
长江口及闽浙海域浮游植物动力学模型研究
2015-03-02 01:30:08王一鹤吴辉朱建荣沈健
华东师范大学学报(自然科学版) 2015年4期
王一鹤, 吴辉, 朱建荣, 沈健
(1.华东师范大学 河口海岸学国家重点实验室,上海 200062;2.威廉玛丽学院 弗吉尼亚海洋研究所,费吉尼亚 23062,美国)
长江口及闽浙海域浮游植物动力学模型研究
王一鹤1, 吴辉1, 朱建荣1, 沈健2
(1.华东师范大学 河口海岸学国家重点实验室,上海 200062;2.威廉玛丽学院 弗吉尼亚海洋研究所,费吉尼亚 23062,美国)
基于一个三维、高分辨率、水动力泥沙生态数值模式,综合考虑多种物理及生态因子,模拟研究了长江口及闽浙海域甲藻和硅藻两种浮游植物生物量的时空分布规律.数值实验的结果表明,该海域藻类生长主要集中在春末、夏季及秋初,硅藻有春季和夏季两次水华,而甲藻仅在春季发生一次水华.该海域浮游植物的高值区主要集中在30-50 m等深线附近.该处是外海高温、低浊度水体与近岸低温、高浊度、高营养盐长江冲淡水的交汇区域,满足浮游植物生长所需要的外部物理环境和内部营养盐需求.本文进一步探讨了径流、陆架环流等关键物理过程对浮游植物生长的影响.
长江口及闽浙海域; 浮游植物; 物理过程; 数值模拟
0 引 言
浮游植物作为海洋生态系统的主要初级生产者,是该生态系统中的重要组成部分,其生物量的多少很大程度上决定着该海域的水产性能[1].因此,研究浮游植物动力学[2],即研究浮游植物在物理动力驱动下受营养盐限制、浮游动物摄食以及自身新陈代谢等一系列作用影响的生物量时空分布规律,对于研究海区营养传递及能量流动,以及进一步研究赤潮形成的外界环境和物理过程机制有着重要的意义.
以往的研究表明,限制浮游植物生长的因子通常包括光照、水温、和营养盐等[3].目前,已经有一套较为成熟的理论来解释大洋中一年内浮游植物生物量的变化特征.冬季,由于水体的混合作用使沉积于底层的营养盐带到表层,但由于水温的限制作用,浮游植物未能达到生长需求;春季,温度开始回升,自冬季储存的营养盐在适合的温度条件下被浮游植物吸收,浮游植物生物量剧增;夏季,由于表层水体受太阳辐射作用持续升温造成水体层化现象明显,不利于底层的营养盐向上输送,同时由于春季水华造成的海表面营养盐匮乏,此时浮游植物生物量剧增现象得到缓解;秋季,水温开始下降,层化现象逐渐被打破,表层营养盐得到了补充,故使浮游植物生物量再次增加[4,5].
然而,河口及近岸地区相对于大洋动力因素更为复杂,同时还涉及径流等陆源营养盐的输入.河口地区的关键物理过程如何对浮游植物生物量的时空变化产生影响仍然存在一些不明朗的地方,尤其缺少定量研究.因此,本文基于研究组已经建立的三维高分辨率、渤黄东海水动力泥沙生态动力学数值模式,模拟研究长江口及闽浙海域的初级生产力过程;探讨径流和陆架环流等关键物理过程对浮游植物生物量时间和空间分布上的影响.
1 研究区概况
长江口及闽浙海域一直是我国东南沿海关注的焦点区域.这里动力条件复杂,且季节性变化明显,深受长江冲淡水、台湾暖流、沿岸流及季风的影响.冬季,受北风和科氏力的影响,长江冲淡水出口门外向南输送,在闽浙沿岸由东北向西南运动,给闽浙海域带来了低温和高营养盐的水体,另一方面,高温的台湾暖流沿长江口外水下河谷流向长江口区域,这两股水流在闽浙海域附近交汇并在舟山群岛至台湾海峡中北部海域形成一条海洋锋面;夏季,由于东南季风的作用,长江冲淡水在出口门外先向东南方向运动,随后转向东北,形成冲淡水羽状锋,而闽浙海域的沿岸流也随着长江冲淡水的转向逐渐消退,锋面也逐渐消失[6,7].
