
自发特质推理的神经基础*——基于“镜像系统-心理化系统”模型的解析
2015-02-25 05:07贾磊,张常洁,张庆林
心理学探新 2015年5期
自发特质推理的神经基础*
——基于“镜像系统-心理化系统”模型的解析
贾磊1,张常洁1,张庆林2
(1.浙江工业大学心理系,杭州 310014;2.西南大学心理学部,重庆 400715)
摘要:自发特质推理是一种无需意识努力即可从他人行为描述自动推论其人格特质的社会认知过程;它与心理理论及内隐加工均存在紧密联系。本文从Van Overwalle“镜像系统-心理化系统”的社会脑模型出发,围绕二者在行为知觉编码及特质表征下的功能特性,及其与语义记忆系统和中枢控制系统的协作联系,分析了自发特质推理的认知过程及其神经基础。
关键词:自发特质推理与有意特质推理;心理化系统;镜像系统
1引言
人格特质是个体稳定且持久的内部心理特征,因此常被用以概括、预测和解释行为。基于行为描述做出特质归纳与判断是特质推理的主要表现,也是人们在复杂环境中快速认识他人的重要途径。特质推理不仅与知觉的对象特征有关,而且与主体内化的自身经验、信念期望及社会图式紧密联系,并会因内隐人格理论的不同而存有个体差异。因此它也被视为心理理论(Theory of Mind)的核心成分,与个体社会化发展密切相关(Uleman,Adil Saribay,& Gonzalez,2008;王美芳,陈会昌,2009)。根据意识参与程度及加工流畅性的不同,特质推理具有两种表现型:一是有意识参与且带有意图的有意特质推理(intentional trait inferences,ITI),二是无需意识努力且能自动发生的自发特质推理(spontaneous trait inferences,STI)。
与有意特质推理相对,自发特质推理是一种无需意识努力,即可从他人行为自动推论其人格特质的社会认知过程(Uleman,Rim,Adil Saribay,& Kressel,2012)。依据经典过程理论观点,自发特质推理实质是有序的特质加工过程,包含三个内隐加工阶段:一是行为知觉编码诱发的特质概念激活阶段,二是特质概念与有关信息形成自动联结的特质归因阶段,三是特定环境下基于联系做出特质判断的特质影响阶段。虽然过程理论仅就自发特质推理的认知过程给出了行为层面的构想,它并不能突破黑箱困境以阐述动态的神经加工过程;但毫无疑问,掌握从行为编码到特质表征,再到特质判断的动态过程,是弄清楚自发特质推理认知过程的关键(Skowronski,Carlston,Mae,& Crawford,1998)。
2镜像系统、心理化系统与自发特质推理过程
2.1镜像系统和心理化系统
随着神经科学的发展,研究者发现,社会行为与人格特质作为特殊信息,其认知加工有赖于特殊的社会认知神经表征系统——镜像神经元系统(Mirror Neuron System)的参与。这在方法上提供了一种新的可能,即从社会认知特有的神经基础出发,分析和阐释社会心理与行为产生的神经机制,进而建构理解社会心理的社会脑模型。社会认知神经科学(Social Cognitive Neuroscience)这一学科视角也由此诞生(Ochsner & Lieberman,2001)。
