
光周期和温度对条纹小斑蛾羽化和交配节律的影响
2015-03-02 03:35何海敏肖海军薛芳森
江西农业大学学报 2015年6期
何海敏,肖 亮,傅 淑,肖海军,薛芳森
(江西农业大学昆虫研究所,江西南昌330045)
光周期和温度对条纹小斑蛾羽化和交配节律的影响
何海敏,肖亮,傅淑,肖海军,薛芳森*
(江西农业大学昆虫研究所,江西南昌330045)
摘要:为了探明条纹小斑蛾Thyrassiapenangae(Moor)羽化及交配行为的发生规律,在实验室不同光周期和温度条件及自然条件下详细调查了该虫的羽化和交配节律。结果表明:在自然条件下,成虫羽化高峰期出现在上午07:00—08:00。在 25、28和30 ℃恒温条件下,羽化高峰期随着温度升高而提前,分别出现在上午08:00—09:00,07:00—08:00和06:00—07:00;成虫羽化时间基本不受光周期的影响。在自然条件下,成虫越冬代至第3代出现的交配时间随季节日长的延长而推迟,交配高峰期分别为15:00—17:00,16:00—18:00,17:00—19:00和18:00—20:00。在25、28、30 ℃恒温条件下,交配时间随日长的延长而推迟,交配高峰出现的时间在日长11 h时为16:00—17:00,日长13 h时为17:00—18:00,日长15 h时为19:00—20:00;交配时间受温度的影响较小。
关键词:条纹小斑蛾;羽化节律;交配节律;光周期;温度
昆虫的发育和某些行为如迁移、取食、孵化、羽化、交配、产卵等对个体而言一生只发生一次,其发生时间受内在的生物钟控制[1]。但这些行为的发生,还是会受到外界环境条件的干扰,如光周期、温度或者两者同时影响[2-5]。在昆虫的诸多行为节律中,人们研究最多的是它们的羽化节律。例如,果蝇Drosophilaspp.[2,6-9]、黄粪蝇Scopeumastercoraria[10]、棉红铃虫Pectinophoragossypiella[11]、环带锦斑蛾Pseudopidorusfasciata[12]。同样,昆虫的交配行为也受内在的生物钟基因控制[13]。研究昆虫的羽化和交配模式,可以帮助人们了解它们的生殖策略,从而选择最佳的防治时机。
条纹小斑蛾Thyrassiapenangae(Moor) 属鳞翅目Lepidoptera、斑蛾科Zygaenidae,是乌蔹莓Japanesecayratia的重要害虫,在江西南昌1年发生4~5代,以老熟幼虫结茧越冬[14-15]。该虫属于典型的长日照型昆虫,短光照诱导滞育,在滞育决定中,光期长度起了重要作用[16]。本文重点测试温度和光周期对条纹小斑蛾的羽化和交配行为节律是否存在影响。
1材料和方法
1.1供试虫源与光暗处理方法
每年9月底,从江西农业大学校园附近(28°46′N,116°50′E)采集末龄幼虫于养虫室内饲养结茧,供次年观察羽化、交配情况和进行世代繁殖。成虫羽化后接到养虫笼中让其自然交配,交配的成虫放入圆柱形塑料杯中产卵。幼虫孵化后立即接入透明的塑料盒(7.5 cm×7.5 cm×6.0 cm)中,在自然条件(均温27.3 ℃)下用乌蔹莓的新鲜叶片、花、果实饲养至结茧,每盒接虫数不少于50头,供实验备用。
成虫羽化和交配均在光照培养箱(LRH-250-G型)中进行,光照强度约为500~700 lx,箱内的温度变化为±0.5 ℃。所有的暗期处理采用不透光的黑袋子进行人工遮光。
1.2不同温度和光周期条件下羽化和交配节律
初孵化的幼虫分别置于25,28和30 ℃,光周期LD16:8饲养至结茧后,即转入光周期LD11:13(光期为早上06:00至下午17:00),LD13:11和LD15:9下,详细观察各条件下成虫的羽化和交配情况,每小时一次并计数。

图1 条纹小斑蛾在光周期LD11:13、LD13:11和LD15:9分别配合25 ℃、28 ℃及30 ℃的羽化(A)和交配(B)节律Fig.1 Circadian eclosion(A) and mating(B) rhythm of Thyrassia penangae underLD11:13、LD13:11 and LD15:9 combined with 25 ℃、28 ℃ and 30 ℃