长江每年携带大量的营养盐汇入东海,同时受外海台湾暖流及黑潮次表层水入侵所携带的大量磷酸盐和硅酸盐影响,长江口附近海域浮游植物特点显著.据近几年的调查结果显示,浮游植物生物量较高的时期主要集中在春季和夏秋季,种类上以硅藻门和甲藻门为主,硅藻细胞丰度占优[8-10].近些年来,目标海域赤潮问题愈演愈烈,其中诸如米氏凯伦藻和东海原甲藻的赤潮出现频率增加[11].从《2013年中国海洋环境状况公报》(http://www.coi.gov.cn/)[12]可以看到,2009—2013年间,东海作为我国赤潮事件爆发最多的海域,其引发赤潮第一优势种为东海原甲藻.闽浙海域赤潮爆发的地点主要集中在30~50 m等深线附近(见图1),此区域正是长江冲淡水与外海水体的交汇区域,外海的高温低浊度水体与长江冲淡水的高营养盐水体在此交汇,满足了浮游植物生长所需要的外部环境和内部营养需求,从而造成了浮游植物生物量的剧增.

图1 研究区域地形图Fig.1 Topographic map of the study area
2 研究方法
本文采用的数值模式为ECOM-si[13],其来源于普林斯顿海洋模型POM[14],并采用HSIMT-TVD平流格式求解物质输送方程[15].模型的生态模块移植自FVCOM的可变生态模型FBM[16,17],选择了包括氮盐、磷酸盐、硅藻、甲藻、浮游动物及碎屑共6个生态变量的N2P2ZD生态动力学模块,建立长江口及闽浙海域生态动力学数值模型.具体的模式设置如下.
2.1 模式范围和网格、计算时间
模式的范围为一扇形,包括整个长江河口及渤黄东海,东边到西太平洋边缘及日本海区域(133°E附近),北边囊括整个渤海,南边到达台湾岛以南(22°N附近)(见图2).模式网格分辨率较高,所关心的长江口及闽浙海域网格分辨率从长江口内的几百米到冲淡水扩展区的2~3 km.网格正交性、平滑性、岸线拟合和局部加密均得到了很好的体现,这也保证了模式的稳定性和计算精度.模式垂向采用σ坐标,将垂向分为非均匀的20层,取表层的厚度更薄,是为了着重研究表层水体中的生物过程以及表层冲淡水对浮游植物生物量变化的影响.此数值模式已成功的应用于长江冲淡水的模拟研究[18,19].

图2 模式计算范围和网格Fig.2 Calculating scope and grid of the numerical model
因水体温度、盐度以及营养盐等对外部动力的响应调整较慢,为使模式能更好的模拟生物过程,在模型验证及数值试验中,模式的计算时间均为三年.模式第一年只计算物理场,计算参数包括流速、温度、盐度等;第二年打开营养盐输入开关,使营养盐以保守物质输入;第三年开始计算耦合了生态模块的水动力模式.
2.2 N2P2ZD生态动力学模块介绍
N2P2ZD生态动力学模块包含氮盐、磷酸盐、硅藻、甲藻、浮游动物以及生物碎屑6个生态变量.考虑的生物过程包括光合作用、呼吸作用、死亡作用、动物摄食,矿化作用以及种间竞争等(见图3).浮游植物通过光合作用按照redfield比(N∶P=16∶1)摄取营养盐,光合作用同时还受温度和光强的限制.浮游植物通过呼吸作用按照redfield比转化为营养盐;通过死亡作用转化为碎屑;同时,浮游植物被浮游动物摄食.浮游动物在摄食浮游植物过程中伴随着主动呼吸消耗以及摄食损失,仅有部分被摄食浮游植物被转化为浮游动物组织,其余部分伴随着主动呼吸消耗转化为营养盐以及由于摄食损失被转化为碎屑;浮游动物同样包含呼吸作用和死亡作用,产物同浮游植物.碎屑通过矿化作用按照redfield比被矿化为营养盐.生态模块与外部环境之间的关系包括河流边界及外海开边界的营养盐输入,平流扩散作用及沉降作用,水温及光照对各生物作用的限制等.
