
野生硬核种子的表型变异1)
2015-03-07 07:15吴裕张凤良毛常丽曾建生何美莹
东北林业大学学报 2015年8期
吴裕 张凤良 毛常丽 曾建生 何美莹
(云南省热带作物科学研究所,景洪,666100)
责任编辑:潘 华。
硬核(Scleropyrum wallichianum)是檀香科(Santalaceae)硬核属(Scleropyrum)常绿小乔木,我国云南、海南、广西等省区有野生分布,在云南主要分布在勐腊、景洪、勐海、孟连等地海拔700~1 600 m 的山地疏林中[1-3]。初步调查表明,硬核种群数量较大,开花结实正常,自然更新能力较强,种仁含油率高达70%,当地的傣族、基诺族、哈尼族等将硬核种子烤熟后舂碎作香料食用,其香味独特[4-5]。据文献记载,硬核种子油含亚油酸37.11%,亚麻酸37.81%[6],另有文献报道含有炔酸39%,可能有特殊工业用途[7]。因此初步认为硬核有可能被作为云南特色经济植物或功能油料植物开发利用。
天然群体表型变异的研究能使我们认识群体变异式样,是引种驯化和遗传育种的基础,也是揭示其适应性的有效途径之一。种子形态是重要的表型性状,被认为在种内相对稳定,常常作为植物分类的依据,然而由于其所处地理位置、演替阶段及更新方式的不同,植物种在群体间、群体内单株间种子的形态存在很大差异[8-13]。种子的形态作为表型的重要研究内容,对植物分类、发展演替、种群更新、物种保护及遗传育种等都具有重要意义。目前关于硬核种子形态变异的研究文献未见报道,本文选择云南野生分布的硬核群体调查采样,开展种子表型变异研究,探讨群体间和群体内种子变异程度,揭示种子表型变异规律,为硬核资源开发利用提供依据。
1 材料与方法
1.1 群体选择与样品采集
在前期调查18 个样点的基础上[4],根据各样点的水平距离远近和天然隔离屏障等自然因子将16个样点划分为相对独立的7 个野生群体(有2 个样点因植株少而被删除)。其中,景洪市包括2 个群体(基诺P1、勐旺P6),勐腊县包括3 个群体(象明P2、倚邦P3、易武P4),孟连县包括1 个群体(娜允P5),勐海县包括1 个群体(西定P7)。2014年8—9月份,硬核果实成熟时进行采样,选择生长良好的植株,分单株记录,采样时保持株间距离大于50 m,以最大限度地排除近缘单株,同时尽量选择表型差异大的植株,共采集到80 株树的种子。采样过程中用GPS 定位仪对每个群体进行整体和单株定位,采样群体代码及其基本信息列于表1。
1.2 性状选取与定量测定
以烘干种子为材料,对每株树的种子随机抓取30 粒,用游标卡尺测量种子长和种子宽,测量精度为0.01 mm;用电子天平测单粒质量,测量精度为0.001 g;不足30 粒者全部测定。测量数据计算算术平均值和种子长宽比。针对种子质量性状,描述种子形状、种子先端、种子基部、种子刻纹深浅,并进行编码,编码信息见表2。
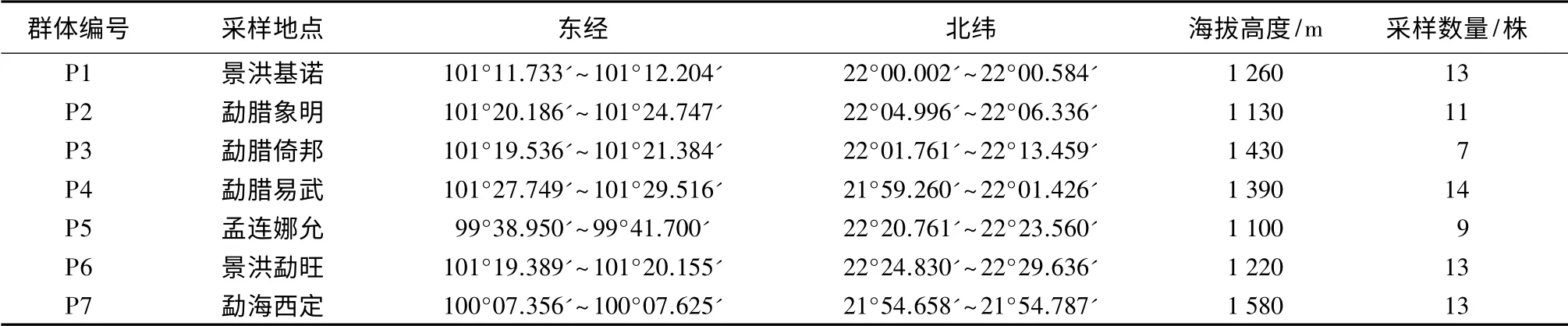
表1 硬核采样群体基本信息

表2 硬核种子质量性状分级代码
1.3 数据处理与分析方法
性状变异特征:以种子长和宽的数据计算种子的长宽比。应用Excel 2003 计算各性状的平均值(X)、标准差(S)、变异系数(CV)、极差(R)、相对极差(RR)。用变异系数表示表型性状的离散程度,用相对极差表示极端差异程度。RR=R/X,其中R=Xmax-Xmin。
