
满归地区偃松单木生物量模型1)
2015-03-06 09:17庄会霞刘琪璟孟盛旺贾全全邓留宝
东北林业大学学报 2015年7期
庄会霞 刘琪璟 孟盛旺 贾全全 邓留宝
(北京林业大学,北京,100083) (内蒙古自治区满归林业局)
责任编辑:王广建。
偃松(Pinus pumila)属于松亚科松属五针松组植物,树干通常伏卧地面匍匐生长,树干长度可达3~7 m,但其垂直高度仅1.5~1.8 m,是一种慢生树种[1]。同多种木本植物一样,偃松是一种强适应的多型种[2-3],在适应生境的同时不断改变自己。偃松寿命可达250年,成熟龄为30 a,耐阴性较强,耐火性较差,种子传播距离可达150 m,萌发能力较差[4]。偃松资源分布在欧亚大陆的东部,在我国集中分布在大兴安岭、小兴安岭、张广才岭及长白山的高海拔地带[5];多呈孤岛状分布于高峰顶端,成为森林植被垂直分布的上限,其分布下限的海拔高度自南向北逐渐降低[6]。在高海拔地区,偃松与高山植物(小灌木和草本)混交;在低海拔地区,作为林下植被物种(下木),与高冠植物混交。在大兴安岭地区共有17.7万hm2的偃松矮曲林,另有偃松兴安落叶松林57.8万hm2[7]。大兴安岭的偃松矮曲林主要分布在北部1 000~1 500 m的高海拔处,呈岛状分布,每块的面积10~50 hm2,有的达100 hm2以上。偃松矮曲林所处的位置海拔高,风力大,土壤贫瘠。
国外有关偃松的研究主要涉及群落学特征[8-9]、高生长[10]、生长量[11-14]、光合作用[15-16]、温度适应性[17-19]、种子传播[20]、林龄[21]等方面。我国对偃松的研究主要集中于资源开发利用、形态学、遗传学、引种等方面[22-25],对偃松生物量的研究鲜有报道。
生物量是生态系统生产力的重要体现,对生态系统结构的形成影响巨大[26]。灌木作为生态系统的重要类型之一,在涵养水源、保持水土、生态系统演替等方面具有重要作用。目前,对生物量的研究主要集中在耕地、草地以及森林植被的乔木部分。虽然有一些针对灌木层生物量及其模型的研究[27-28],但主要研究的是阔叶灌木树种[27,29-31]。
偃松作为松类中少有的针叶灌木,或者灌木状小乔木,既具有灌木丛生的特点,又有针叶树种轮生的特性。因此,调查时,既不能完全依照乔木的调查方法,又不能盲目地效仿灌木的调查方法。无论乔木还是灌木,传统生物量研究多采用全株收获称重法,其工作量较大,且耗费时间。本文突破传统研究方法,使用嵌套回归法[32]对偃松单木进行解剖调查,建立偃松各组分各级生物量模型和单木地上各组分生物量模型,为进一步研究偃松生物量提供必要的依据。
1 研究区概况
研究区域位于内蒙古满归林业局,地处大兴安岭北部,隶属于内蒙古根河市。该地区为高纬度寒冷地区,地理坐标为东经121°05'44″~122°47'05″,北纬51°51'02″~52°30'52″,海拔高度1 000~2 000 m。年平均气温为-0.1℃,全年总日照时间为2 467.8 h。最大积雪深度平均为25 cm,全年降水量为504.2 mm。
调查样地位于满归林业局的高地林场的山脊处,海拔1 000 m左右,为偃松单优群落,偃松密度为14 000株/hm2。乔木层伴生少量生长不良的白桦(Betula platyphylla)和兴安落叶松(Larix gmelinii)。林下植被有岩高兰(Empetrum nigrum var.japonicum)、细叶杜香(Ledum palustre)和越桔(Vaccinium vitis-idaea)等。