3 模型设置及验证
3.1 模型设置

图3 生态模块示意图Fig.3 Ecological module diagram
本文共设计了6组数值试验,1组用于营养盐资料的验证,采用2005至2007年1 h一个的实际观测长江径流量(大通水文站实时观测资料)、6 h一个的风资料(取自NCEP)驱动模式.外海开边界条件为陆架环流和潮流的叠加,陆架环流和潮流皆以流通量形式给出.陆架环流为SODA月平均数据,潮流由11个主要分潮组成.模型温盐初始场资料源于SODA数据.海气热通量由大气相对温度、压强、湿度、云量等资料计算得到,上述资料均来源于NCEP.另外,模型的生态模块初始场和边界场值仅考虑了营养盐的边界条件,其余生态变量的初始值皆为0.01 Nmol/m3.营养盐的径流边界根据他人研究资料给出[20],外海开边界及初始场皆源于SODA数据.
另外5组实验均是气候态下试验,用于分析该海域浮游植物分布特点以及径流量及陆架环流作用对浮游植物生物量变化的影响.控制实验除了长江径流量和风资料采用多年月平均条件外,其他条件与第一组试验皆一致.其余四组为敏感性实验,分别改变单一物理动力因子,具体的实验条件见表1.由于长江径流和陆架环流是此区域最重要的营养盐来源,因此本文只讨论这两个因素的影响.实验中改变径流和陆架环流皆只改变流量值,并没有改变边界输入的营养盐浓度.
3.2 模型验证
本文所采用的水动力模型相对较为成熟,能准确的反映出长江口及附近海域的流场、温度及盐度,并在许多实例中得到运用.关于物理场的验证可参考先前的研究论文[19,20].

表1 各组敏感性实验设置Tab.1 Settings of each sensitivity experiment
3.2.1 营养盐验证
营养盐作为浮游植物生长的营养来源,直接决定了浮游植物的生物量.因此,正确的模拟营养盐是一个生态动力学模型的根本.本文利用2007年5月闽浙海域走航的观测营养盐数据进行验证.图4、图5显示的是观测值与该时段该区域模型计算的氮盐及磷酸盐对比.从图中可以看出,模型计算结果良好,能较好的展现出营养盐的水平和垂向结构.氮盐表层浓度大于底层,而磷酸盐恰恰相反.表层的营养盐主要由长江径流携带向外海输运,出口门外向南沿岸输送,与此时外海的低营养水体混合,在50m等深线附近形成一条营养盐的梯度锋面.底层营养盐主要来源于外海台湾暖流及黑潮次表层陆架环流水体的入侵,特别是磷酸盐,观测及模型皆呈现出50m等深线以东底层磷酸盐浓度大于表层的特征.

