
飞播马尾松纯林补植木荷后土壤酶活性与土壤养分变化1)
2015-03-06 09:17林惠章郭孝玉欧阳勋志吴自荣
东北林业大学学报 2015年7期
林惠章 郭孝玉 欧阳勋志 吴自荣
(江西农业大学,南昌,330045)
责任编辑:程 红。
土壤酶对森林生态系统物质循环、能量转换以及土壤肥力的形成起着重要作用,是评价土壤质量的重要生物学指标之一[1]。长期以来,关于土壤酶与土壤养分之间的关系倍受学者们的关注[2-5]。作为土壤环境的重要组成部分,土壤酶及土壤养分对林分植被有着重要的作用[6],同时,林型[7]及经营措施[8-9]的差异反过来也影响着土壤酶活性的发挥。
马尾松(Pinus massoniana)是我国亚热带森林主要针叶树种,具有耐干旱瘠薄,适应性强等特点,也是亚热带地区主要的飞播造林树种。不少学者对马尾松林的土壤养分[10-11]、土壤酶活性[12]及其二者关系[13]进行了研究,如葛晓改等[13]发现马尾松林土壤养分与酶活性之间关系密切;林德喜等[11]研究得出马尾松林下补植闽粤栲(Castanopsis fissa)、青栲(Cyclobalanopsis myrsinaefolia)和苦槠(Castanopsis sclerophylla)等阔叶树能显著提高养分周转率,改善土壤养分状况;相关研究表明,马尾松纯林补种阔叶树,能有效促进其进展演替,是提高土壤养分和改善生态环境的有效形式[14-15]。然而,树种组成不同凋落物及根系不同,都将直接或间接影响土壤养分含量与土壤酶的活性[16-17]。
赣南曾是我国水土流失较为严重的地区,为改善生态环境,减少水土流失,自1965年开展马尾松飞播造林工作,并取得显著效果,但单一针叶纯林的弱点也逐渐暴露出来。20世纪90年代初,为优化森林结构,对部分纯林补植木荷(Schima superba)形成了马尾松—木荷混交林,而这种混交林的土壤酶活性及土壤养分与纯林有何不同目前尚未见相关报道。因此,以赣南飞播马尾松种子造林及补植木荷具有代表性的兴国县为研究区,探讨飞播马尾松纯林、马尾松—木荷混交林2种林型土壤酶活性和土壤养分的关系,以期了解补植木荷对土壤酶活性和土壤养分的影响,为飞播马尾松林的经营管理提供科学依据。
1 研究区概况
研究区位于赣南的兴国县(26°03'~26°42'N,115°01'~115°51'E)境内,地处武夷山支脉雩山地段,以低山、丘陵为主,属亚热带温暖湿润气候,年平均气温18.9℃,年平降水量1 539 mm,海拔130~1 200 m,境内土壤以红壤为主,森林类型主要为马尾松林、杉木(Cunninghamia lanceolata)林、针阔混交林、常绿阔叶林等。据兴国县林业局相关统计资料,至2001年,兴国县马尾松飞播造林面积达77 633.3 hm2,其中保存面积64 000 hm2,占全县现有林地面积的29.5%[18],其林下植被主要有檵木(Loropetalum chinense)、铁芒萁(Dicranopteris linearis)、五节芒(Miscanthus floridus)等。上世纪90年代初,对部分飞播马尾松纯林进行间伐、补植木荷的试验,形成了马尾松—木荷混交林。
2 材料与方法
2.1 样地布设
2012年7月份在对飞播马尾松纯林、马尾松—木荷混交林典型分布区(中丘地貌,海拔100~300 m)踏查的基础上,选取林分设置典型样地,每种类型3个重复,样地大小为30 m×30 m,调查记录各样地林分胸径、林龄、郁闭度及林下植被等因子。马尾松—木荷混交林林下植被主要有檵木、美丽胡枝子(Lespedeza formosa)、铁芒萁、五节芒。马尾松纯林林下植被主要有檵木、铁芒萁、五节芒。2种林型样地概况见表1。

表1 飞播马尾松纯林及马尾松—木荷混交林样地基本概况
2.2 土样采集
在每块典型样地的上、中、下部采用土钻法(土钻直径5 cm)分0~10、>10~20、>20~40 cm 3个土层采集土壤样品,每个层次各取3钻。将各层土壤样品混合并均分为2份:其中一份样品去掉石砾、动植物根系及残体,混均过筛后装入保鲜袋低温处理(置于冰箱中3~4℃),用于测试土壤酶活性;另一份样品经风干、磨细、过筛后装入保鲜袋用以测定土壤养分。