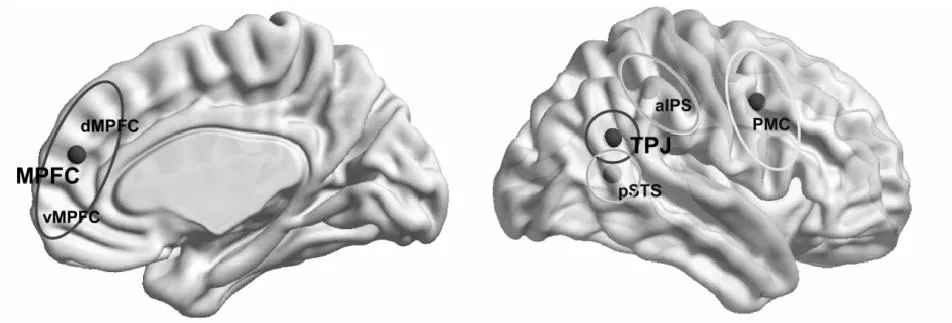
图1 心理化系统(黑圈)与镜像系统(灰圈)的主要脑区
注:(1)心理化系统:内侧额叶皮层(MPFC);背内侧前额皮层(dMPFC);腹内侧前额皮层(vMPFC);颞顶连接区(TPJ);(2)镜像系统:后部颞上沟(pSTS);前部顶内沟(aIPS);前运动皮层(PMC)。
从该视角出发,布鲁塞尔自由大学的Van Overwalle团队基于自身研究,将经典的镜像神经元系统分离为镜像系统(mirror system)和心理化系统(mentalizing system)两大模块,构建起新的社会脑模型(图1)。其中镜像系统因其对行为的“镜像”知觉功能得名,它主要承担对行为的知觉编码,负责快速识别他人行为并将其与记忆中的行为模板进行模式匹配,从而类似镜像过程一样,完成行为的知觉识别并为其模仿提供基础。心理化系统则因其心理理论功能而得名,它是特质表征及信念判断的承担者,负责从知觉到的行为信息中提取并分析他人意图,从而读懂他人想法并理解其观点和信念(Van Overwalle & Baetens,2009)。作为对镜像神经元理论的发展,该模型通过引入心理化系统,尝试将该理论所支持的心智模拟论同经典心理理论结合在同一模型框架下。实际研究中,该团队以自发特质推理这一典型内隐社会认知现象为突破点,开展模型的验证与完善工作。本文即综合该模型有关镜像系统及心理化系统的理论解释,分析阐述自发特质推理的认知机制。
2.2镜像系统与特质激活中的行为编码
2.2.1镜像系统的构成及认知功能
镜像系统包含后部颞上沟、前部顶内沟及前运动皮层三部分脑区。三者在结构上恰好处于额顶颞三区的交汇处。其中后部颞上沟紧邻颞顶连接区,而前部顶内沟和前运动皮层则毗邻运动皮层。实验和元分析均证实三者主要负责社会行为的知觉编码,涉及行为的模式识别与背景分析。颞上沟分布有多种感觉神经元,负责接受并整合各种感觉信息,其后部对行为意图尤为敏感;而前部顶内沟和前运动皮层则被视作镜像系统的核心组成,二者接受后部颞上沟传递来的信息,负责对行为模式及其意图加以识别(Van Overwalle & Baetens,2009)。元分析显示,镜像系统呈现由后向前的工作模式:行为信息经感觉输入,先由后部颞上沟进行知觉识别,再由前部顶内沟分析“客体-客体”及“客体-背景”间的联系,最后传递至前运动皮层,与已有动作模板进行模式匹配,最终确定目的意图(Van Overwalle & Baetens,2009)。
2.2.2镜像系统与自发特质推理的行为编码