注:左边为观测资料,右侧为模型验证结果;上方两张图反映的是氮盐的验证结果,单位为mmol N/m3,下方两张图反映的是磷酸盐的验证结果,单位为mmol P/m3;图5与此图设置相同图4 表层营养盐验证结果Fig.4 Surface nutrient concentration verification
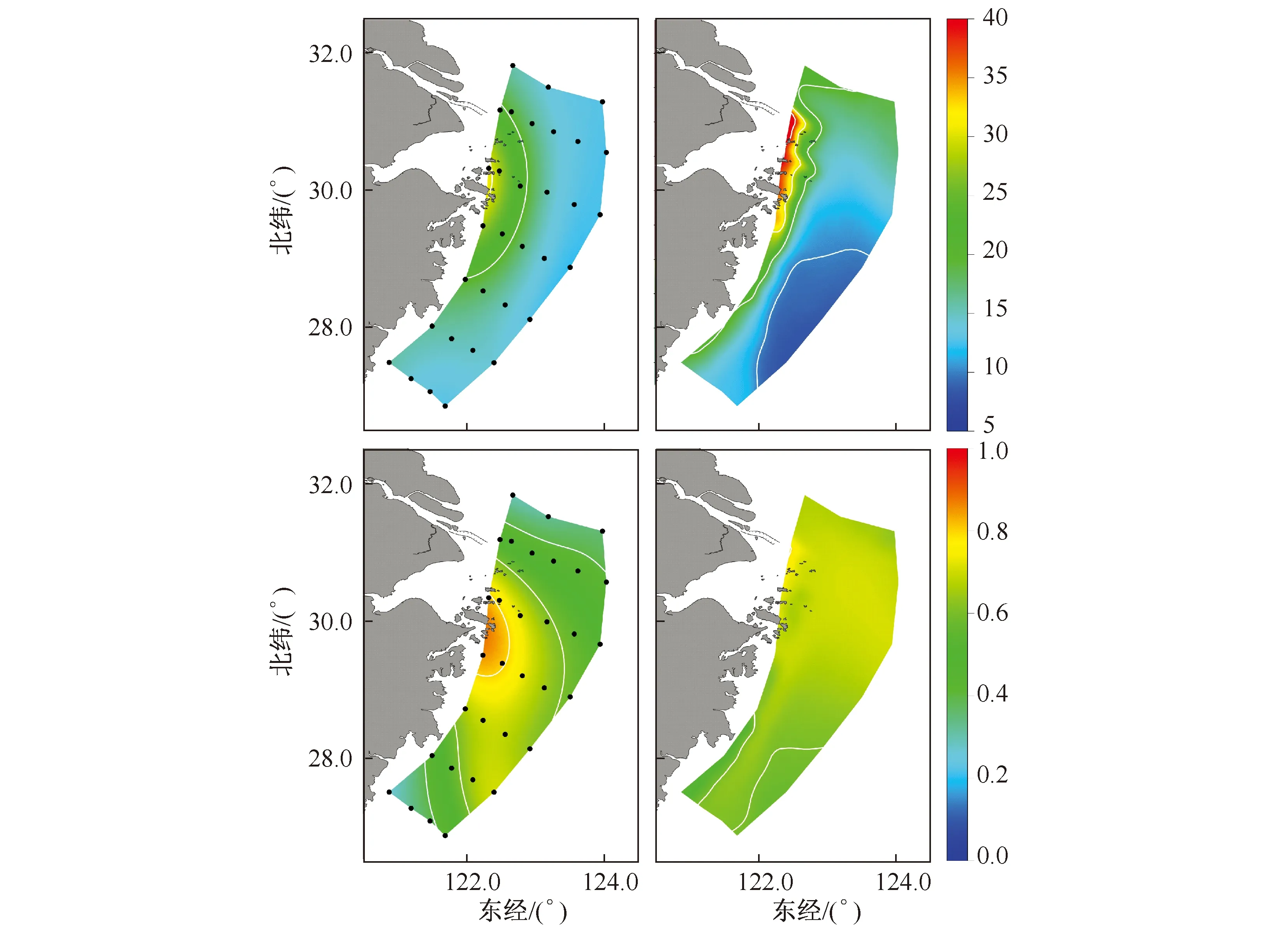
图5 底层营养盐验证结果Fig.5 Bottom nutrient concentration verification
3.2.2 浮游植物分布
相对于保守或半保守的温度、盐度和营养盐等数值模拟,浮游植物的定量模拟目前还普遍存在困难,其原因为营养盐的摄取通常采用了营养盐半饱和方案,且并没有考虑浮游植物孢子休眠等复杂生物过程,相关生态过程参数也仅能经验性的确定.同样的,本研究模式模拟的浮游植物分布在定量上存在一定的误差,但在定性上和历史资料比较相似,能较好的反应浮游植物的分布特点.对浮游植物的分布特点将在下一章中进行详细的介绍.