2 研究方法
2.1 生物量调查
本研究以最小限度干扰的原则,2012年6月份在满归地区高地林场山脊(1 100 m)偃松矮曲林采伐15株偃松。由于偃松以丛生为主,丛内每一个个体视为一个单木。从基部至梢顶,以50 cm为区分段,测定各区分段的带皮直径、皮厚和树干总长度,地上生物量忽略繁殖器官部分。由山脊伐倒木各区分段带皮直径和皮厚,建立去皮直径和带皮直径之间的关系方程。另外,在5株偃松活立木上,也测定了树干各区分段带皮直径,据此可推算相应的去皮直径。利用平均断面积区分求积法,求算树干带皮体积和去皮体积,二者之差即为树皮体积。在树干中部取木材和树皮样品,各取3个重复,采用排水法测其鲜样体积。65℃下烘干至恒质量,计算木材密度和树皮密度。木材(树皮)的生物量=鲜样体积×木材(树皮)密度。
根据嵌套回归法的基本原理[32],对伐倒木的枝条结构进行解析,测定单木所有的一级枝条的基径,选择基径较大的一级枝条测定其上所有的二级枝条基径,依次类推。不分枝条级别测定不同基径枝轴(去掉次级分枝后就是枝轴。)的长度,通过平均断面积区分求积法,计算枝轴体积。选树干中部的枝条,取枝条中部作为样品,采用与树干相同的处理方法,计算枝条密度、体积及生物量。
在偃松伐倒木上,随机选取基径大小不同的枝轴,查数不同枝轴基径所具有的针叶束数。在树干中部的枝条中部取针叶样品,从中随机取50束针叶为一组,3个重复,烘干称质量,求针叶单针干质量。针叶生物量=针叶数量×针叶单针干质量。
2.2 生物量模型建立
根据研究对象的尺度不同,可将生物量模型划分为单木模型和林分模型,本文建立的是单木生物量模型。可采用的单木生物量模型很多[33-35],其中以幂函数模型应用较广。
典型性和乔木型灌木的生物量模型多以地径(D)、离地10 cm处的直径(D10)、高度(H)、冠幅(C)、植冠面积(AC)、植冠体积(VC)及其组合等为自变量[36-37]。但由于偃松株数密度较大,丛与丛之间交错生长,所以相对于低矮的典型性灌木和稀疏的乔木型灌木来说,调查偃松高度、冠幅等比较困难,且准确度很难把握,相反,其基径比较容易调查。因此,本研究以偃松树干基径为自变量,以各器官生物量为因变量,建立二者之间的异速生长方程。模型表达式为W=aDb。式中:W为某组分或全株的生物量;D为枝条或树干带皮基径;a,b为模型参数。
偃松最多具有5级枝条,依据文献报道的嵌套回归法[32]从倒一级枝条(即枝轴)依次拟合各级枝条生物量方程,直至单木枝条总生物量与树干基径之间的回归模型。针叶生物量模型拟合过程与枝条类似,但树干上只有梢顶附近才有针叶生长,所以忽略了树干上的针叶生物量。具体的拟合过程详见嵌套式回归建立树木生物量模型[32]。
分别各分量和总量建立独立生物量模型,但各分量模型间不相容,即分量之和与总量不一致,所以需要进行相容性处理[38-41]。本研究的处理方法是假设W1=f1(x)、W2=f2(x)、W3=f3(x)、W4=f4(x)、W5=f5(x)、W6=f6(x)、W7=f7(x)分别为总量、树干、木材、树皮、树冠、枝条、针叶的独立生物量模型形式。采用三级控制法,以主干生物量为基础,采用两级分配,建立各组分相容性模型。