1.3自然条件下羽化和交配节律
自然条件下连续繁殖的各代幼虫结茧后,观察各代成虫的羽化和交配情况,每小时1次(早上06:00至晚上20:00),并记录羽化和交配数量。
2结果与分析
2.1不同温度和光周期条件下羽化和交配节律
从图1A可以看出,羽化高峰随着温度升高而提前。例如,在LD11:13光周期下,25 ℃、28 ℃和30 ℃条件下,25 ℃下的羽化高峰期出现在光期开始后的第3小时,即上午08:00—09:00;28 ℃下羽化最高峰出现在光期开始后的第2小时,即上午07:00—08:00;30 ℃下羽化高峰期出现在光期开始后的第1小时,即上午06:00—07:00。在LD13:11和LD15:9光周期下,结果也是如此。可见,温度在条纹小斑蛾的成虫羽化事件中起了主要作用,而光周期对此事件影响极小。
从图1B可以看出,在25、28和30 ℃恒温条件下,各光周期的交配时间均随日长的延长而推迟,交配高峰出现的时间在日长11 h时为16:00—17:00,日长13 h时为17:00—18:00,日长15 h时为19:00—20:00,这说明条纹小斑蛾的交配行为与光周期有紧密联系,而与温度关系不大。
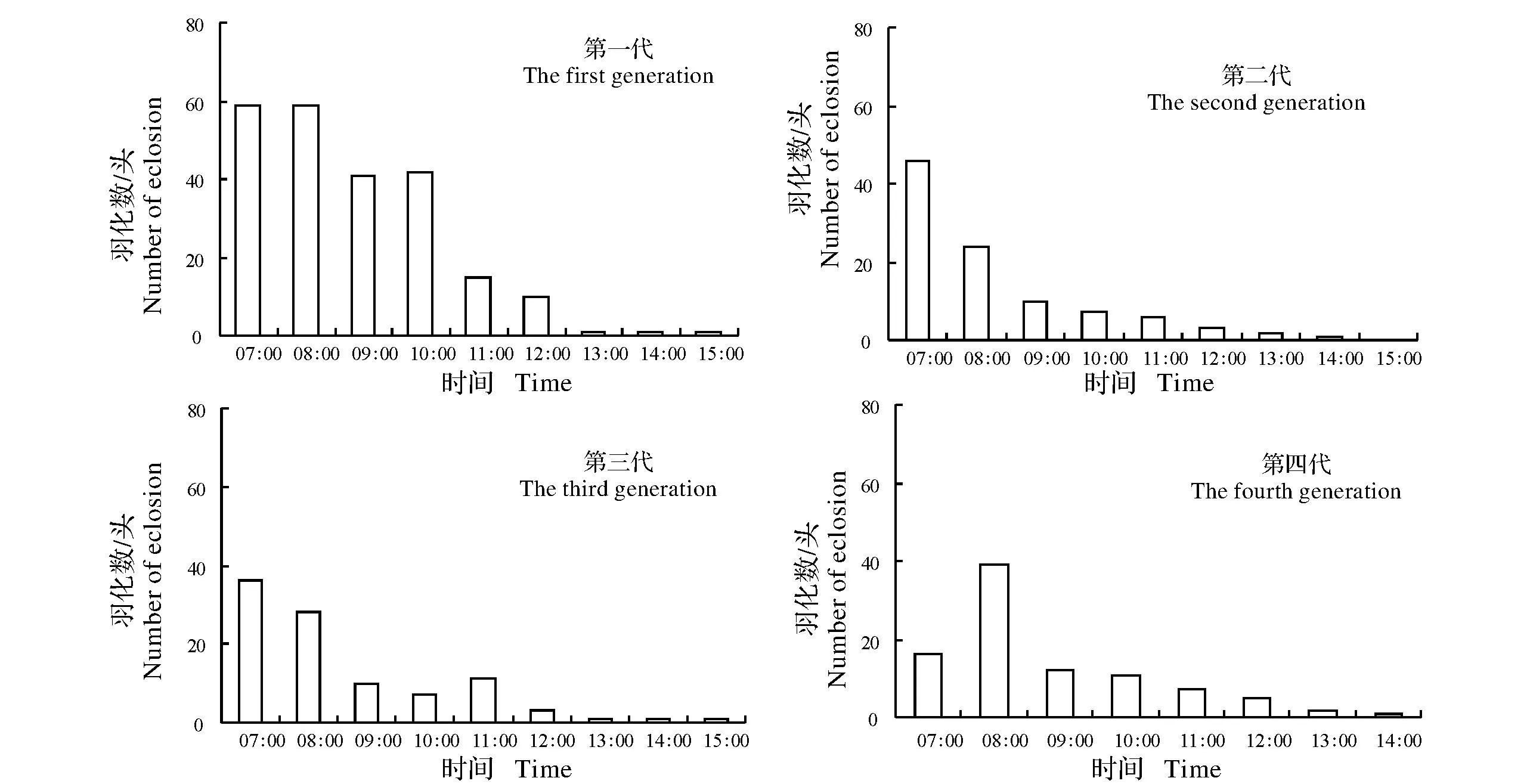
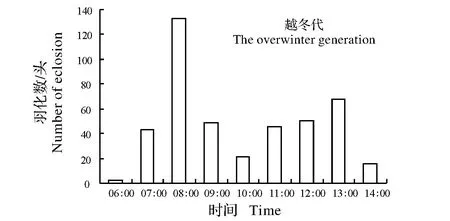
图2 条纹小斑蛾的越冬代(5月,均温22.7 ℃,平均日长约14 h 10 min)、第一代(6月,均温26.1 ℃,平均日长约14 h 45 min)、第二代(7月,均温29.6 ℃,平均日长约14 h 30 min)、第三代(8月,均温29.3 ℃,平均日长约13 h 33 min)和第四代(9月,均温25.1 ℃,平均日长约13 h 5 min)成虫在自然条件下的羽化情况Fig.2 Circadian eclosion rhythm of Thyrassia penangae adults for the overwintering generation (in May when the average temperature was22.7 ℃ and the average daylength was 14 h10 min),the first generation (in June when the average temperature was 26.1 ℃ and the average daylength was 14 h 45 min),the second generation (in July when the average temperature was29.6 ℃ and the average daylength was 14 h 30 min),the third generation (in August when the average temperature was 29.3 ℃ and the average daylength was 13 h 33 min) and the fourth generation(in September when the average temperature was 25.1 ℃ and the average daylength was 13 h 5 min) under the natural conditions
2.2自然条件下羽化和交配节律
图2显示,越冬代成虫羽化高峰为上午07:00—08:00,第一代成虫羽化高峰为上午06:00—08:00,第二、三代成虫羽化高峰为上午06:00—07:00,第四代成虫羽化高峰为上午07:00—08:00。