4 生态动力学模型结果讨论
4.1 气候态下浮游植物春夏季模拟结果
模型分别计算了硅藻和甲藻两种藻类,统计了模型1—9月(0—270日)研究区域(118°—124°E、24°—32°N)表层平均浮游植物生物量浓度值,如图6.从图中我们得知,甲藻的整体生物量相对于硅藻偏低.分析其原因,是因为甲藻的生长率比硅藻小,因此在温度光照营养盐等条件皆满足的情况下,甲藻的整体生物量不及硅藻.甲藻生物量的高值主要集中在春季(90—180日),而硅藻在整个春夏秋都保持着很高的生物量.这是因为甲藻生长的适宜温度相较于硅藻较低,进入夏季之后,水温较高,对甲藻的生长产生了温度抑制作用.甲藻仅在春季发生一次水华,而硅藻在春季和夏秋季发生两次水华.这是因为在春夏交替的季节,由于春季水华的发生,表层的营养盐消耗殆尽,甲藻相较于硅藻更加能适应低营养盐的环境,故在120—150日这段时间里,硅藻的生物量有略微下降的现象,而此时甲藻生物量依旧在持续增加.

图6 模型计算的硅藻、甲藻1—9月表层平均浓度变化Fig.6 Model calculating biomass concentration of diatom and dinflagellata during the time from January to September
将模型计算输出的表层结果从4—9月做逐月平均处理,进一步分析春末、夏及秋初长江口及闽浙海域浮游植物生物量的分布特点.硅藻和甲藻的空间分布结果见图7.从浮游植物时空分布特点来看,浮游植物的生物量主要集中在30~50m 等深线附近,而这里正是近岸长江冲淡水与外海水体的交汇区.近岸水体虽然携带了大量营养盐,但是同时也携带了大量其他陆源悬浮物,导致该区域水体透明度较低,从而受到浮游植物生长的光限制,远岸水体虽然透明度较高,但由于水体携带营养盐较少,故亦不适合浮游植物生长.恰恰在两者的交汇区,由于水体的交换,近岸营养盐向外扩散,且摆脱光限制,从而造成浮游植物生物量的暴增.4月初,由于长江冲淡水出口门后沿岸向南输运,冲淡水的高浊度低温水体不适合浮游植物生长,故在浙江沿岸及杭州湾地区有一条明显的浮游植物真空带;而离岸较远的地区以及福建及台湾海峡附近,由于浊度降低,加上富营养盐的长江冲淡水和温暖的台湾暖流水相混合,促进了浮游植物的生长.进入5、6月,海表面温度增加,浮游植物的生长区域进一步向北扩展,但是杭州湾及长江口门内由于泥沙含量较大,水体透明性较差,依然不适宜藻类的生长.另一方面,由于4月浮游植物在适合条件下的生物量爆发导致该区域营养盐不足,相对而言,甲藻比硅藻更加适应低营养盐的环境,故在5、6月甲藻生物量稳定增加,而硅藻在闽浙海域由于营养盐供给不足出现了浮游植物生物量下降的现象.7、8月,虽然此时长江冲淡水在东南风的驱动下不再沿岸向南输送,但由于夏季台湾暖流的增强,同样带来了足够的营养盐,特别是磷酸盐的补给又给予藻类再次爆发的可能.随着温度进一步升高,由于甲藻的不耐高温性,甲藻生物量明显下降,而硅藻此时顺势爆发.9月,再一次面临营养盐供给不足的问题,硅藻生物量明显下降,而甲藻在温度回落以及低营养盐状态下生物量反而有所增加,此时北部相较于南部浮游植物生物量较高,这是因为北部离长江口较近,仍能得到一定的营养盐补充.
4.2 物理因素变化对浮游植物的响应
长江径流和陆架环流作为影响长江口及闽浙海域两个主要的物理过程,对该海域水文物理场的分布起到至关重要的作用,同时,长江径流和陆架水流入侵携带的营养盐又为浮游植物的生长提供营养来源.因此,为进一步研究径流量和陆架环流作用对浮游植物生物量时空变化的影响,本文分析了另外4组单一物理因子变化试验.由于进入夏季,在温度营养盐条件都较为符合的条件下,营养盐及浮游植物参与生态循环作用显著,物理因素的改变对浮游植物生物量改变研究起来就显得更为复杂,故本文着重分析4月(即浮游植物水华首发时间)时物理因素变化对浮游植物的影响.以甲藻为例,各敏感性实验结果见图8.