第一级:W1=f2(x)+f5(x),W2=f2(x),W5=f5(x)。
第二级:W4=f4(x),W3=f2(x)-f4(x)。
第三级,W7=f7(x),W6=f5(x)-f7(x)。
3 结果与分析
3.1 单木各组分生物量模型
3.1.1 树干生物量模型
由于木材生物量和树皮生物量是由其体积和体积密度计算得到,所以活立木的数据也可用于建模。用于生物量建模的单木共15株,基径为5.98~154.45 mm,平均为89.05 mm;树干长度为57~702 cm,平均491 cm。经测定,偃松木材和树皮的密度分别为0.60和0.36 g/cm3。由体积推算的树干部分生物量与树干基径的关系如下。
木材生物量:

树皮生物量:

树干生物量:
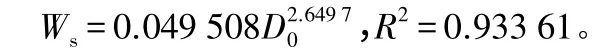
式中:W指生物量;D0指树干基径。
3.1.2 枝条生物量模型
所调查枝轴数量共91条,枝轴基径为3.75~85 mm。倒一级枝条(也可称为枝轴)共372条,基径0.76~5.6 mm。用于建模的倒二级枝条共426根,基径0.76~16.75 mm,对基径0~9 mm的枝条,取每mm的中间两组数据对,其他数据舍去,最终得到25组数据对,基径0.89~16.75 mm。倒三级枝条共46条,基径0.89~31.71 mm。倒四级枝条共54条,基径小于36.01 mm。一级枝条57条,基径小于50.12 mm。由枝轴体积、枝条密度(0.50 g/cm3)和枝轴基径拟合枝轴生物量方程,然后依次拟合倒二级枝条、倒三级枝条直至一级枝条的生物量方程。
倒一级枝条生物量:
Wb5=0.013 972D2.9502,R2=0.963 62。
倒二级枝条生物量:
Wb4=0.013 330D3.0710,R2=0.995 96。
倒三级枝条生物量:
Wb3=0.013 910D3.0642,R2=0.986 25。
倒四级枝条生物量:
Wb2=0.013 651D3.0822,R2=0.980 19。
一级枝条生物量:
Wb1=0.013 865D3.0736,R2=0.989 84。
单木枝条生物量:
Wb=0.001 851D03.1612,R2=0.934 88。
式中:W指生物量;D指对应各级枝条的基径。
3.1.3 针叶生物量模型
基径0.76~21.75 mm的枝轴,其针叶数量为3~27束。共85组数据对(基径,叶束数)。以1 mm为径阶距,统计各径阶叶束数。由此可以得到不同基径的枝轴对应的针叶生物量。各级枝条所具有的针叶生物量的处理方法和枝条生物量一样。依据嵌套回归法拟合各级针叶生物量方程和单木针叶生物量方程。
倒一级针叶生物量:
Wl5=0.398 00D0.99603,R2=0.820 02。
倒二级针叶生物量:
Wl4=0.272 37D1.6996,R2=0.905 78。
倒三级针叶生物量:
Wl3=0.226 15D1.9121,R2=0.903 02。
倒四级针叶生物量:
上述法律人士同时表示,在司法实践中,罚金刑经常会遇到执行难问题,为了解决这一问题,有不少法院会以从轻处罚作为交换条件,促使被告人在正式宣判之前先缴纳罚金,从而解决“判了白判”的问题。所以,开庭前催罚金也就成了不少法院(尤其是基层法院)的普遍做法。
Wl2=0.211 48D1.9708,R2=0.905 46。
