
楸叶泡桐嫁接无性系苗期生长优良品种选择1)
2015-03-07 07:15乔杰王炜炜王保平崔令军周海江伊焕杨代贵
东北林业大学学报 2015年10期
乔杰 王炜炜 王保平 崔令军 周海江 伊焕 杨代贵
(国家林业局泡桐研究开发中心,郑州,450003) (河南省商丘市梁园区林业局) (湖北省荆门市林业局)
责任编辑:张 玉。
楸叶泡桐是典型的北方泡桐种,分布于山东、河南、河北、山西、陕西等省。具有冠型紧凑、冠幅窄、自然接干能力强、材色好、密度高、花纹美观等优良特性,为泡桐属11 种中材质最佳者[1-6];但由于其生长速度较北方主栽的兰考泡桐和诸多杂交无性系相对缓慢。在单纯追求速生的培育目标下,其系统研究一直未能开展,栽培利用严重萎缩,且由于其花药败育[1-2],接受周围栽培毛泡桐花粉所获得的种质皆为种间杂交,不能保持其原有典型特征,资源濒临丧失。随着泡桐装饰和高档家具新产品的开发,对泡桐木材材质要求的提高,对楸叶泡桐优良特性的研究和利用日益迫切,亟需选育出生长表现良好、抗性强的优良无性系和研究制定出高效培育的技术措施。
泡桐无性系苗期生长分析和选择,是选育泡桐优良无性系的关键环节,同时也是制定苗期管理措施的重要依据。针对常规的泡桐埋根育苗方法,采用不同生长方程对不同无性系高、径生长动态进行模拟、差异性分析和选择的研究已有一些报道[7-9],但针对楸叶泡桐无性系及嫁接育苗方法的研究不多,在指标选择和统计分析上也尚需商榷。本文对不同楸叶泡桐无性系嫁接育苗的基部断面积和高生长指标的生长动态,采用Richards 方程进行模拟,对其生长时间、生长速度、生长量等参数进行差异性分析,继而选择相关生长和质量指标对参试无性系予以聚类分析,以期选择出苗期生长和质量良好的优良无性系。
1 试验区概况
试验地位于河南省原阳县蒋庄乡靳屋村西北,地理位置为东经113°34'~113°52',北纬34°53'~35°05',地处黄河故道,有部分沙丘,属暖温带大陆性季风气候区。年平均气温14.4 ℃,1月份平均气温-0.6 ℃,7月份平均气温27.0 ℃,极端最高气温44.4 ℃,极端最低气温-13.9 ℃;年降水量582.9 mm,其中60% 集中在6—8月份,年蒸发量2 034 mm;年平均日照时间2 324.5 h;无霜期224 d。土壤为黄河冲积风沙土,土层深厚,以粉沙粒为主。在0~60 cm 土层,0.01~0.25 mm 粒径平均占89.8%,<0.001 mm 粘粒仅占7.3%,透气性强,保水保肥能力极差;土壤密度1.49 g/cm3,pH 值7.9~8.6。0~60 cm 土层,有机质、全氮、有效磷平均质量分数分别为4.314 g/kg、0.145 g/kg、3.506 mg/kg,养分含量较低。
2 材料与方法
2.1 试验材料