2.3 土壤酶活性与养分测定
土壤酶及土壤养分测定方法为[18-19]:脲酶采用苯酚钠—次氯酸钠比色法(靛酚蓝比色法);蔗糖酶采用3,5—二硝基水杨酸比色法;酸性磷酸酶采用对硝基苯磷酸二钠比色法;全氮采用硫酸消化—凯氏定氮法;速效氮采用碱解扩散法;全磷采用酸溶—钼锑抗比色法;速效磷采用氟化铵盐酸比色法;全钾采用氢氧化钠熔融—火焰光度法;速效钾采用醋酸铵提取—火焰光度法;土壤有机质采用重铬酸钾硫酸氧化—硫酸亚铁滴定法。
2.4 数据处理
使用SPSS 17.0软件进行数据处理和分析。采用单因素方差分析(One-way ANOVA)分析同一林型不同土层间土壤养分及酶活性的差异显著性;采用独立样本T检验分析同一土层土壤养分与酶活性在不同林型间的差异性;采用Pearson相关分析土壤酶与土壤养分之间的相关关系。
3 结果与分析
3.1 土壤酶活性
2种林型不同土层土壤酶活性变化规律(表2)显示,同一土层土壤脲酶、蔗糖酶活性均表现为混交林大于纯林,而酸性磷酸酶表现为纯林大于混交林。混交林0~10、>10~20、>20~40 cm土层土壤脲酶依次为0.199、0.159、0.125 mg·g-1,蔗糖酶为17.036、15.326、8.108 mg·g-1,酸性磷酸酶分别为0.151、0.085、0.070 mg·g-1;纯林0~10、>10~20、>20~40 cm土层土壤脲酶分别为0.132、0.126、0.112 mg·g-1,蔗糖酶为11.281、4.563、2.935 mg·g-1,酸性磷酸酶为0.173、0.130、0.109 mg·g-1。可以看出,2种林型土壤脲酶、蔗糖酶及酸性磷酸酶的活性均随土层的加深而降低。单因素方差分析表明:不同林型同一土层蔗糖酶差异显著(P<0.05);而脲酶在0~10 cm土层差异显著(P<0.05),>10~20、>20~40 cm土层差异不显著(P>0.05);酸性磷酸酶在0~10 cm土层差异不显著(P>0.05),>10~20、>20~40 cm土层差异显著(P<0.05)。

表2 飞播马尾松纯林及马尾松—木荷混交林土壤酶活性
3.2 土壤养分
由表3可知,不同林型同一土层同一养分含量均表现为混交林大于纯林,且各土层全氮、速效钾差异显著(P<0.05),全钾、速效氮、速效磷差异不显著(P>0.05),而全磷、有机质在0~10、>10~20 cm土层差异显著(P<0.05)、>20~40 cm土层差异不显著(P>0.05)。同一林型不同土层土壤养分含量均随土层加深而降低,且全氮、全磷、全钾、速效磷、速效钾土层间差异不显著(P>0.05),>10~20 cm土层的速效氮、有机质质量分数显著低于0~10 cm土层(P<0.05),与>20~40 cm土层差异不显著(P>0.05)。

表3 飞播马尾松纯林及马尾松—木荷混交林土壤养分质量分数
3.3 土壤酶活性与土壤养分的相关性
2种林型土壤酶活性与土壤养分的Pearson相关分析表明,脲酶和蔗糖酶均与全氮、全磷、速效氮、速效钾、有机质呈极显著正相关(P<0.01),与全钾呈显著正相关(P<0.05),而速效磷与蔗糖酶呈极显著正相关(P<0.01),与脲酶相关性不显著(P>0.05);酸性磷酸酶与全磷、速效磷、有机质呈极显著正相关(P<0.01),与速效氮呈显著正相关(P<0.05),与全氮、全钾、速效钾无显著相关性(P>0.05)(表4)。
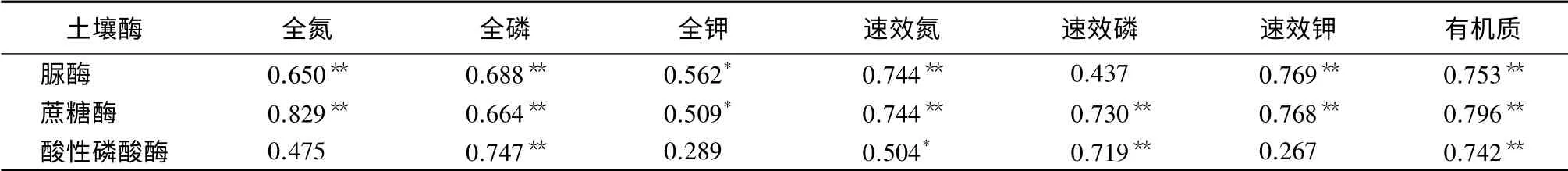
表4 土壤酶活性与土壤养分间的相关性
4 结论与讨论