一级针叶生物量:
Wl1=0.203 95D1.9931,R2=0.956 61。
单木针叶生物量:
Wl=0.102 93D2.1281,R2=0.991 71。
式中:W指生物量;D为对应各级枝条的基径。
3.2 单木相容性生物量模型
单木各组分生物量与主干基径的相容性模型均为幂函数或者幂函数代数和的形式,决定系数均大于0.80,拟合效果均显著,可以用来估计偃松各组分生物量。
单木地上生物量:
树干生物量:
Ws=0.049 508,R2=0.933 61。
树皮生物量:
Wsb=0.106 32,R2=0.941 16。
木材生物量:
Ww=0.049 508-0.106 32,R2=0.802 84。
树冠生物量:
Wc=0.038 744,R2=0.951 81。
针叶生物量:
Wl=0.102 93,R2=0.991 71。
枝条生物量:
Wb=0.038 744-0.102 93,R2=0.892 05。
式中:W指生物量;D为对应各级枝条的基径;D0为树干的基径。
3.3 偃松单木生物量
随着树干基径的增大,偃松单木各组分和单木地上总生物量随之增大,且木材生物量最高,枝条生物量次之,针叶生物量较低,树皮生物量最低(见表1)。

表1 单木各器官生物量及地上总生物量
单木各组分生物量占地上总生物量的百分比例随树干基径的变化而变化,木材生物量和枝条生物量所占百分比例随树干基径增大而增大,树皮生物量和针叶生物量所占百分比例随之减小,且各组分生物量百分比例随基径的变化程度越来越小(见表2)。树冠生物量约占单木地上生物量的40%,随基径变化不大。作为光合器官的针叶,其生物量占9%~37%。
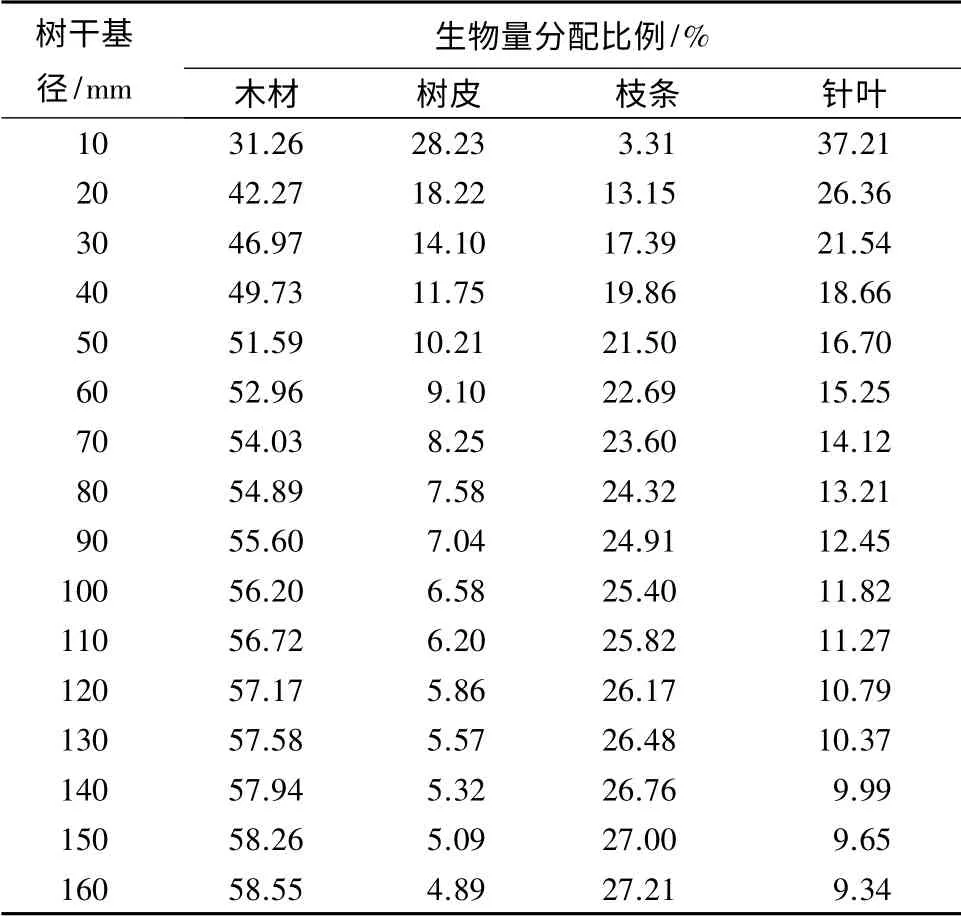
表2 单木生物量分配
4 讨论与结论
由模型结果可以看出,各级枝条生物量模型(除单木枝条生物量模型)的参数a的值为0.013~0.014,参数b的值为2.95~3.10。各级针叶生物量也呈现出相似的现象,验证了嵌套式回归法所建立的模型之间存在紧密的嵌套关系。日本中部偃松单木生物量也是使用异速生长模型进行拟合,但由于调查项目不同,其自变量与本研究不同[12-14]。与各级枝条生物量模型拟合效果相比,针叶各级生物量模型的拟合效果较差,这是因为叶片生物量模拟值往往比较大[42]。由于偃松枝下高非常低,枝条和针叶数量极多,全株实测困难极大,故未对模型进行检验,需要进一步的校正。
虽然用于建立单木模型的偃松单木只有15株,但是每个单木有4~114个一级枝条,某一级枝轴上着生大量的次级枝轴。所以,完全解析一株偃松就可以建立各级枝条和针叶的生物量模型。但是,本文从不同的单木上解析枝条,既保证了数据的代表性,也提供了足够大的建模数据。山西太岳山典型灌木生物量[29]、千烟洲灌木生物量[36]、长白山区林下主要灌木生物量[43]以及东北天然次生林下木树种生物量[37]建模样本为9~56株,无论是乔木型还是典型灌木,其模型样本数量选择10株的居多。另有研究利用5株样木生物量数据拟合日本黑松(Japanese black Pine)各组分生物量模型[44];利用9株样木生物量数据拟合赤松(Pinus densiflora)各组分生物量模型[45];使用10株样木的生物量,拟合挪威云杉(Norway spruce)各组分和单木地上生物量模型[46];利用18株样木生物量数据,拟合西伯利亚落叶松(Larix sibirica)的生物量模型[47]。日本中部偃松单木生物量模型的样本大小为5、10或20株,拟合效果较好[12-14]。本研究建模样本较小,一方面受调查条件的限制,另一方面为了尽可能地减小调查对偃松群落的干扰,但不影响模型拟合的效果。
