinus thunbergii, Juniperus chinensis cv kaizuka, Fraxinus chinensis, Populus langfanggensis and Robinia pseudoacacia) were determined with a TPS-1 portable photosynthesis system in the coastal forest shelterbelt of Yantai. Meanwhile, the carbon use efficiency (CUE), photosynthetic nitrogen-use efficiency(PNUE), and the ash content were determined as well. Additionally, the comprehensive indices of resource use efficiency of 6 tree species were calculated with the subordinate function for the evaluation of their growth potential. The results showed that the light saturation point for photosynthesis was relatively high in all the 6 species, which was favourable for the trees to adapt to intense light. However, the apparent quantum yield, photosynthetic rate and light use efficiency (LUE) were significantly lower in T. chinensis, P. Thunbergii and J. chinensis cv kaizuka than in F. chinensis, P. langfanggensis and R. pseudoacacia, with the J. chinensis cv kaizuka showing the lowest efficiency. In addition, the photosynthetic nitrogen-use efficiency and carbon use efficiency were also significantly lower in T. chinensis, P. Thunbergii and J. chinensis cv kaizuka than in F. chinensis, P. langfanggensis and R. pseudoacacia, but the water use efficiency (WUE) was apparently higher in the former 3 species. In terms of the mineral use efficiency (MUE), T. chinensis was at the top and P. thunbergii was at the bottom. The comprehensive index of resource use efficiency, in terms of the utilization of various resources and the photosynthetic performance indicators, could be listed in the following order from high to low: R. pseudoacacia and F. chinensis, P. langfanggensis and T. chinensis, P. thunbergii, J. chinensis cv kaizuka.
Keywords:Photosynthetic performance; resource use efficiency; coast-protective forest; growth potential; tree species
作者简介:刘玉静(1971-),女,硕士,研究方向:生物学. E-mail:chunxiao6@163.com*通讯作者: Author for correspondence. E-mail:qingmeibu@163.com
基金项目:烟台市科技发展计划项目(2012124)
收稿日期:2015-01-06修回日期: 2015-03-02
中图法分类号:S718.43; Q945.79
文献标识码:A
文章编号:1000-2324(2015)04-0570-6
猜你喜欢
风景园林(2022年1期)2022-03-07
林业科技(2020年3期)2021-01-21
意林·少年版(2020年17期)2020-10-12
发明与创新·中学生(2020年4期)2020-04-17
发明与创新(2020年14期)2020-04-16
农民致富之友(2019年11期)2019-05-23
江西农业(2019年1期)2019-03-05
现代农村科技(2019年4期)2019-01-06
现代园艺(2018年2期)2018-03-15
现代园艺(2017年11期)2017-06-28