方差分析和表型分化系数:用SAS 软件对各数量性状采用巢式设计进行方差分析[10-11],线性模型为:Yijk=u+Si+T(i)j+e(ij)k,式中,Yijk为第i 个群体第j个单株第k 个观测值;u 为总均值;Si为群体效应(固定);T(i)j为群体内单株效应(随机);e(ij)k为试验误差。
遗传多样性指数:利用Shannon-Weaver 遗传多样性指数(Shannon Weaver index of genetic diversity)来衡量群体遗传多样性大小[14]。
统计分析的性状分为两类:第1 类为质量性状,统计各性状类别的频率分布(Pi),用于计算遗传多样性指数(H');第2 类为数量性状,根据平均值、标准差将材料分为10 级,从第1 级X1<(x-2s)到第10 级X10≥(x+2s),每0.5 s 为1 级,每一组的相对频率(Pi)用于计算多样性指数(H');计算公式H'=其中Pij为j 性状在第i 个级别出现的频率[15-16]。各个种群的多样性指数采用Excel 2003软件进行计算。
聚类分析:利用SAS 8.0 软件采用最短距离法对7 个群体进行聚类分析。
2 结果与分析
2.1 群体间和群体内的变异特征及变异来源
群体内各表型性状的变异系数存在差异,从表3可以看出,在所测量性状中,单粒质量的平均变异系数最大(22.07%),种子长(8.66%)略大于种子宽(8.13%)。综合分析各表型性状可以看出,P1、P3 和P6 这3 个群体的变异较大,P5 最小。由群体平均值求得群体间各表型性状的变异系数表明,单粒质量的变异系数最大(8.04%),种子长(3.68%)略大于种子宽(3.12%)。群体间各表型性状的变异模式与群体内的变异模式一致,但各表型性状在群体间的变异系数远远小于群体内的变异系数。
极差是表示最大值与最小值之间的差距,用极差来比较同一性状不同群体的极端变异程度比较直观。但是,由于不同表型性状间量纲不同,用极差进行比较可比性差,故为了消除量纲差异使性状间具有可比性,采用相对极差来表示各群体内的极端变异程度,4 个性状的极端变异按从大到小依次为:单粒质量、种子长、种子长宽比、种子宽。种子表型性状在7 个群体内部的极端变化趋势比较一致,其中P1、P6 变化较大,P5 最小。群体内种子性状相对极差的变化趋势与其变异系数的变化趋势比较吻合,具有较强的规律性。

表3 硬核种子长和宽的群体变异
巢式设计方差分析可以揭示性状在群体内和群体间两个层次的差异性,且可以划分出群体内和群体间的变异组分,从而估量性状变异的主要来源。对种子长、种子宽和单粒质量3 个性状进行巢式方差分析(表4,表5)。分析表明,各表型性状在群体内差异均达到极显著水平(p<0.000 1),在群体间差异均不显著,说明在其群体内存在丰富的变异,群体间变异不丰富;群体间各性状的表型分化系数为4.47%~8.18%,均不到10%,即群体内的平均表型变异占90%以上,说明群体内的变异是云南野生硬核的主要变异来源。巢式方差分析的结果与变异系数的结果一致,综合说明了群体内的多样性远远大于群体间的多样性。

表4 硬核群体种子数量性状方差分析结果
2.2 遗传多样性指数