相关研究结果显示,基径21~44 mm时,偃松单木地上生物量为0.2~1.6 kg[11],基径80~100 mm时,偃松单木地上生物量为10~24 kg[13],满归林业局对应的单木地上生物量分别为0.26~1.82和8.74~15.71 kg,与前者接近,但低于后者。针叶树种单木地上部分生物量各器官生物量分配比例基本表现为:木材>树枝>树皮>针叶,但偃松单木地上生物量分配中,针叶生物量分配比例大于树皮。在大兴安岭北部,兴安落叶松天然林单木木材生物量占地上生物量的60.37%~76.80%,所占比例随年龄增加先增加再减小;树皮生物量占7.15%~20.11%,所占比例随年龄增加而减小;树枝生物量占8.51%~14.29%,所占比例随年龄增加基本呈现增加趋势;树叶生物量占5.12%~7.09%,在低林龄期时所占比例最大[48]。在蒙古国的西伯利亚落叶松单木地上生物量分配到针叶中的仅有5%[47]。满归林业局偃松单木各组分生物量分配及其随年龄的变化,与以上研究存在差异,其单木生物量分配到枝条和针叶的比例较高,可能与树种、立地条件和林分起源等因子有关。在爱尔兰,由于立地条件等因素导致19年生北美云杉地上生物量分配到树叶的比例较高[49]。树干基径为8~10 cm时,日本中部偃松单木地上生物量,树干部分占66.00%~75.68%,枝条占15.32%~22.00%,针叶占9.01%~12.00%[13]。本研究基径8~10 cm的偃松单木地上生物量分配到枝条和针叶的较高,可能是因为林分密度、林龄、立地条件等不同所致。如在日本Tateyama地区,基径1.54~1.83 cm,株数密度36~88株/m2的偃松林,其平均单木针叶生物量占单木地上总生物量的29%~34%[11]。
本研究采用嵌套式回归,利用异速生长模型对偃松伐倒木和活立木进行生物量调查。嵌套式回归的测定样本小,且快速实用,预测值和实测值一致性较高[32]。由于偃松枝下高非常低,枝条和针叶数量极多,全株实测困难极大,故未对模型进行检验,需进一步的验证。为了减小对偃松这种高寒植被的干扰,本研究采样样本较小,且树种生长条件具有地域差异。因此,本研究所建立的模型适用于相似地块的偃松生物量的预估,实际应用时需要进一步的校正。
[1]Sano Y,Matano T,Ujihara A.Growth of Pinus pumila and climate fluctuation in Japan[J].Nature,1977,266:159-161.
[2]Ishii S.On the various forms of Pinus pumila distinguished by the structure of leaves with special reference to their distribution[J].Japanese Journal of Forestry Research,1938,20(6):1-19.