试验材料采自13 个楸叶泡桐优树,其中3 个来自湖北省赤壁市泡桐基因库保存的3 个优树无性系,编号为CHB01、CHB02、CHB03;10 个来自2013年2月至2014年3月在山东省系统开展楸叶泡桐资源调查选择的优树,编号为CSD01、CSD03、CSD04、CSD05、CSD06、CSD07、CSD08、CSD09、CSD11、CSD13;以兰考泡桐C125 无性系为对照(CK)。优树选择的标准:冠型紧凑、冠幅窄、细枝型、分枝角度<70°、自然接干能力强、树龄6 a 以上、无丛枝病。在2014年2月下旬,分别采集其1年生枝条,采用保湿、沙藏等措施保存备用。2014年3月8日,在现有育苗圃地,选择统一品种“9501”无性系留床苗为砧木材料,就地枝接。砧木苗株行距为0.8 m×1.0 m,基径为3~5 cm,砧木高度为1~2 cm。按3 个区组、6 株小区的随机区组试验设计。接穗统一选自穗条中部木质化程度较高部位,保留1 对有效芽,劈接后包套透明塑料袋保温、保湿、防损伤。每日观测发芽情况,及时破膜,待芽长20~30 cm 时选健壮芽定芽;根据劈接口愈合情况及时解缚,并封土覆盖砧木3~5 cm。根据土壤墒情及时灌水,5月中旬和8月中旬分2 次施肥,每株分别穴施尿素100 g、(NH4)2HPO4复合肥150 g。
2.2 研究方法
从2014年3月8日嫁接后开始,至当年10月下旬泡桐生长期间,每隔15~20 d 一次,共连续12次,分别对13 个楸叶泡桐无性系(以下简称楸系)和CK 的苗木高生长量(H)、地径生长量(D0、统一距砧木平面5 cm 以避免基部膨大影响)进行测定,最终测定时加测胸径(D1.3),并计算基部断面积生长(S0)、胸高断面积生长(S1.3)、高径比(α = H/D0)、苗干削度(β=(D0-D1.3)/1.3)。
选用方程(1)分别对13 个楸系和CK 各单株苗木的D0、S0、H 的年生长动态进行模拟[10],采用方程(2)~(11)分别计算其生长节律的速生点(t0)、最大连日生长量(Hmax)、速生期开始时间(t1)和结束时间(t2)、速生期(t)、速生期开始时的生长量(H1)、速生期结束时的生长量(H2)、速生期生长量(Ht)、生长后期生长量(Hf)、速生期平均连日生长量(Htm)、生长期(T)及其平均连日生长量(HTm)等生长参数[11,12]。

式(6)中的Z1、Z2按式(12)计算。

式中:Y 分别代表D0、S0、H 三项生长指标。t 分别为从2014年3月8日嫁接开始算起的时间(按天计)。A、k、m 分别为待定参数,A 为各指标生长曲线的渐近值,即各生长指标的最大值;m 决定生长曲线拐点的位置,反映生长函数的类型;k 起着调节生长时间的作用,此值越大其生长时间越短,与A、m 一起反映生长速度,并起更重要作用。方程(2)~(11)各生长参数,分别由方程(1)求1~3 次导数,并令其为0 求得。
采用单因素方差分析方法和Duncan 多重比较方法(又称SSR 法、新复极差法),分析不同无性系的D0、S0、H 三项指标各生长参数的差异性。
选择生长量、生长时间、生长速率等相关生长参数,采用适当系统聚类方法、数据标准化方法和距离计算方法,对13 个楸系和CK 进行聚类,从中选择出苗期生长表现综合良好的楸叶泡桐无性系。