对硬核种子质量性状(种形、先端、基部、刻纹深浅)及数量性状(种子长、种子宽和单粒质量)进行了遗传多样性指数H'计算(表6)。总体来看,硬核群体内质量性状的遗传多样性指数为0.714~1.484,数量性状的为1.929~2.111,几个数量性状的多样性丰富程度都要高于质量性状。硬核种子的形状多样,但大多为倒卵圆形(59%)和近球形(22%);种子先端突尖大小呈两极分化,大突尖占40%,微突尖占36%;基部形态从圆钝到尖再到锐尖占的比例基本相等,从遗传多样性指数也看出,多样性程度较丰富,具有广泛的变异;刻纹深度大部分为中等类型(76%),变异类型较单一,多样性指数较低。80 株树的种子4 个数量性状从频率分布来看,种子长21.63~26.18 mm 的占77%,种子宽17.19~20.53 mm 的占77%,单粒质量2.80~4.59 g 的占80%,群体内种子长、宽和单粒质量多样性比较丰富。

表5 硬核种子方差分量与表型分化系数
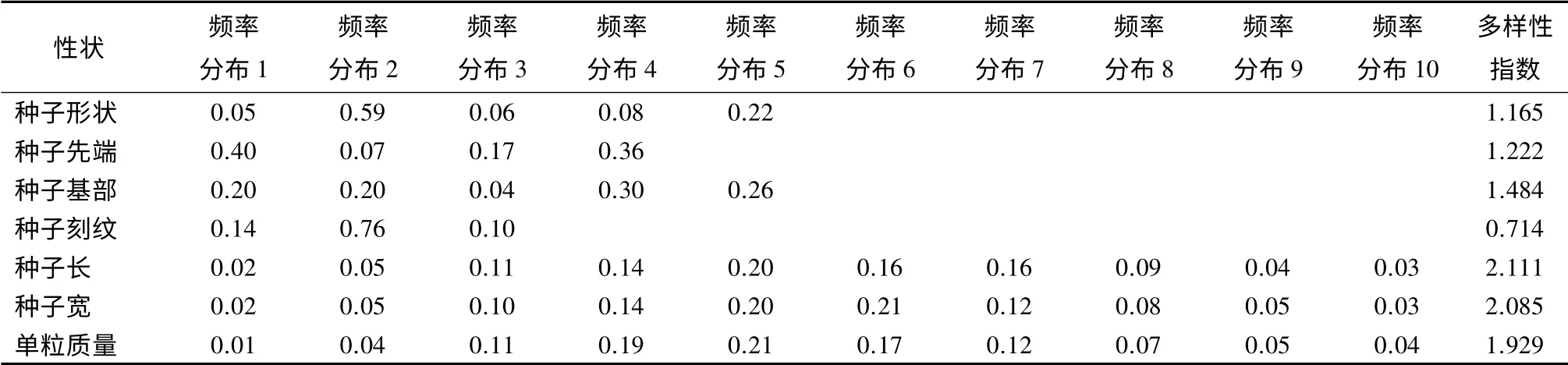
表6 硬核种子性状分布频率和多样性指数
2.3 各表型性状间及其与地理因子间的相关性
各性状及其与地理因子之间的相关性分析表明(表7),几个质量性状间相关性较弱,相关系数为0.025~0.120,受地理因子的影响也较小;几个数量性状间均呈极显著相关性(p<0.01),受环境影响也较大,尤其受海拔高度的影响更明显(p<0.01),种子长、宽和单粒质量随海拔升高而减小,即高海拔地区的种子更小一些。各数量性状及其与质量性状间有较好的相关性,较宽的种子一般基部较圆钝、刻纹较深、种子较长、质量较大;单粒质量极显著地受种子长和宽的影响,尤其与种子宽的相关系数达到了0.901。需要说明的是,海拔1 100 m 的P5 与海拔1 260 m 的P1 和海拔1 390 m 的P4 种子性状相近;P2 和P5 海拔相近,但P2 的种子却明显大得多。可以认为在纬度和海拔相近的条件下,种子大小随经度的变化表现出一定规律,即越往西种子越小,越往东种子越大。但是本研究采样群体较少,尚需进一步研究。

表7 硬核种子各性状及其与地理因子间的相关系数
2.4 数量性状的聚类分析
选取种子长、宽和单粒质量3 个表型性状进行聚类分析,从聚类关系树状图(图1)可以看出,在阈值约为0.75 时可将7 个群体分为3 枝,其中海拔高度在1 100~1 390 m 之间的P1(基诺)、P5(娜允)、P6(勐旺)和P4(易武)聚为一枝,相似程度较高;海拔高度在1 430~1 580 m 之间的P3(倚邦)和P7(西定)聚为一枝,但相似程度相对较低;P2(象明)单独为一枝(海拔1 130 m)。水平距离相近的群体没有很好的聚在一起,而是海拔高度相近的群体聚在一起。需要说明的是,P2 和P5 海拔相近,但是P2 单独分为一枝,P5 却与海拔较高的P1 聚在一起。孟连县与西双版纳州的同纬度同海拔地区相比,热量较低,这可能是导致海拔1 100 m 的P5 与海拔1 260 m 的P1 性状最相近的原因。如果不考虑P5,那么其它6 个群体的表型变异则随海拔高度呈连续性。

图1 硬核7 个群体种子性状聚类关系树状图
3 结论与讨论