图3可以看出,越冬代成虫交配高峰为16:00—17:00,第一代成虫交配高峰为17:00—18:00,第二、三代成虫交配高峰为18:00—19:00,第四代成虫交配高峰为下午17:00—18:00。
3结论与讨论
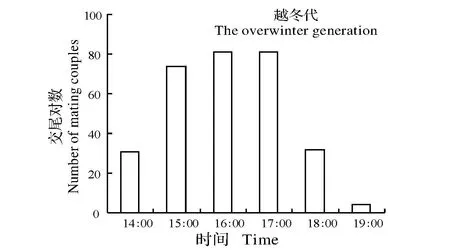

图3 条纹小斑蛾的越冬代(5月,均温22.7 ℃,平均日长约14 h 10 min)、第一代(6月,均温26.1 ℃,平均日长约14 h 45 min)、第二代(7月,均温29.6 ℃,平均日长约14 h 30 min)、第三代(8月,均温29.3 ℃,平均日长约13 h 33 min)和第四代(9月,均温25.1 ℃,平均日长约13 h 5 min)成虫在自然条件下的交配情况Fig.3 Circadian mating rhythm of Thyrassia penangae adults for the overwintering generation (in May when the average temperature was22.7 ℃ and the average daylength was 14 h 10 min),the first generation (in June when the average temperature was 26.1 ℃ and the average daylength was 14 h 45 min),the second generation (in July when the average temperature was29.6 ℃ and the average daylength was 14 h 30 min),the third generation (in August when the average temperature was 29.3 ℃ and the average daylength was 13 h 33 min) and the fourth generation(in September when the average temperature was 25.1 ℃ and the average daylength was 13 h 5 min) under the natural conditions
对条纹小斑蛾羽化节律的观察结果表明,在25 ℃、28 ℃和30 ℃条件下,该虫的羽化最高峰分别出现在光期开始后的第3小时,第2小时,第1小时,三种光周期下结果一致(图1A)。可见,温度在条纹小斑的成虫羽化事件中起了主要作用,羽化高峰随着温度升高而提前,而光周期对此事件影响极小,这与果蝇Drosophilapseudoobscura[17]、环带锦斑蛾Pseudopidorusfasciata[12]羽化节律相类似。自然条件下,条纹小斑蛾越冬代成虫羽化高峰为上午07:00—08:00,第一代成虫羽化高峰为上午06:00—08:00,第二、三代成虫羽化高峰为上午06:00—07:00,第四代成虫羽化高峰为上午07:00—08:00(见图2)。第四代成虫(9月中下旬)羽化高峰比第二(7月中下旬)、三代(8月中下旬)成虫羽化高峰推后了一个小时,可能是因为第四代成虫羽化时的温度更低导致的,与试验结果基本一致。
对条纹小斑蛾交配行为节律的观察结果表明,该虫主要在光期结束前3 h开始交尾,少数交尾事件发生在暗期开始前5~6 h,而且该虫的交尾高峰出现在暗期开始前2 h,在LD11:13、LD13:11、LD15:9下的交尾开始时间随着光期的延长以2小时往后递增,3种温度都显示了类似的结果(图1B)。自然条件下对该虫交配行为的观察也证实了交尾时间随季节日长的延长而推后(图3)。这说明条纹小斑蛾的交配行为与光周期有紧密联系,而与温度关系不大。这与冷杉梢斑螟Dioryctriaabietella[18]类似,而与黑斑皮蠧Trogodermaglabrum[19]相反。
因此,本试验结果的重要意义是揭示了该虫羽化节律主要由温度调节,而其交配节律主要由光周期调节。
参考文献:
[1]Saunders D S.Insect Clocks,3nded[M].Oxford:Pergamon Press,2002:6-13.
[2]Pittendrigh C S.On temperature independence in the clock controlling emergence time inDrosophila.Proc[J].Nat Acad Sci USA,1954,40(11):1018-1029.
[3]Saunders D S.The circadian eclosion rhythm inSarcophagaargyrostoma:some comparisons with the photoperiodic clock[J].J comp Physiol,1976,110(2):111-133.
[4]Smith P H.Responsiveness to light of the circadian clock controlling eclosion in the blowfly,Luciliacuprina[J].Physiol Entomol,1985,10(4):323-336.
[5]Lankinen P and Riihimaa A.Effects of temperature on weak circadian eclosion rhythmicity inChymomyzacostata(Diptera:Drosophilidae)[J].J Insect Physiol,1997,43(3):251-260.
[6]Bünning E.Zur Kenntniss der endogonen Tagesrhythmik bei Insekten und Pflanzen[J].Ber dt bot Ges,1935,53(6):594-623.
[7]Kalmus H.Periodizität und Autochronie (Ideo-chronie) als Zeitregelnde Eigenschaffen der Organismen[J].Biol Generalista,1935,11(1):93-114.
[8]Brett W J.Persistent diurnal rhythmicity inDrosophilaemergence[J].Ann Ent Soc Am,1955,48(1):119 -131.
[9]Palaksha and Shakunthala V.Effect of different light regimes on eclosion rhythm ofDrosophilaagumbensisandDrosophilanagarholensis[J].Biol Rhythm Res,2014,45(3):219-227.
[10]Lewis C B,Bletchley J D.The emergence rhythm of the dung-flyScopeumastercoraria(L.)[J].J Anim Ecol,1943,12(1):11-18.
[11]Pittendrigh C S,Minis D H.The entrainment of circadian oscillations by light and their role as photoperiodic clocks[J].Am Nat,1964,98(2):261-294.
[12]Wu S H,Refinetti R,Kok L T,et al.Photoperiod and temperature effects on the adult eclosion and mating rhythms inPseudopidorusfasciata(Lepidoptera:Zygaenidae)[J].Environ Entomol,2014,43(6):1650-1655.