数据处理和分析,采用Excel-2007 和PASW Statistics 18 软件。
3 结果与分析
3.1 不同泡桐无性系苗木生长曲线的拟合
3.1.1 生长曲线的拟合
D0、S0、H 分别为苗木基径生长、基部断面积生长和高生长的三大指标,由于S0与D0相比更能准确反映不同无性系生长参数的差异性,因此,仅对S0和H 的生长曲线进行拟合和分析。
对13 个楸系和CK 的252 株苗木S0和H 生长曲线拟合的结果表明(见表1):Richards 生长方程拟合的复相关系数最小,分别为0.974 4、0.990 8,均达极显著水平。因此,参试无性系苗期的S0和H年生长节律,均可用Richards 方程准确描述。
3.1.2 生长曲线的拟合参数
A 值既是生长曲线的最大值,又是生长指标的总生长量参数和苗木培育的主要目标。由表1可见:不同无性系苗木S0和H 的A 值,分别变化在8.272 3~17.192 8 cm2、292.436 2~436.350 2 cm;在各无性系内的变异幅度,分别达18.97%~34.07%、9.32%~19.82%;无性系间的差异,均达极显著水平。与CK 的S0的A 值相比,CHB03 楸系高6.69%,CHB02、CSD03 二个楸系分别低8.79%、14.78%,与CK 间的差异,均未达显著水平;其它楸系较CK 低16.35%~48.67%,其间的差异均达显著及以上水平。与CK 的H 的A 值相比,CHB03 楸系低3.61%,其间的差异未达显著水平;其它楸系低15.91%~32.98%,与CK 间的差异均达显著及以上水平。
m 值的大小决定着曲线的形状和拐点的位置。由表1可见:不同无性系S0和H 的m 值,分别变化在0.890 4~0.939 6 和0.938 2~0.977 1;从各无性系内的变异幅度看,仅分别为2.23%~5.21%和0.98%~2.78%,为相对稳定参数,但无性系间的差异均达极显著水平。与CK 的S0的m 相比,CHB03楸系低4.02%,与CK 的差异达显著水平;其它12个楸系分别由低2.53%到高1.28%,与CK 的差异均未达显著水平。与CK 的H 的m 相比,CSD06、CHB02、CSD03、CSD07、CSD08、CHB03 六个楸系低1.54%~3.06%,分别与CK 差异显著或极显著;其它7 个楸系,均与CK 差异不显著。
k 值具有调节生长时间和生长速度的重要作用。不同无性系S0和H 的k 值,分别变化在0.025 1~0.035 4 和0.032 3~0.048 7;在各无性系内的变异幅度,分别为7.34%~23.11%和10.90~29.83%;无性系间的差异,均达极显著水平。与CK 的S0的k 相比,CHB03 楸系低16.05%,其间差异达显著水平;CSD05、CSD09 分别高17.39%、18.40%,与CK的差异分别达显著水平;其它10 个楸系,分别由低0.67%到高10.03%,与CK 的差异均未达显著水平。与CK 的H 的k 相比,CSD03、CSD07、CSD06、CSD08、CHB02、CHB03 六个楸系低15.33%~26.09%,与CK 的差异分别达显著水平;其它7 个楸系较CK 由低12.13%到高11.44%,与CK 的差异分别未达显著水平。