据7 个天然群体80 株树的种子形态特征在表型变异系数和表型分化系数方面的分析结果,硬核种子大小和形态存在不同程度的变异。主要表现为群体内的平均表型变异占93.72%,即硬核表型变异的主要来源是群体内。聚类分析表明,水平分布相邻的群体没有很好的聚在一起,而是海拔高度相近的群体聚在一起(P2 除外)。海拔1 100~1 260 m的3 个群体(P1、P5、P6)平均种子长分别为24.79、24.66 和24.29 mm,平均种子宽为19.02、18.94 和18.95 mm,平均单粒质量为3.75、3.80 和3.94 g;海拔1 430~1 580 m 的2 个群体(P3、P7)平均种子长为22.27 和22.37 mm,平均种子宽为18.88 和17.91 mm,平均单粒质量为3.34 和3.17 g;单独聚为一类的P2,平均种子长24.29 mm,平均种子宽为20.28 mm,平均单粒质量为4.33 g。随海拔升高,种子渐小,但是种子形态和种皮刻纹深浅受环境因子影响较小。
表型性状既有变异性又具稳定性,受其本身的遗传组成和所处生态环境等方面的影响,反映了群体遗传稳定性与环境复杂性的互作关系。本研究7个群体3 个数量性状在群体间的变异模式与群体内一致,均表现为CV单粒质量>CV种子长>CV种子宽,在其他树种上也得到类似的变异模式[17-20],说明这几个表型性状的变异趋势相对稳定,为选择育种提供了依据。各性状群体间的变异系数远小于群体内的变异系数,表明群体内多样性更丰富。同一性状在不同群体内变异系数也有一定的差异,但这种差异与水平分布和垂直分布都没有规律。
聚类分析结果表明,7 个群体分为3 组,地理上相邻的群体没有很好的聚在一起,表现出表型性状变异的不连续性。有研究证明,在一定的区域内,海拔高差对种子形态特征构成影响,高差约200 m 的种群,种子性状差异显著[21-22]。本研究中6 个群体分布在景洪、勐腊和勐海(澜沧江流域),北有青藏高原,东有哀牢山脉,阻障寒流入侵,同时受西部沙漠的干暖气流影响,冬季温暖而干燥;夏季,印度洋孟加拉湾和太平洋北部湾暖湿气流顺澜沧江河谷北上,降雨丰富。采样群体东西跨度约150 km,南北跨度约50 km,水平差异导致的生态环境变化不明显,这可能是数量性状变异与经纬度不相关的原因;相比较而言,海拔从1 130 到1 580 m,海拔差异明显影响降水量、气温、云雾等生态因子,海拔差异导致的环境异质性明显影响硬核种子的大小,表现为海拔越高,种子越小。
硬核的花极小,花粉量极少,花色不鲜艳,未发现明显的气味,这些性状都不会被认为具有远距离传粉的可能性,然而群体间变异小,特别是质量性状变异更小,群体内变异却大,可以推测云南野生硬核大群体在很多年以前处于连续分布状态,共享一个基因库,随着地理气候变迁和森林群落的演替,使得硬核种群隔离,成了目前斑块状的分布格局。一般理论认为,遗传基础比性状更具有稳定性,更能反应群体的遗传关系远近,至于硬核这7 个群体间的遗传多样性分布式样尚在研究过程中。
硬核果实和种子的形态特征在单株内保持良好的一致性,其大小也比较均匀。定量分析群体内种子形态多样性的丰富程度表明,数量性状多样性的丰富程度都要高于质量性状。说明野生硬核种子形态特征主要受遗传因素控制,环境因素在一定程度上起到修饰作用。种子大小虽然是一种简单的生物学现象,但包含着丰富的内容,种子大小具有重要的生态学意义,要深入分析影响种子大小的生态因子,尚需设计更精细的研究。
[1] Wu Z Y,Raven P H,Hong D Y.Flora of China[M].BeiJing:Science Press,2003.