通过对比发现,同一土层脲酶、蔗糖酶活性与土壤养分均表现为混交林大于纯林,而酸性磷酸酶却相反;2种林型同一土层蔗糖酶、全氮、速效钾差异显著(P<0.05),脲酶在0~10 cm土层差异显著(P<0.05),酸性磷酸酶则在>10~20、>20~40 cm土层差异显著(P<0.05),全磷、有机质在0~10、>10~20 cm土层差异显著(P<0.05)。混交林土壤脲酶、蔗糖酶活性及土壤养分高于纯林的主要原因可能是纯林凋落物较单一,松针分解速率慢,养分归还周期较长[10],从而影响土壤酶的活性[20];补植木荷后,增加了主林层的生物多样性以及凋落物种类及数量,改善了林分结构和凋落物组成,加快了凋落物的分解,增加了微生物活性,从而促进部分酶的活性[21];而纯林土壤酸性磷酸酶活性高于混交林这一特殊情况,与胡亚林等[22]的研究结果有着相似之处,说明土壤酶活性不仅与凋落物种类及树种组成有关,而且还受酶的来源、生物学特性以及微地形变化的影响[23];整体而言,马尾松—木荷混交林土壤质量及土壤环境优于马尾松纯林,与戴凌等[16]研究结果相一致。本研究发现,同一林型土壤酶活性及土壤养分均表现为随着土层深度的增加而降低,主要是因为土壤表层累积较多的凋落物及其光照、水分及通透性较好,能有效促进凋落物的分解,增加土壤养分含量,为土壤酶促反应提供较多的底物[24],随着土层加深,通气状况下降,土壤养分含量减少,土壤酶活性也呈下降趋势,这与其他学者的研究结果相一致[25-26],表明酶活性在一定程度上能够表征土壤养分状况。相关性分析表明,土壤酶活性与土壤养分存在一定的相关性,与周静等[27-29]的研究结果基本一致。3种土壤酶活性均与有机质、全磷、速效氮存在显著或极显著正相关关系(P<0.05),表明有机质和N、P可能是影响土壤酶活性的重要因子;而酸性磷酸酶与全磷、速效磷呈极显著正相关(P<0.05),与全氮、全钾、速效钾相关性不显著(P>0.05),可知土壤中P元素可能是影响酸性磷酸酶活性的主要因子。
综上所述,研究区土壤脲酶、蔗糖酶、酸性磷酸酶等3种土壤重要酶与土壤养分间有着显著相关关系,在马尾松纯林中补植木荷有利于提高土壤养分含量及脲酶、蔗糖酶的活性,因此在今后对飞播马尾松林的经营管理中可考虑适当补植木荷,以达到提高林地土壤质量,加快群落演替进程的目的。
[1]Verstraete W,Voets JP.Soil microbial and biochemical characteristicsin relation to soil management and fertility[J].Soil Biologyand Biochemistry,1977,9(4):253-258.
[2]Badiane N N Y,Chotte JL,Pate E,et al.Use of soil enzyme activities to monitor soil quality in natural and improved fallows in semi-arid tropical regions[J].Applied Soil Ecology,2001,18(3):229-238.
[3]Singh K P,Mandal T N,Tripathi S K.Patterns of restoration of soil physicochemical properties and microbial biomassin different landslide sites in the sal fores t ecosystem of Nepal Himalaya[J].Ecological Engineering,2001,17(4):385-401.
[4]郭春兰.油茶林土壤微生物量氮和酶活性的时空变异与影响因素[J].江西农业大学学报,2014,36(5):940-947.
[5]张日升,贾树海,徐贵军,等.营林措施对沙地樟子松人工林土壤养分、酶活性及微生物量碳的影响[J].土壤学报,2011,42(1):65-68.
[6]陈彩虹,叶道碧.4种人工林土壤酶活性与养分的相关性研究[J].中南林业科技大学学报,2010,30(6):64-68.
[7]司登宇,张金池,陈莉莎,等.浙江省凤阳山不同林分类型土壤酶活性研究[J].水土保持通报,2013,33(6):258-263.