综合不同无性系S0和H 的A、k、m 变化情况表明:CHB03 楸系的S0和H 均具有A 值高、k 值和m值低的特点;CHB02、CSD03 二个楸系的S0具有A值较高、k 和m 相对较低的特点。

表1 不同无性系苗木生长曲线拟合参数的差异性
3.2 不同泡桐无性系苗木生长参数的差异性
3.2.1 生长时间参数的差异性
t1、t2、t0是生长曲线的主要时间节点参数,其早晚决定着t 的长短;根据t1、t2将生长过程划分为3个阶段,t1之前为生长初期,t1和t2之间为生长速期,t2之后为生长后期。对参试无性系S0和H 二指标的t1、t2、t0分析的结果表明:无性系间的差异显著性水平(p 值)在5.43 ×10-11~0.046 之间,除了H指标的t1为差异显著外,其它均为差异极显著。
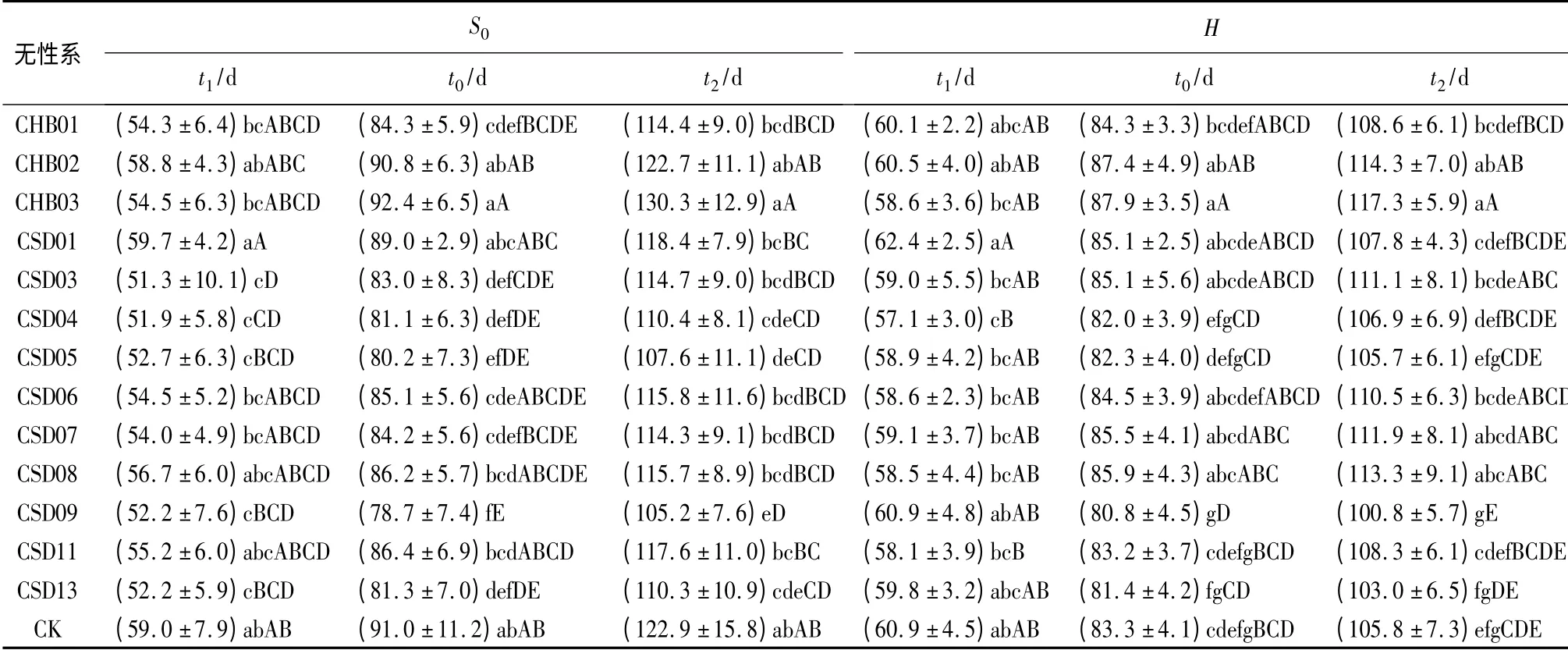
表2 不同无性系苗木t1、t0 和t2 的差异性
由表2中S0指标t1、t2、t0参数的差异性情况可以看出:不同无性系的t1变化,在4月27日—5月6日(嫁接后51~60 d),前后相差9 d;与CK 的t1相比(5月5日),CSD01 延后1 d,CHB02、CSD08、CSD11、CSD06、CHB03、CHB01、CSD07 七个楸系提早0~5 d,均与CK 无显著差异;其它5 个楸系提早6~9 d,均与CK 差异显著或极显著。不同无性系的t2变化,在6月20日—7月15日,前后相差25 d;与CK的t2相比(7月8日),CHB03 楸系延后7 d,CHB02、CSD01、CSD11、CSD06、CSD08、CSD03、CHB01、CSD07 八个无性系分别提早0~9 d,与CK 无显著差异;其它4个无性系提前13~18 d,与CK 差异显著或极显著。不同无性系的t0变化,在5月25日—6月7日,前后相差13 d;与CK 的t0 相比(6月6日),CHB03 楸系延后1 d,CHB02、CSD01、CSD11、CSD08 四个楸系提早0~5 d,其间无显著差异;其它8 个楸系则提前6~12 d,与CK差异显著或极显著。
由表2中H 指标t1、t2、t0参数的差异性情况可以看出:不同无性系的t1变化,在5月3日—5月8日,前后相差5 d,分别较其S0的t1晚2~9 d;与CK 的t1相比(5月7日),除CSD01 延后1 d 外,其它12 个楸系均有所提早,但仅提早1~4 d,且仅有CSD04 楸系提早显著。不同无性系的t2变化,在6月16日—7月2日,前后相差16 d,分别较其S0的t2早2~13 d;与CK 的t2相比(6月21日),CSD05、CSD13、CSD09三个楸系提早0~5 d 外,其它10 个楸系均有所延后,延后时间为1~11 d,其中CSD07、CSD08、CHB02、CHB03 四个楸系延后6~11 d,与CK 差异显著。不同无性系的t0变化,在5月27日—6月3日,前后相差7 d,分别较其S0的t0提早或延后,延后最多2 d,而提早最多8 d;与CK 的t0相比(5月29日),CHB03、CHB02 二个楸系分别延后4、5 d,与CK 差异显著,而其它11 个楸系则与CK 无显著差异。