[3]张翰杰.中国松属一新种[J].植物研究,1985,5(1):151-156.
[4]徐化成.中国大兴安岭森林[M].北京:科学出版社,1998.
[5]林春芳,崔克诚.偃松资源开发利用[J].林业实用技术,2002(1):41-43.
[6]《新疆森林》编委编著.新疆森林[M].乌鲁木齐:新疆人民出版社,1989:117-121.
[7]姜孟霞.对大兴安岭偃松林分布、生长的初步研究[J].林业科学,1982,18(2):203-205.
[8]Kobayashi K.Phytosociological studies on the scrub of dwarf pine(Pinus pumila)in Japan[J].Japan Science,1971,14:1-52.
[9]Araki M,Naoki T,Araki M,et al.Community structure and growth habit of siberian dwarf pine(Pinus pumila)[J].Japanese Forestry Journal,1993,75(5):463-468.
[10]Takahashi K,Yoshida S.How the scrub height of dwarf pine Pinus pumila decreases at the treeline[J].Ecological Research,2009,24(4):847-854.
[11]Shidei T.Productivity of haimatsu(Pinus pumila)community growing in alpine zone of Tateyama-Range[J].Japanese Forestry Journal,1963,45(6):169-173.
[12]Kajimoto T.Aboveground biomass and litterfall of Pinus pumila scrubs growing on the Kiso Mountain Range in Central Japan[J].Ecological Research,1989,4(1):55-69.
[13]Kajimoto T.Dynamics and dry matter production of belowground woody organs of Pinus pumila trees growing on the Kiso mountain range in central Japan[J].Ecological Research,1992,7(3):333-339.
[14]Kajimoto T.Aboveground net production and dry matter allocation of Pinus pumila forests in the Kiso mountain range,central Japan[J].Ecological Research,1994,9(2):193-204.
[15]Kajimoto T.Photosynthesis and Respiration of Pinus pumila needles in relation to needle age and season[J].Ecological Research,1990,5(3):333-340.
[16]Ishida A,Nakano T,Sekikawa S,et al.Diurnal changes in needle gas exchange in alpine Pinus pumila during snow-melting and summer seasons[J].Ecological Research,2001,16(1):107-116.
[17]Ohmori H,Yanagimachi O.Thermal conditions of the lower limit of Pinus pumila zone and of the upper and lower limits of betula ermanii zone in Hokkaido,Japan[J].The Geographical Journal,1989,98(1):1-18.
[18]Okitsu S.Consideration on temperature condition of the lower limit of a Pinus pumila zone:a comment on ohmori and yanagimachi[J].The Geographical Journal,1989,98(2):1-18.
[19]Takahashi N,Sato K.Summer ground temperature conditions in the Pinus pumila community in the alpine zone of the Daisetsuzan Mountains[J].Geographical Review of Japan,1996,69(8):693-705.
[20]Hayashida M.Seed dispersal of Japanese stone pine by the Eurasian Nutcracker[J].Ornithological Science,2003,2:33-40.
[21]Okitsu S.Age estimation of above-ground parts of Pinus pumila REGEL stands in Japan[J].Japanese Journal of Forestry Research,1987,69(5):195-197.
[22]孙祝宾.林木的季节生长[J].内蒙古林业科技.1980(3):34.
[23]陈国富,韦昌雷,朱万昌,等.偃松及其价值[J].特种经济动植物,2000(1):28-32.
[24]尹君,曹海波,崔克城,等.偃松的观赏药用价值及加工利用[J].特种经济动植物,2002(2):29.
[25]范瑞红,栾连航,倪薇,等.偃松引种研究[J].林业科技,2009,(6):5-7.
[26]宇万太,于永强.植物地下生物量研究进展[J].应用生态学报.2001,12(16):927-932.
[27]曾慧卿,刘琪璟,冯宗炜,等.红壤丘陵区林下灌木生物量估算模型的建立及其应用[J].应用生态学报,2007,18(10):2185-2190.
[28]杨昆,管东生.森林林下植被生物量收获的样方选择和模型[J].生态学报,2007,27(2):705-714.