[8]颜欢欢,吴承祯,洪伟,等.千年桐生态林近自然经营对土壤酶活性的影响[J].西南林业大学学报,2014,34(3):1-6.
[9]郝俊鹏,凌宁,李瑞霞,等.间伐对马尾松人工林土壤酶活性的影响[J].南京林业大学学报:自然科学版,2013,37(4):51-56.
[10]马元丹,江洪,余树全,等.不同起源时间的植物叶凋落物在中亚热带的分解特性[J].生态学报,2009,29(10):5237-5345.
[11]林德喜,樊后保.马尾松林下补植阔叶树后森林凋落物量、养分含量及周转时间的变化[J].林业科学,2005,41(6):7-15.
[12]葛晓改,肖文发,曾立雄,等.三峡库区马尾松林土壤-凋落物层酶活性对凋落物分解的影响[J].生态学报,2014,34(9):2228-2237.
[13]葛晓改,肖文发,曾立雄,等.三峡库区不同林龄马尾松土壤养分与酶活性的关系[J].应用生态学报,2012,23(2):445-451.
[14]Brookes P C,Landman A,Pruden G,et al.Chloroform fumigation and the release of soil nitrogen:a rapid direct extraction method to measure microbial biomass nitrogen in soil[J].Soil Biology and Biochemistry,1985,17(6):837-842.
[15]樊后保,李燕燕,苏兵强,等.马尾松-阔叶树混交异龄林生物量与生产力分配格局[J].生态学报,2006,26(8):2463-2473.
[16]戴凌,黄志宏,文丽,等.长沙市不同森林类型土壤养分含量与土壤酶活性[J].中南林业科技大学学报,2014,34(6):100-105.
[17]张鹏,田兴军,何兴兵,等.亚热带森林凋落物层土壤酶活性的季节动态[J].生态环境,2007,16(3):1024-1029.
[18]丁松,应学良,吕丹,等.赣南飞播马尾松林林下植被盖度对土壤质量的影响[J].水土保持研究,2014,21(3):31-36.
[19]关松荫.土壤酶及其研究法[M].北京:中国农业大学出版社,1986.
[20]吴永铃,王兵,戴伟,等.杉木人工林土壤酶活性与土壤性质的通径分析[J].北京林业大学学报,2012,34(2):78-83.
[21]陈莉莉,王得祥,于飞,等.松栎混交林土壤微生物数量与土壤酶活性及土壤养分关系的研究[J].土壤通报,2014,45(1):77-84.
[22]胡亚林,汪思龙,黄宇,等.凋落物化学组成对土壤微生物学性状及土壤酶活性的影响[J].生态学报,2005,25(10):2662-2668.
[23]邱莉萍,张兴昌,程积民.坡向坡位和撂荒地对云雾山草地土壤酶活性的影响[J].草业学报,2007,16(1):87-93.
[24]Kautz T,Lopez-Fando C,Ellmer F.Abundance and biodiversity of soil microarthropods as influenced by different types of organic manure in a long-term field experiment in Central Spain[J].Applied Soil Ecology,2006,33(3):278-285.
[25]范艳春,王鹏程,肖文发,等.三峡库区2类典型森林5种土壤酶季节动态及其与养分的关系[J].华中农业大学学报,2014,33(4):39-44.
[26]罗飞,谢书妮,张海燕,等.酸雨区不同林龄杉木林土壤酶活性季节动态[J].福建林学院学报,2014,34(2):131-137.
[27]方晰,田大伦,秦国宣,等.杉木林采伐迹地连栽和撂荒对林地土壤养分与酶活性的影响[J].林业科学,2009,49(12):65-71.
[28]周静,袁华明,李佳泳,等.杉木-光皮桦混交林对微生物量碳氮与酶活性的影响[J].东北林业大学学报,2015,43(3):83-86,123.
[29]刘云鹏,申卫博,张社奇,等.黄河中游湿地土壤养分与酶活性特征及相关性研究[J].草地学报,2013,21(3):474-478,484.
猜你喜欢
内蒙古林业(2022年8期)2022-09-08
中国林副特产(2022年4期)2022-08-08
散文诗世界(2022年5期)2022-05-23
林业与环境科学(2020年3期)2020-07-21
农民致富之友(2019年7期)2019-05-23
新农民(2019年11期)2019-02-20
草原与草业(2018年3期)2018-10-23
森林工程(2018年4期)2018-08-04
方圆(2018年23期)2018-01-07
花卉(2016年14期)2016-07-31