由于t1、t2、t0的上述差异性,致使不同无性系的t 产生相应的差异。对参试无性系S0和H 二指标的t 的分析结果表明:无性系间的差异显著性水平(p 值)分别达8.27 ×10-7和3.82 ×10-11,均达极显著差异水平。由表3可见:不同无性系S0指标的t变化,在53~76 d,在各无性系内的变异幅度达6.79%~20.39%;与CK 的t 相比(64 d),除CHB03楸系延长12 d 和CSD09 楸系短11 d,与CK 差异显著外,其它11 个楸系均与CK 无显著差异。不同无性系H 指标的t 变化,在40~59 d,分别较其S0指标的t 短4~19 d,在各无性系内的变异幅度达10.42%~20.92%;与CK的t 相比(45 d),CSD06、CSD03、CSD07、CHB02、CSD08、CHB03 六个楸系长7~14 d,与CK 差异显著;其它7 个楸系与CK 差异不显著。
由于无性系苗木S0和H 的T 与t 的比值,分别平均为2.074 4、2.075 5,仅受m 取值范围(0.796 3~0.997 3)的影响,分别变化在2.073 4~2.076 9和2.073 5~2.077 9,变异系数仅分别为0.038%和0.046%,因此,不同无性系的T 变化趋势与其t 的变化趋势高度一致。对参试无性系S0和H二指标的T 的分析结果表明:无性系间的差异显著性p 值分别达7.96 ×10-7和3.81 ×10-11,均达极显著差异。由表3可见:不同无性系S0指标的T 变化,在110~157 d,在各无性系内的变异幅度达6.77%~20.39%;与CK 的t 相比(133 d),除CHB03楸系延长24 d 和CSD09 楸系短23 d,与CK 差异显著外,其它11 个楸系均与CK 无显著差异。不同无性系H 指标的T 变化,在83~122 d,分别较其S0指标的t 短9~39 d,在各无性系内的变异幅度达10.40%~20.88%;与CK 的T 相比(93 d),CSD06、CSD03、CSD07、CHB02、CSD08、CHB03 六个楸系长15~29 d,与CK 差异显著;其它7 个楸系与CK 差异不显著。
综合无性系间S0和H 的t1、t2、t0、t、T 的差异性可以看出:CHB03 楸系具有t1偏中、t2和t0最晚、t和T 最长的特点;CHB02 楸系具有t1、t2、t0均偏晚、t和T 偏长的特点。
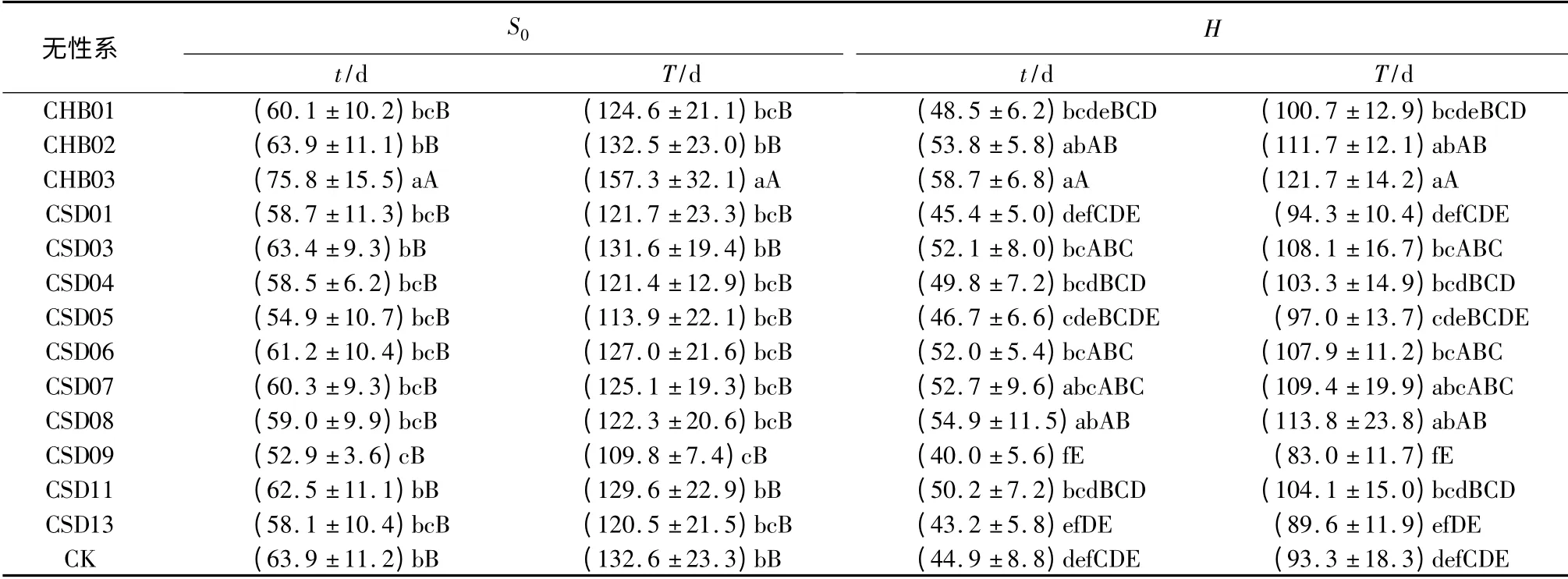
表3 不同无性系苗木t 和T 的差异性
3.2.2 生长速度参数的差异性