[13]Sakai T and Ishida N.Circadian rhythms of female mating activity governed by clock genes inDrosophila.Proc[J].Natl Acad Sci USA,2001,98(16):9221-9225.
[14]李仁烈.条纹小斑蛾的初步研究[J].昆虫知识,1988,25(5):283.
[15]何海敏,黄芳,杨东,等.条纹小斑蛾的生物学特性[J].昆虫知识,2009,46(3):411-414.
[16]He H M,Xian Z H,Huang F,et al.Photoperiodism of diapause induction inThyrassiapenangae(Lepidoptera:Zygaenidae)[J].J Insect Physiol,2009,55(11):1003-1008.
[17]Zimmerman W F,Pittendrigh C S,Pavlidis T.Temperature compensation of the circadian oscillation inDrosophilapseudoobscuraand its entrainment by temperature cycles[J].J Insect Physiol,1968,14(5):669-684.
[18]Fatzinger C W.Circadian rhythmicity of sex pheromone release byDioryctriaabietella.[Lepidoptera:Pymlidae (Phycitinae)]and the effect of a diel light cycle on its precopulatory behavior[J].Ann Entomol Soc Am,1973,66(6):1147-1153.
[19]Hammack L,Burkholder W E.Circadian rhythm of sex pheromone-releasing behaviour in females of the dermestid beetle,Trogodermaglabrumregulation by photoperiod[J].J Insect Physiol,1976,22(3):385-388.
Effects of Photoperiod and Temperature on Adult Eclosion and
Mating Rhythms ofThyrassiapenangae(Lepidoptera:Zygaenidae)
HE Hai-min,XIAO Liang,FU Shu,XIAO Hai-jun,XUE Fang-sen*
(Institute of Entomology,Jiangxi Agricultural University,Nanchang 330045,China)
Abstract:In order to explore the occurrence regularity of eclosion and mating behavior of Thyrassia penangae (Lepidoptera:Zygaenidae),whose daily distributions of eclosion and mating activities were recorded under natural and various laboratory conditions.The result indicated that the majority of adults emerged in early morning with an eclosion peak between 07:00 and 08:00 under natural conditions.At the constant temperatures of 25,28 and 30 ℃,the peak eclosion times appeared earlier with increasing temperature,occurring in 08:00—09:00,07:00—08:00 and 06:00—07:00,respectively.Photoperiod had little influence on eclosion time.Under natural conditions,the mating time from overwintering generation to the third generation was postponed with the increase of season daylength,with the peak mating time occurring in 15:00—17:00,16:00—18:00 and 18:00—20:00,respectively.At the constant temperatures of 25,28 and 30 ℃,the mating time was postponed with the increase of daylength,with the peak mating time occurring in 16:00—17:00,17:00—18:00 and 19:00—20:00,respectively.Temperature had little influence on mating time.
Key words:Thyrassiapenangae;eclosion rhythm;mating rhythm;photoperiod;temperature
作者简介:何海敏(1984—),男,实验师,主要从事昆虫生理生态研究,E-mail:hehaimin1984@163.com;*通信作者:薛芳森,教授,E-mail:xue_fangsen@hotmail.com。
基金项目:国家自然科学基金项目(31560608),江西农业大学青年基金项目(QN201109)
收稿日期:2015-02-08修回日期:2015-06-18
中图分类号:Q969.431.6
文献标志码:A
文章编号:1000-2286(2015)06-1011-05
何海敏,肖亮,傅淑,等.光周期和温度对条纹小斑蛾羽化和交配节律的影响[J].江西农业大学学报,2015,37(6):1011-1015.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
中学生数理化·八年级物理人教版(2021年10期)2021-11-22
疯狂英语·新读写(2021年8期)2021-11-05
蔬菜(2020年4期)2020-12-14
浙江农业学报(2019年7期)2019-08-14
浙江农业学报(2018年9期)2018-10-11
小学生优秀作文(高年级)(2018年4期)2018-09-11
少儿科学周刊·少年版(2017年7期)2017-09-29
少儿科学周刊·儿童版(2017年7期)2017-09-29
少儿科学周刊·儿童版(2016年1期)2016-03-14