已有充分证据显示镜像系统参与了两类特质推理过程(表1)。但不同特质推理任务下镜像系统的工作模式却存在差异。例如,当特质推理结果与事实不符时,负责行为模式知觉的左侧后部颞上沟和右侧前运动皮层明显偏向有意特质推理,而负责背景分析的右侧前部顶内沟却在自发特质推理中更为活跃(Ma et al.,2012)。综上所述,镜像系统应该是自发特质推理首阶段行为知觉编码及特质激活的主要承担者(Ma,Vandekerckhove,Van Overwalle,Seurinck,& Fias,2010)。
但镜像系统并不是特质激活的唯一承担者,实际心理化系统也会参与对社会行为的知觉编码。元分析发现,二者在行为知觉中近似处于拮抗状态——若个体行为同已有经验存在冲突,那么镜像系统活动抑制,但心理化系统反而活跃。原因在于,出现异常行为时,心理化系统会投入更多资源去解释行为。此时心理活动超越了对行为的知觉识别,进而转向了对异常行为的理解与关注。尽管如此,元分析却显示镜像系统并不会向心理化系统提供支持。对此的解释是,二者可能存有加工深度上的功能分工,镜像系统只负责较低层次的行为识别,而心理化系统则是在深层次上分析行为的意义和背景(Van Overwalle & Baetens,2009)。
2.3心理化系统与特质表征
2.3.1心理化系统的构成及其认知特性
心理化系统包括内侧前额皮层、颞顶连接区及楔叶一部;前二者作为核心心理化脑区(core mentalizing areas),在特质表征与推理中起关键作用(Ma et al.,2012)。颞顶连接区位于枕叶、颞叶与顶叶交汇处,紧挨与语义记忆有关的缘上回(supramarginal gyrus),毗邻枕部高级视觉皮层及颞叶听觉中枢,邻近负责语义理解的角回(Wernicke区),属多感觉通道交汇整合的关键区。据前人研究结果,可将颞顶连接区功能总结为三点:一是作为具身(embodiment)认知中枢参与自我参照加工,负责从主客体上区分自我与他人;二是作为社会信息的表征中枢,对行为意图加以分析,并参与特质、信念、共情及道德等高级社会认知加工;三是作为意图的自动编码中枢,参与刺激驱动的注意选择及内隐认知(Hahn,Ross,& Stein,2006;Van Overwalle,2009;Van Overwalle & Baetens,2009)。
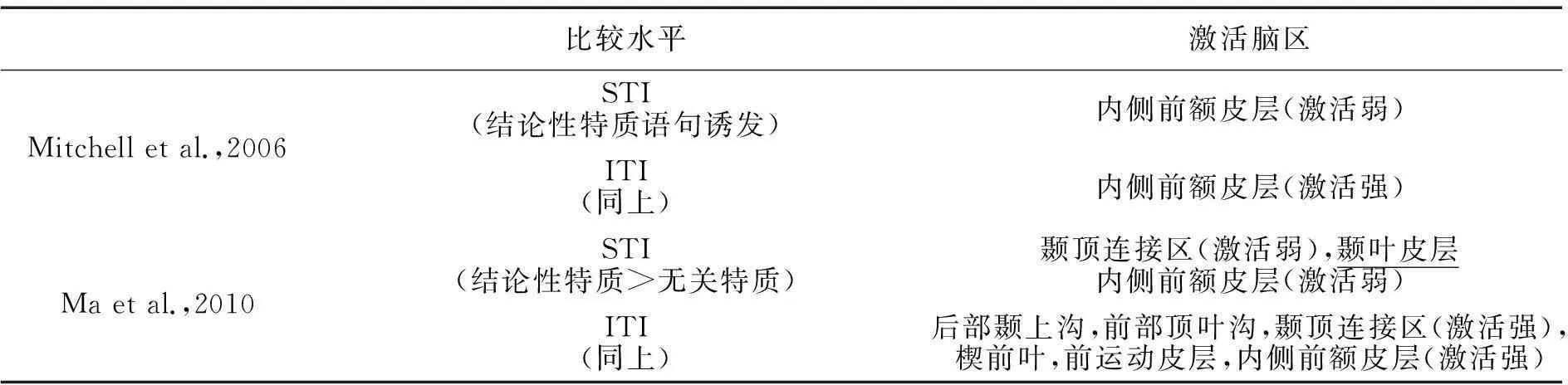
表1 fMRI研究中自发特质推理较有意特质推理诱发的激活脑区
续表1
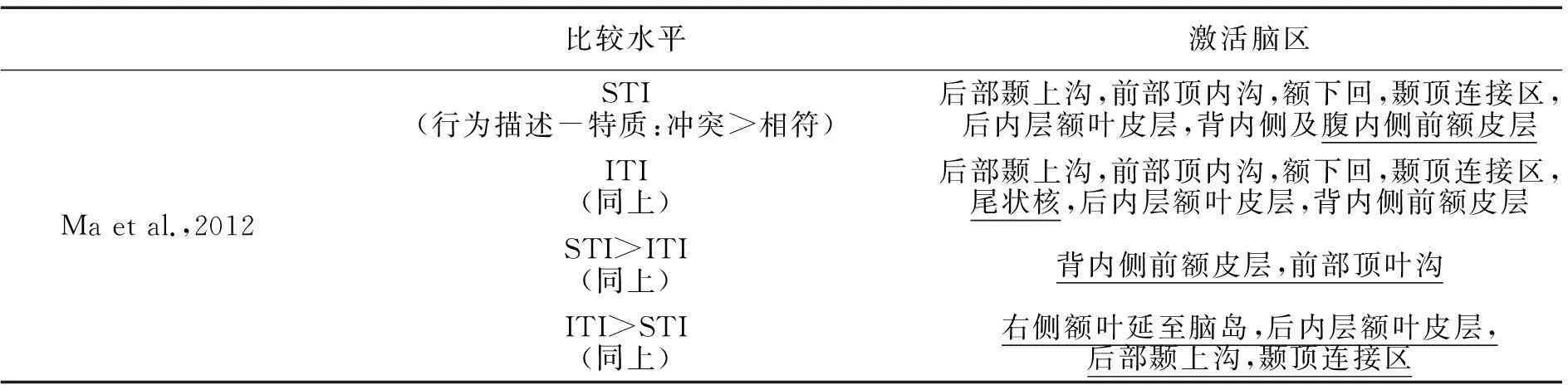
比较水平激活脑区Maetal.,2012STI(行为描述-特质:冲突>相符)后部颞上沟,前部顶内沟,额下回,颞顶连接区,后内层额叶皮层,背内侧及腹内侧前额皮层ITI(同上)后部颞上沟,前部顶内沟,额下回,颞顶连接区,尾状核,后内层额叶皮层,背内侧前额皮层STI>ITI(同上)背内侧前额皮层,前部顶叶沟ITI>STI(同上)右侧额叶延至脑岛,后内层额叶皮层,后部颞上沟,颞顶连接区
注:括号标注了比较的具体条件,如(行为描述-特质:冲突>相符)表示行为描述与特质冲突与相符两种条件的相互比较。下划线显示该条件下的特异性脑区。