Hmax、Htm、HTm为3 个重要的生长速度参数。由于参试无性系苗木S0与H 的Htm分别为其Hmax的85.79%和85.93%(变异系数分别为0.164%和0.086%)、HTm分别为其Hmax的67.93%和67.95%(变异系数分别为0.032%和0.009%),此系列比值仅受m 取值范围的影响变异极小,因此,在无性系间的差异性趋势具有高度一致性。对参试无性系间的差异性分析表明,S0和H 的Hmax、Htm、HTm,在无性系间的差异性均达极显著差异水平。由表4可见:不同无性系苗木S0的Hmax、Htm、HTm均以CHB03、CHB02、CSD06、CSD03 四个楸系较CK 稍有降低,降低幅度为8.88%~13.84%,均与CK 无显著差异;其它9 个楸系则较CK 降低显著或极显著。不同无性系H 的Hmax、Htm、HTm均以13 个楸系显著或极显著低于CK,降低幅度为16.60%~44.58%,这主要与CK 的H 生长量最高、t 和T 较短,而不同楸系H 生长量较低或t 和T 较长密切相关。

表4 不同无性系苗木生长速度参数的差异性
3.2.3 生长量参数的差异性
生长量参数包括3 个生长时期的生长量(H1、Ht、Hf)和最大生长量(A),尤以A 和Ht更为重要,无性系间A 的差异情况如前所述,因此仅对Ht予以分析。由于参试无性系苗木S0和H 的Ht分别为其A 的60.88%、60.93%,受m 取值范围的影响,变异系数仅分别为0.109%、0.034%,因此,Ht与A 二参数在无性系间的差异性趋势也具有高度一致性。由表5可见,不同无性系苗木S0和H 的Ht值,分别变化在5.039~10.452 cm2、178.202~265.906 cm,在各无性系内的变异幅度分别达18.97%~34.00%、9.31%~19.81%,无性系间差异性均达极显著水平。与CK 的S0的Ht值相比,CHB03 楸系高6.51%,CHB02、CSD03 二个楸系分别低8.81%、14.85%,与CK 间的差异均不显著;其它楸系较CK 低16.36%~48.65%,与CK 间的差异均显著或极显著。与CK的H 的Ht值相比,CHB03 楸系低3.66%,其间的差异不显著;其它楸系低15.93%~32.98%,与CK 间的差异均达显著或极显著水平。综合无性系间的A和Ht的差异性可以看出,CHB03 楸系具有S0和H生长量均高的特点,CHB02、CSD03 二个楸系具有S0生长量较高的特点。

表5 不同无性系苗木生长量参数的差异性
3.3 泡桐无性系的苗期选择
时间参数选择t1、t2和T(t1、t2的均值和差数,即为t0和t);生长速度参数选择Hmax、HTm(Hmax、Htm同为速生期生长速度参数,且其变化趋势高度一致);生长量参数选择A、Ht和年末测定的S1.3;苗木质量参数选择年末测定计算出的高径比(α)和苗干削度(β);共选择出17 个生长和质量参数,作为系统聚类的参数。选择采用最远距离系统聚类方法、各指标均值为1 的数据标准化方法和平方欧氏距离计算方法,对13 个楸系和CK 进行系统聚类。聚类结果(见图1)表明:13 个楸系和CK 可分为4 类。