4.2.1 长江径流的影响
如图所示,径流量的改变基本上只对长江口门附近的浮游植物产生影响,在远离口门的闽浙海域几乎看不出浮游植物生物量的变化.这可以说明,在4月,闽浙海域及以南海域浮游植物的生长对长江径流变化的响应不敏感.从某种程度上这是可以理解的,因为闽浙海域营养盐的集聚是一个长时间冲淡水累积过程,且营养盐总体较为充沛,不太受长江径流波动的影响.

注:图中左一自上而下分别为硅藻4、5、6月表层分布,左二自上而下为硅藻7、8、9月表层分布,左三是甲藻4、5、6月表层分布,最后一列为甲藻7、8、9月表层分布;单位为mmol N/m3图7 硅藻、甲藻各月平均表层分布Fig.7 Surface average monthly concentration of diatom and dinflagellata

注:第一行从左至右分别是控制实验、实验一、实验二,第二行依次是实验三、实验四;单位为mmol N/m3图8 各实验4月表层甲藻分布Fig.8 Surface average monthly dinflagellata concentration of each sensitivity experiment in April
4.2.2 陆架环流的影响
陆架环流的增减对甲藻生物量变化显著.陆架环流的增强或减弱造成了甲藻高值区的南北移动.当陆架环流增强时,甲藻生物量的高值区向北偏移,而陆架环流减弱时则出现相反的现象.陆架环流的强弱会影响该海域的营养盐和温度分布,从而改变浮游植物的生长条件.
从营养盐的分布角度来说(见图9和图10),虽然陆架环流同样能够带来营养盐特别是磷酸盐的补充,但是由于该区域在浮游植物生物量未爆发前本身就积累了大量营养盐,且其值由于长江冲淡水一个冬季的积累往往比陆架环流补充的值要大.因此,总体上,本区域营养盐总量比较充沛,在爆发初期的4月份不至于成为一个显著的限制因素.
我们发现海水温度的变化可能是更重要的一个因素(见图11).在控制实验下,可以看到闽浙海域的温度存在近岸低、远岸高和北部低、南部高的特点.在这样的情况下,温度和营养盐一样成为限制浮游植物生长的重要因素.东海主要的甲藻种东海原甲藻可以忍受低营养盐环境并种群增长,其最适温度约为22 ℃,适温范围在15~25 ℃之间[21].只有当“富营养盐水体”的温度上升到适宜的生长温度(大于15 ℃)后,初级生产力才可能爆发.由于陆架环流、特别是台湾暖流和黑潮带来了大量高温水体,因此对本区海温的回升起到重要作用,特别是近岸温度较低但富含营养盐的水体.在陆架环流减弱的情况下,闽浙海域的海温显著降低(降幅超过1 ℃),因此海温适宜浮游植物生长的区域发生南移.长江口和舟山以东海域,由于水温过低, 初级生产力受抑制.相反,在陆架环流增强的情况下,高温水体可以输送

注:第一行从左至右分别是表层控制实验、实验三、实验四,第二行依次是底层控制实验、实验三、实验四;单位为mmol P/m3图10 各控制实验4月表底层磷酸盐分布Fig.10 Average monthly phosphorus concentration of each sensitivity experiment in April
到更北的区域,使得整个海域的表层温度升高超过1 ℃.这显著减轻了北部区域富营养盐水体的浮游植物生长温度限制,从而促使该区域初级生产力的爆发.影响浮游植物生长的因素很多,温度只是其中的一个方面.同时,陆架环流的增强或减弱不仅是改变了海温,还会对近岸水体的离岸交换产生影响.这些都是值得深入研究的问题,但受篇幅限制本文不做展开讨论.

注:左为控制实验下的SST,中为实验三的SST变化(和控制实验相减),右为实验四的SST变化;单位为℃图11 各实验下的海水表面温度(SST)比较Fig.11 Sea surface temperature comparison
5 总 结
本文利用了一个三维泥沙生态水动力模型,初步探讨了长江口及闽浙海域春夏季浮游植物生长规律,并分析了径流和陆架环流等物理因子的改变对浮游植物生物量变化的影响.本文利用径流及风等的实际数据模拟了长江口及闽浙海域的营养盐分布情况,结果较为合理,并将模型计算的浮游植物结果与近些年来该海域初级生产力爆发事件的规律进行对比,情况也基本与事实相符.之后,本文就浮游植物的分布特点展开讨论,分析其动力机制.最后,本文利用单动力因子实验,分析关键物理因子对浮游植物分布的影响.