内侧前额皮层则涵盖了自后部额叶皮层(pMFC)向前,从喙后部内侧额叶皮层(prMFC)至眶部内侧额皮层(oMFC)间,含扣带回的广大额叶皮质。该脑区涉及的功能异常复杂,总体可归为四点:一是作为自我和心理化中枢,直接参与印象形成和特质推理,负责自我参照加工及对他人特质和信念的分析;二是与注意系统及中枢控制系统协作,综合进行信息筛选、认知监控和行为抑制;三是与位于外侧前额的工作记忆系统和语义记忆系统联系,参与快速记忆检索,并就输入和提取的信息做深入整合与精细评估;四是接受边缘系统及眶额皮层的情绪输入,在认知判断中融入情绪信息(Amodio & Frith,2006;Van Overwalle,2009)。
2.3.2心理化系统与特质表征
已有大量证据显示,颞顶连接区参与了两类特质推理下的特质表征过程(表1和表2)。ERP溯源发现,该脑区激活可出现在关键行为特征呈现后的N100阶段,显示快速自动化的特征。此外它对外显和内隐的行为意图均十分敏感,承担对他人心理状态的快速知觉分析工作(Van Duynslaeger,Van Overwalle,& Verstraeten,2007;Van Overwalle,Van den Eede,Baetens,& Vandekerckhove,2009)。
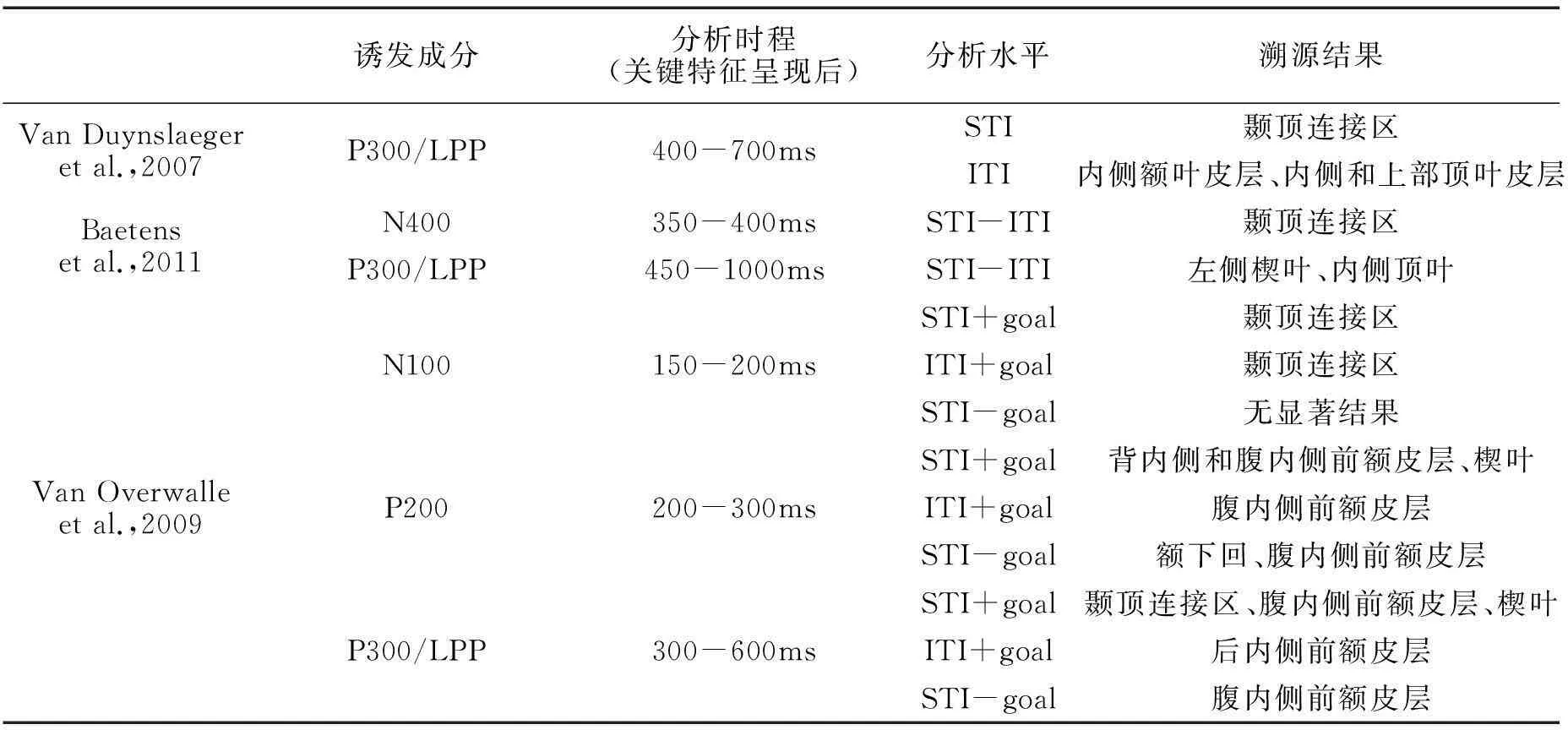
表2 两类特质推理诱发的ERP成分及其溯源结果
注:STI-ITI表示两种特质推理比较;+goal表示推理信息带有内在意图指向,-goal则表示排除意图指向后的条件水平。内侧前额皮层则是特质表征及印象形成的神经中枢。依据结构功能的差异,内侧前额皮层可分为背内侧和腹内侧两部分。背内侧前额皮层主要参与对陌生他人的特质加工,与印象形成及其改变密切相关(Gilron & Gutchess,2012);而腹内侧前额皮层则对自我或熟悉对象的特质敏感,常参与自我参照加工,并在特质编码中呈现特殊的适应性效应——当使用与对象特征相符、相反或无关的语义描述来重复启动特质推理时,腹内侧前额皮层会变得越发敏感。此外,相对背内侧前额皮层的有意精细编码,腹内侧前额皮层的特质编码呈现内隐自动化的特性,更易出现在自发特质推理中(Ma,Baetens,Vandekerckhove,Kestemont,Fias,& Van Overwalle,2014)。
为了考察核心心理化脑区在两类特质推理下的功能差异,Ma等(2012)采用fMRI技术检验了其在加工相符、不相符及无关联三类语义组合时的编码特性。研究发现:(1)两类特质推理均依赖颞顶连接区与内侧前额皮层的参与。(2)相对行为描述与无关特质的组合,当行为描述与结论特质相符时,背内侧额叶皮层在有意特质推理中的激活显著强于自发特质推理,而颞顶连接区的激活模式却与之相反。(3)当行为描述与结论特质相矛盾时,左侧颞顶连接区和内侧前额皮层在有意特质推理下的激活较自发特质推理更强烈;此外负责冲突监控的认知控制系统,如外侧额叶皮层(LPFC)、后部内侧额叶皮层(pMPFC)、扣带回(ACC)等,也会参与两类特质推理过程,显示心理化系统在特质推理中同时受意识状态以及行为与特质间编码一致性的影响。