第1 类为CHB03 和CK,第2 类为CHB02、CSD03 和CSD06,第3 类为CHB01、CSD01、CSD04、CSD05、CSD07、CSD08、CSD11、CSD13,第4 类为CSD09。由表6可见:第1 类(仅指CHB03),具有速生期和生长期长、生长速度高(S0)或较高(H)、生长量大、高径比和削度适中的特点,为速生类楸系,可作为苗期优良无性系;第2 类3 个楸系,具有速生期和生长期较长、S0生长速度和生长量较大、高径比低的特点,为较速生类,可作为备选的苗期优良无性系;第3 类8 个楸系,具有速生期和生长期较短、生长速度和生长量较低(S0)或低(H)、高径比较低的特点,为慢生类楸系;第4 类楸系,具有速生期和生长期短、S0生长速度低而H 生长速度高、生长量低(S0)或较低(H)、高径比高、削度小的特点,也属慢生类楸系。

图1 不同无性系苗木的生长聚类分析
4 结论与讨论
参试的13 个楸叶泡桐无性系和对照无性系嫁接苗的基部断面积(S0)和高(H)生长过程,均可用Richards 方程准确描述。其A、k、m 三个拟合参数和求解出的生长时间参数(速生期开始时间(t1)、速生期结束时间(t2)、速生点(t0)、速生期(t)、生长期(T))、生长速度参数(最大连日生长量(Hmax)、速生期平均连日生长量(Htm)、生长期平均连日生长量(HTm))、生长量参数(最大生长量(A)、速生期生长量(Ht)),在无性系间的差异均达显著或极显著水平。

表6 4 类楸叶泡桐无性系苗木的主要生长和质量参数
综合不同无性系间S0和H 二指标各参数的差异性分析结果,CHB03 楸系的各参数除了H 生长速度参数显著低于CK 外,其生长量参数和S0生长速度参数均与CK 无显著差异,而其生长时间参数显著优于CK(S0的t 和T 分别长12 d 和24 d、H 生长的t 和T 分别长14 d 和29 d);CHB02 楸系具有t 和T 较长、S0生长量和生长速度较高的特点;CSD03 楸系具有S0生长量和生长速度较高的特点。
选择S0和H 生长的t1、t2、T、Hmax、HTm、A、Ht等和S1.3、高径比和苗干削度等17 个生长和质量参数,对参试无性系进行系统聚类,将13 个楸系分为4 类。其中:第1 类为CHB03 楸系,S0和H 生长量分别达17.19 cm2(地径4.68 cm)、420.58 cm,具有速生期和生长期长、S0生长速度高、生长量大、高径比和削度适中的特点,为速生类楸系,可作为苗期优良无性系;第2 类为CHB02、CSD03 和CSD06 楸系,S0和H 生长量分别达13.97 cm2(地径4.22 cm)、348.10 cm,具有速生期和生长期较长、S0生长速度和生长量较大、高径比低的特点,为较速生类,可作为备选的苗期优良无性系。