[29]陈遐林,马钦彦,陈宗伟,等.山西太岳山典型灌木林生物量及生产力研究[J].林业科学研究,2002,15(3):304-309.
[30]黎燕琼,郑绍伟,龚固堂,等.不同年龄柏木混交林下主要灌木黄荆生物量及分配格局[J].生态学报,2010,30(11):2809-2818.
[31]黄劲松,邸雪颖.帽儿山地区6种灌木地上生物量估算模型[J].东北林业大学学报,2011,39(5):54-57.
[32]刘琪璟.嵌套式回归建立树木生物量模型[J].植物生态学报,2009,33(2):331-337.
[33]冯宗炜,陈楚莹,张家武,等.湖南会同地区马尾松林生物量的测定[J].林业科学,1982,18(2):127-134.
[34]刘兴良,刘世荣,宿以明,等.巴郎山川滇高山栎灌丛地上生物量及其对海拔梯度的响应[J].林业科学,2006,42(2):1-7.
[35]吕晓涛,唐建维,何有才,等.西双版纳热带季节雨林的生物量及其分配特征[J].植物生态学报,2007,31(1):11-22.
[36]曾慧卿,刘琪璟,马泽清,等.千烟洲灌木生物量模型研究[J].浙江林业科技,2006,26(1):13-17.
[37]李晓娜,国庆喜,王兴昌,等.东北天然次生林下木树种生物量的相对生长[J].林业科学,2010,46(8):22-32.
[38]张会儒,赵有贤,王学力,等.应用线性联立方程组方法建立相容性生物量模型研究[J].林业资源管理,1999(6):63-67.
[39]邢艳秋,王立海.基于森林调查数据的长白山天然林森林生物量相容性模型[J].应用生态学报,2007,18(1):1-8.
[40]程堂仁,冯菁,马钦彦,等.小陇山油松林乔木层生物量相容性线性模型[J].生态学杂志,2008,27(3):317-322.
[41]曾伟生,唐守正.东北落叶松和南方马尾松地下生物量模型研建[J].北京林业大学学报,2011,33(2):1-6.
[42]Wang C.Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests[J].Forest Ecology and Management,2006,222(1/3):9-16.
[43]何列艳,亢新刚,范小莉,等.长白山区林下主要灌木生物量估算与分析[J].南京林业大学学报:自然科学版,2011,35(5):45-50.
[44]Konôpka B,Tsukahara H,Netsu A.Biomass distribution in 40-year-old trees of japanese black pine[J].JFor Res,2000,5(3):163-168.
[45]Hiromi T,Takashi N,Manami M,et al.Biomass and net primary production of a Pinus densiflora forest established on a lava flow of Mt[J].Fuji in central Japan Journal of Forest Research,2003,8(4):247-252.
[46]Weis W,Gruber A,Huber C,et al.Element concentrations and storage in the aboveground biomass of limed and unlimed Norway spruce trees at Höglwald[J].Eur J Forest Res,2009,128(5):437-445.
[47]Purevragchaa B,Jamsran T,Choimaa D,et al.Equations for estimating the above-ground biomass of Larix sibirica in the foreststeppe of Mongolia[J].Journal of Forestry Research,2013,24(3):431-437.
[48]肖生苓,杨嘉龙.大兴安岭北部兴安落叶松天然林单木地上生物量[J].林业科学,2014,50(8):22-29.
[49]Green C,Tobin B,O’Shea M,et al.Above-and belowground biomass measurements in an unthinned stand of sitka spruce(Picea sitchensis(Bong)Carr.)[J].Eur J Forest Res,2007,126(2):179-188.
猜你喜欢
农业工程学报(2022年14期)2022-10-19
实验室研究与探索(2022年4期)2022-08-06
林业科学(2020年10期)2020-11-30
四川林业科技(2019年5期)2019-07-07
作文周刊·小学四年级版(2019年8期)2019-04-23
遥感信息(2019年1期)2019-03-22
安徽农学通报(2019年1期)2019-02-14
湖北林业科技(2019年6期)2019-01-06
森林工程(2018年5期)2018-05-14
湖北林业科技(2017年2期)2017-05-25