浮游植物生长主要受营养盐、温度和光照的限制.在长江口及闽浙海域,浮游植物水华主要发生在春季和夏季.甲藻仅在春季发生一次水华,而硅藻在春季和夏季发生两次水华.从浮游植物空间分布来看,其高值区主要集中在30~50 m等深线附近.此区域由于受长江冲淡水与外海的混合作用,在特定条件下满足了浮游植物生长所需要的外部环境和内部营养需求,从而造成了浮游植物生物量的剧增,而近岸由于浊度高、远岸由于营养盐浓度较低皆不适合藻类生长.另一方面,由于受到温度及营养盐供给的影响,浮游植物在时间上的空间分布也呈现一定的规律,春季高值区偏南,而夏秋高值区逐渐往北移动.
径流和陆架环流能改变浮游植物生长所需要的营养和外部环境,从而改变浮游植物生物量的时空分布.从模型计算的该海域4月结果来看,径流对目标区域浮游植物生物变化影响较小,且基本位于长江口门附近;陆架环流的强弱能造成浮游植物生物量空间上的北南移动,陆架环流增强时,浮游植物生物量高值区向北移动,而陆架环流减弱时呈现出相反的特征.这与陆架环流改变导致的温度场变化有关.
综上所述,本研究初步揭示了长江口及闽浙海域浮游植物生物量的时空分布特征,并且分析了径流及陆架环流流量变化对浮游植物生物量变化的影响.但物理因子如何具体作用于浮游植物的外部环境和营养需求的探讨还不够深入,这是今后继续研究的方向.
[1] 陆健健. 河口生态学[M]. 北京: 海洋出版社, 2003.
[2] 魏皓, 赵亮, 武建平,等. 浮游植物动力学模型及其在海域富营养化研究中的应用[J]. 地球科学进展, 2001, 16(2): 220-225.
[3] ZHANG J. Nutrient elements in large Chinese estuaries[J]. Continental Shelf Research, 1996, 16(8): 1023-1045.
[4] FINDLAY H S, YOOL A, NODALE M, et al. Modelling of autumn plankton bloom dynamics[J]. Journal of Plankton Research, 2005, 28(2): 209-220.
[5] 宋洪军, 季如宝, 王宗灵. 近海浮游植物水华动力学和生物气候学研究综述[J]. 地球科学进展, 2011, 26(3): 257-265.
[6] 朱建荣,沈焕庭. 长江冲淡水扩展机制[M]. 上海:华东师范大学出版社,1997.
[7] 黄荣祥. 台湾海峡中、北部海域温、盐度特征[J]. 海洋科学, 1989(6): 33-38.
[8] 栾青杉, 孙军. 2005年夏季长江口水域浮游植物群集特征及其与环境因子的关系[J]. 生态学报, 2010, 30: 4967-4975.
[9] 赵冉, 白洁, 孙军,等. 2006年夏季长江口及其邻近水域浮游植物群集[J]. 海洋湖沼通报, 2009(2): 88-96.
[10] 林军, 朱建荣, 张经,等. 长江口外海区浮游植物生物量分布及其与环境因子的关系[J]. 水产学报, 2011, 35(1): 74-87.
[11] 栾青杉, 孙军. 2005年秋季长江口及其邻近水域浮游植物群集[J]. 长江流域资源与环境, 2010, (2): 202-208.
[12] 国家海洋局. 2013年中国海洋环境状况公报[EB/OL]. (2014-03)[2014-03-31]. http://cn.chinagate.cn/economics/2014-03/31/content_31952090_7.htm
[13] BLUMBERG A F. A primer for ECOM-si[R]. Technical Report of HydroQual, Mahwah N J,1994:66.