2.3.3心理化系统在特质表征与推理中的动态过程
借助ERP高时间分辨率的优势,心理化系统在特质表征中的动态特征也得以揭示。早先Van Duynslaeger等(2007)发现,有意和自发特质推理的过程差异主要体现为关键行为特征诱发的P300分离效应,溯源结果则显示颞顶连结区与内侧额叶皮层的激活程度差异是其产生的原因。以此为基础,Van Overwalle等(2009)深入分析了自发特质推理的动态过程。该研究采用2(目的 vs.目的+特质)×2(有意 vs.无意)设计,以指导语提供任务目的,引导被试完成有意目的推理、自发目的推理、有意特质推理及自发特质推理四种任务。实验仍关注关键行为特征诱发的ERP效应。实验发现,四种推理均未在N100阶段发生过程变异。但在P200和P300阶段,自发特质推理较其他任务出现过程变异。溯源分析显示:(1)N100阶段,带有目的四类推理任务都显著激活了颞顶连接区和楔叶。其中自发特质推理和自发目标推理下的右侧颞顶连接区激活更强,而有意特质推理和有意目标推理下的左侧颞顶连接区激活更强,但上述差别均未达到显著水平。(2)至P200阶段,自发特质推理下颞顶连接区和楔叶激活消失,但其他任务下二者却持续激活至P300阶段。同时,有意特质推理和自发特质推理却额外激活了腹内侧及后内侧前额皮层,且此二者在自发特质推理下尤为活跃。(3)至P300阶段,有意特质推理和自发特质推理的差异仍体现在腹内侧前额皮层上,但已消失的颞顶连接区激活却在自发特质推理下重新活跃,并以左侧的激活程度更为强烈。
综上所述,自发特质推理至少包含两个加工阶段:阶段一负责对目标行为进行初级特征分析并展开意图的自动觉察,该任务主要由颞顶连接区承担,代表成分为N100。由于所处阶段早且颞顶连接区只具有低级知觉功能,因此不涉及抽象特质的高级编码整合。阶段二则是核心的特质表征阶段,也是自发特质推理较其它推理加工产生过程分离的阶段,代表成分为P200和P300,内容涉及内侧前额皮层对特质的精细编码以及以它为基础的特质判断。在此阶段,虽然有意和自发特质推理下的特质表征都需要内侧额叶皮层的参与,但二者仍然存在差异——前者更依赖具有有意精细加工特性的背内侧前额皮层,而后者则偏好具有内隐自动特性的颞顶连接区及腹内侧前额皮层(Van Overwalle et al.,2009)。
2.3.4核心心理化系统与特质行为的记忆提取
除参与特质表征外,核心心理化系统在行为特质间的有意回忆中也起着关键作用。研究显示,在特质推理后进行行为描述的有意回忆,回忆的正确率与颞顶连接区和内侧前额皮层的神经活动存在高度相关。Van Duynslaeger等(2007)发现,当线索特质与行为描述相符时,不管是自发特质推理还是有意特质推理,均出现回忆效果与以上脑区激活程度的显著相关。但当线索特质与行为描述项目不相符或改用句子补全任务时,以上效应消失。可见,核心心理化系统能够借助行为与特质间编码的关联性,通过线索回忆来提高对行为项目的回忆效果。但若线索发生改变,行为与特质间编码不符或是变成内隐记忆编码,则记忆促进效应消失。该效应进一步证明核心心理化系统是特质归因的主要承担者,负责特质的精细工作。