[2] 中国科学院中国植物志编辑委员会.中国植物志:24 卷[M].北京:科学出版社,1988.
[3] 中国科学院昆明植物研究所.云南植物志:4 卷[M].北京:科学出版社,1986.
[4] 吴裕,张凤良,何美莹,等.云南油料树种硬核野生分布调查[J].热带农业科技,2015,38(1):24-26.
[5] 何美莹,朱义鑫.西双版纳本土油料植物硬核研究初报[J].热带农业科技,2015,38(1):36-39.
[6] 王惠英,李延辉,李德厚,等.云南热带、亚热带油脂植物研究[C]//中科院西双版纳热带植物园.热带植物研究论文报告集.云南:云南人民出版社,1982.
[7] 王静萍,李京民,于凤兰.硬核油的炔酸成分研究[J].云南植物研究,1992,14(1):101-104.
[8] 李晓洁,徐化成.白皮松种子发芽习性及其种源变异的研究[J].林业科学,1989,25(2):97-105.
[9] 钟章成.植物种群生态适应机理研究[M].北京:科学出版社,2000.
[10] 陈小勇.黄山青冈种子形态变异的初步研究[J].种子,1994(5):16-19.
[11] 操国兴,钟章成,谢德体,等.缙云山川鄂连蕊茶种子形态变异的初步研究[J].西南农业大学学报,2003,25(2):105-107.
[12] 蔡琰琳,金则新.濒危植物夏蜡梅果实、种子形态变异研究[J].西北林学院学报,2008,23(3):44-49.
[13] 唐玲,王艳芳,李荣英,等.滇重楼不同居群的表型多样性研究[J].中国农学通报,2013,29(18):89-95.
[14] Shannon C E,Weaver W.The mathematical theory of communication[M].Urbana:University of Illinois Press,1949.
[15] 赵香娜,李桂英,刘洋,等.国内外甜高梁种质资源主要性状遗传多样性及相关性分析[J].植物遗传资源学报,2008,9(3):302-307.
[16] 董玉琛,曹永生,张学勇,等.中国普通小麦初选核心种质的产生[J].植物遗传资源学报,2003,4(1):1-8.
[17] 李斌,顾万春,卢宝明.白皮松天然居群种实性状表型多样性研究[J].生物多样性,2002,10(2):181-188.
[18] 曾杰,郑海水,甘四明,等.广西西南桦天然居群的表型变异[J].林业科学,2005,41(2):59-65.
[19] 蔡琰琳,金则新.濒危植物夏蜡梅果实、种子形态变异研究[J].西北林学院学报,2008,23(3):44-49.
[20] 孙玉,李庆梅,杨敬元,等.秦岭冷杉球果与种子的形态变异[J].生态学报,2005,25(1):176-181.
[21] 李世超,杨树华,刘海星,等.天山地区弯刺蔷薇居群表型多样性的研究[J].园艺学报,2014,41(8):1723-1730.
[22] 孙玉玲,李庆梅,杨敬元,等.秦岭冷杉球果与种子的形态变异[J].生态学报,2005,25(1):176-181.
猜你喜欢
四川蚕业(2022年1期)2022-06-06
中国食品(2021年21期)2021-11-07
学生天地(2020年17期)2020-08-25
人大建设(2020年2期)2020-07-27
公民与法治(2020年6期)2020-05-30
四川蚕业(2020年4期)2020-02-10
当代陕西(2019年14期)2019-08-26
现代园艺(2017年21期)2018-01-03
中国康复理论与实践(2015年10期)2015-12-24
医学研究杂志(2015年5期)2015-06-10