[14] BlUMBERG A F, MELLOR G L. A description of a three-dimensional coastal ocean circulation model[J]. Three-Dimensional Coastal Ocean Models, 1987:1-16.
[15] WU H, ZHU J. Advection scheme with 3rd high-order spatial interpolation at the middle temporal level and its application to saltwater intrusion in the Changjiang Estuary[J]. Ocean Modelling, 2010, 33(1-2): 33-51.
[16] CHEN C S, BEARDSLEY R C, COWLES G. An unstructured grid, finite-volume coastal ocean model: FVCOM User Manual[R]. SMAST/UMASSD, 2006.
[17] 林军. 长江口外海域浮游植物生态动力学模型研究[D]. 上海:华东师范大学, 2011.
[18] WU H, ZHU J, SHEN J, et al. Tidal modulation on the Changjiang River plume in summer[J]. Journal of Geophysical Research: Oceans (1978-2012), 2011, 116(C8):192-197.
[19] WU H, SHEN J, ZHU J, et al. Characteristics of the Changjiang plume and its extension along the Jiangsu Coast[J]. Continental Shelf Research, 2014, 76(2):108-123.
[20] GAO L, LI D, ZHANG Y. Nutrients and particulate organic matter discharged by the Changjiang (Yangtze River): Seasonal variations and temporal trends[J]. Journal of Geophysical Research: Biogeosciences (2005-2012), 2012, 117(G4):197-205.
[21] 陈炳章, 王宗灵, 朱明远,等. 温度、盐度对具齿原甲藻生长的影响及其与中肋骨条藻的比较[J]. 海洋科学进展, 2005, 23(1): 60-64.
(责任编辑 李万会)
Numerical simulation on the phytoplankton dynamics in Changjiang estuary and Min-Zhe coastal waters
WANG Yi-he1, WU Hui1, ZHU Jian-rong1, SHEN Jian2
(1.StateKeyLaboratoryofEstuarineandCoastalResearch,EastChinaNormalUniversity,Shanghai200062,China;2.VirginiaInstituteofMarineScience,CollegeofWilliam&Mary,VA43062,USA)
This study was based on a high-resolution, three-dimensional, physics-biogeochemistry coupling numerical model that considered multiple dynamics and biochemical factors to simulate the spatial and temporal distribution of the dinoflagellata and diatoms. The results showed that the algae bloomed in late spring, summer and early autumn in this area. The diatoms bloomed twice in spring and summer and dinflagellata bloomed only once in spring. The high phytoplankton mass concentrated around the area of 30 to 50 isobath, which was an frontal area between the high temperature and low turbidity shelf waters and the low temperature and rich nutrient Changjiang diluted water. This area satisfied the physical and biogeochemical requirements for the growth of the phytoplankton. Furthermore, we also analyzed the influences of the physical dynamics such as runoff and shelf circulation on the phytoplankton growth.
Changjiang estuary and Min-Zhe coastal waters; phytoplankton; physical processes; numerical simulation
1000-5641(2015)04-0097-13
2014-10
国家重点基础研究发展规划(2011CB409801);河口海岸学国家重点实验室科研业务费 (2013RCDW01)
王一鹤,男,硕士研究生,从事河口海岸动力研究.E-mail:wyhe11909045@gmail.com.
吴辉,男,副教授,研究方向为河口海岸学.E-mail:hwu@sklec.ecnu.edu.cn.
P731.2
A
10.3969/j.issn.1000-5641.2015.04.011
猜你喜欢
吉林大学学报(地球科学版)(2024年3期)2024-06-03 13:22:07
大自然探索(2023年7期)2023-11-14 13:07:36
课堂内外(小学版)(2023年9期)2023-10-11 14:39:26
沉积学报(2023年1期)2023-04-29 18:57:49
海洋通报(2022年2期)2022-06-30 06:07:04
黑龙江水利科技(2020年8期)2020-01-12 06:26:00
学与玩(2019年8期)2019-10-29 03:32:16
资源节约与环保(2019年10期)2019-01-21 00:22:48
上海建材(2018年1期)2018-04-18 12:15:16
中国科技博览(2016年25期)2016-12-20 18:28:41