2.4心理化系统与特质判断
过程理论所定义的特质影响实质是个体基于特质与行为间内在联系做出判断的过程,它也是自发特质推理连续特质加工的最后一环。但在本质上,特质判断并不是理性的思维推理过程,而是具备“资源吝啬鬼”特性并带有知觉偏差的社会判断过程。特质的“推理”译自inference,实指一种将他人行为特征表征心理化并以之为判断基础的社会认知过程,是心理理论的具体体现。而思维的“推理”则译自reasoning,指的是基于若干已知条件判断推导出未知结论的形式逻辑过程。与思维推理相比,自发特质推理兼具社会认知非理性的特征,它建立在知觉主体的已有知识经验、图式信念乃至人格倾向基础上,受社会信念和期望影响较大,且常为刻板印象与偏见表达的过程载体(Uleman et al.,2012)。
研究显示,内侧前额皮层是特质推理的主要承担者,而思维推理的脑中枢也被证明是内侧前额皮层。为了区别二者,近期有元分析比较了特质推理同思维推理在内侧前额皮层上的机制差异(Hartwright,Apperly,& Hansen,2014;Van Overwalle,2011)。结果显示,思维推理除激活内侧前额皮层外,还激活了负责冲突监控的后内侧前额皮层、背侧扣带回及外侧前额皮层;而特质推理除激活内侧前额皮层外,仅激活了颞顶连接区。此外,虽然特质推理和思维推理可同时激活内侧前额皮层,但当思维推理偏向非社会性信息时,内侧前额皮层的激活却被明显削弱。可见,自发特质推理中的特质影响是不同于形式逻辑的社会认知过程,此推理(inference)非彼推理(reasoning)。
3镜像系统与心理化系统的功能表达:基于自发特质推理语义编码的解析
在模型构成上,Van Overwalle的社会脑模型是对镜像神经元理论的深化与发展,它仅由镜像系统与心理化系统构成,并不包含其他认知神经系统。但在功能实现上,镜像系统与心理化系统却并非孤立;相反,二者的认知加工并不排斥其他认知神经系统的参与。自发特质推理所显示的语义特性即证明了二者同语义记忆系统的协作关系。
3.1自发特质推理中的语义编码
特质的语义表征和记忆均离不开语义记忆系统的支持。行为研究证明,儿童特质推理能力同其语言发展及年龄经验存在显著相关(王美芳,陈会昌,2009)。而在神经层面,自发特质推理诱发的语义N400效应则能显示镜像系统、心理化系统同语义记忆系统间的紧密关联。
图2 N400效应产生的神经模型(Lau,Phillips,& Poeppel,2008)