采用就地嫁接的方式培育楸系苗木,其S0的t1、t2分别变化在4月27日—5月6日、6月20日—7月15日(其D0的t1、t2分别变化在4月3日—4月15日、5月31日—6月14日),H 的t1、t2分别变化在5月3日—5月8日、6月16日—7月2日,S0、D0和H 的速生期仅分别为53~76、50~71、40~59 d。而常规的泡桐无性系埋根育苗,在同类地区的径、高速生期分别开始于5月下旬—6月上旬、6月中下旬,分别结束于8月上中旬、8月上旬,持续时间分别为61~79、40~50 d[9]。二者相比,前者具有高、径速生期早和径生长速生期短的特点。因此,应根据其速生期调整水肥管理措施。
楸叶泡桐因资源所限,尚无充足的种根用于繁育苗木,采集优树当年生枝条嫁接不失为一种加快资源繁育的有效途径。因其生长速度与本地兰考泡桐和诸多杂交无性系相比,相对缓慢,是否可通过选择优良无性系和利用抗性强(抗旱、抗寒、抗丛枝病)、根系发育好、亲和力强的砧木嫁接,达到提高光合反应能力、促进生长的目标,尚有待进一步研究。
[1] 蒋建平.泡桐栽培学[M].北京:中国林业出版社,1990:32-33.
[2] 李芳东,乔杰,王保平,等.中国泡桐属种质资源图谱[M].北京:中国林业出版社,2013:18.
[3] 常德龙,黄文豪,张云岭,等.4 种泡桐木材材色的差异性[J].东北林业大学学报,2013,41(8):102-104,112.
[4] 张云岭,常德龙,崔俊昌,等.不同种源泡桐木材全干密度差异分析[J].西南林业大学学报,2013,33(4):82-84,88.
[5] 黄文豪,常德龙,唐玉红,等.四种泡桐材抗劈力研究[J].国际木业,2014(1):8-11.
[6] 常德龙,张云岭,胡伟华,等.不同种类泡桐的基本材性[J].东北林业大学学报,2014,42(8):79-81.
[7] 傅大立,杨绍斌,丛培生,等.泡桐苗期年生长参数的分析研究[J].林业科学研究,2001,14(3):301-306.
[8] 魏安智,杨途熙,张晴,等.泡桐无性系苗期生长动态分析研究[J].西北植物学报,2000,20(1):68-78.
[9] 孟伟,杨超伟,夏莘,等.基于Richards 函数的泡桐无性系年生长节律[J].浙江林业科技,2014,34(5):26-31.
[10] 张建国,段爱国.理论生长方程与直径结构模型的研究[M].北京:科学出版社,2004:46-47.
[11] 王保平,李吉跃,文瑞钧,等.修枝接干对泡桐年生长节律影响的研究[J].北京林业大学学报,2003,25(4):11-15.
[12] 李淑花,申初联.第二代杉木人工林生物量的时空特征[J].森林工程,2006,22(1):5-7.
猜你喜欢
南方农业·下旬(2022年4期)2022-05-24
江西农业(2022年8期)2022-05-13
绿色科技(2021年21期)2021-11-26
内蒙古林业调查设计(2021年3期)2021-07-06
现代园艺(2018年1期)2018-03-15
现代园艺(2017年13期)2018-01-19
湖南林业科技(2017年1期)2017-02-06
林业与生态(2016年2期)2016-02-27
安徽林业科技(2015年6期)2015-09-25
吐鲁番(2014年2期)2014-02-28