语义N400效应多出现在语义冲突背景下,其形成与语义记忆提取相关,心理意义代表的是语义评估阶段出现的语义期望违背。元分析显示它产生自多个脑区构建的动态神经网络(图2),涉及额颞皮层的左侧额下回(Broca区)、内侧颞叶皮层、前颞叶皮层、颞上沟及相邻的角回(Wernicke区)和颞上回。而以上脑区多与颞顶连接区存在神经联络。因此仅从神经结构看,语义记忆系统就与心理化系统存有天然联系(Lau et al.,2008)。
自发特质推理的N400效应通常出现在与行为描述矛盾的特质判断下,并可用词语判断任务诱发:先配对呈现对象面孔和行为描述,要求被试学习,待学习结束后进行词语判断任务。词语判断先呈现1500ms的面孔,之后呈现200ms真词(特质词)或假词,其中真词含有与行为项目相符或冲突两种条件,要求被试忽略面孔做出真词/假词判断。倘若出现的真词是与对象行为描述相冲突的反义词,即能诱发语义N400效应(Varnum,Na,Murata,& Kitayama,2012)。鉴于特质与行为间的内在联系,该效应显然反映了自发特质推理诱发的语义期望违背。溯源分析发现,这一特殊的N400效应产生自颞上回。该脑区紧挨负责语义理解的Wernicke区及负责社会信息表征的颞上沟和颞顶连接区,显示自发特质推理下存在镜像系统、心理化系统与语义记忆系统的协同加工(Baetens,der Cruyssen,Achtziger,Vandekerckhove,& Van Overwalle,2011)。
3.2文化与社会因素对自发特质推理的影响
更重要的是,自发特质推理是一种后天获得的社会能力,会受文化背景、家庭经济与教育环境的显著影响。Varnum等人(2012)证明,个体所属的社会阶层状况能够影响并预测其自发特质推理能力的发展,证据就源自显示特质语义编码的N400效应。该研究开展于代表美国精英教育的密歇根大学,参加的大学生被试选自工薪阶层(父母仅接受过中等教育)和中产阶级(父母接受过高等教育)两类家庭,实验任务即为前文的词语判断任务。实验发现,自发特质推理诱发的N400效应与个体所属的家庭阶层情况有着密切联系。当被试在学习完面孔与特征描述的组合后,以面孔作为启动线索,将其与特征描述的反义词继时呈现,属中产阶级家庭的个体能够诱发显著N400效应,但属工薪阶层家庭的个体却未出现类似效应。
此外,文化差异及其衍生的人格自我差异也会反映在代表特质语义加工的N400效应上,凸显自发特质推理的跨文化差异。研究发现,相对东方集体主义文化,西方个人主义文化下的个体通常拥有更为独立的人格自我,更易忽略环境背景做出自发特质推理,由此在面对失匹配的“行为—特质”联结时诱发更强的N400效应(Na & Kitayama,2011)。鉴于这种文化特殊性,未来研究探讨镜像系统与心理化系统的认知功能时,不可忽视社会文化以及人格因素的影响。
4认知控制系统对镜像系统和心理化系统的干预
4.1自发特质推理中的认知控制
在语义记忆系统外,中枢认知控制系统及参与认知监控的工作记忆也被认为参与了镜像系统与心理化系统的功能表达。如前文所述,有意和自发特质推理均会受行为与特质间语义一致性的影响。当行为描述与特质语义相违背时,隶属认知控制系统的外侧额叶皮层、后部内侧额叶皮层及扣带回等脑区会干预特质推理过程,这是两类特质推理的共同之处。但功能连接分析却显示两类特质推理下的中枢认知控制仍有较大差异。Ma等(2012)发现自发特质推理中认知控制系统的外侧额叶皮层和后内侧额叶皮层同心理化系统的背内侧前额皮层、左侧颞顶连接区及镜像系统的前运动皮层存在功能连接。但有意特质推理下却未发现类似效应。这说明认知控制系统、心理化系统及镜像系统在自发特质推理下存有动态功能协作,三者共同参与了对异常行为的知觉表征与特质判断;而有意特质推理下,认知控制系统可能仅是独自对语义特质违背进行评估与抑制。
4.2工作记忆对自发特质推理的影响
工作记忆是认知控制开展的重要资源,也是信息进入中枢加工的关键通道。对于认知控制系统的功能分工,现有观点认为外侧额叶皮层与认知冲突下工作记忆负荷的增大有关,而后内侧额叶皮层则与认知冲突的自动觉察有关。因此外侧额叶皮层在自发特质推理中的激活可视为工作记忆的卷入(Ma et al.,2012)。
另一方面,研究发现先验性的目的意图会影响甚至阻碍自发特质推理,原因在于这些目的意图会占用工作记忆资源,进而提高特质表征及判断的认知负荷(Wells,Skowronski,Crawford,Scherer,& Carlston,2011)。而工作记忆负荷的增大则会影响中枢认知监控,进而改变个体特质推理的加工倾向。例如高认知负荷任务下,个体会倾向于采用带有信念期望且具有语义启动特性的刻板印象进行自发特质推理;而低认知负荷下,自发特质推理则会消失,由有意特质推理取代(Ferreira et al.,2012)。
5小结
Van Overwalle的社会脑模型从神经系统的功能差异着眼,将镜像神经元系统分离为负责社会知觉的镜像系统和负责社会认知的心理化系统,进而解释从行为知觉编码至特质表征判断的连续动态过程。在理论构建上,该模型吸收了模拟论有关心智理解的理论长处,同时借鉴了心理理论有关信念、特质的理论优势。相较镜像神经元研究对模拟论的支持印证,传统心理理论近年来之所以衰落,其重要原因就是缺乏脑科学证据的支持。而“镜像-心理化系统”的提出与自发特质推理的有关发现,实质将模拟论与心理理论统一在相同框架下,因此具有重大的理论价值。
而从理论发展角度看,以下问题值得深入思考:(1)多系统间的因果功能连接问题。现有研究并未完全澄清镜像系统和心理化系统同其他功能系统间的动态因果连接。因此,未来可考虑ERP-fMRI融合、动态因果建模等手段来建构动态功能连接模型。(2)心理化系统的楔叶及楔前叶功能作用问题。研究显示,作为信息重要节点的楔叶及楔前叶也参与了自发特质推理,但二者的功能仍属研究空白,未来研究可对此进行探讨。(3)左右颞顶连接区的功能差异。尽管诱发刺激多为语义材料,但自发特质推理却常出现右脑加工优势,具体表现为初期右侧颞顶连接区的强烈激活,之后则迅速转为左侧激活(Van Duynslaeger et al.,2007;Van Overwalle et al.,2009)。右侧颞顶连接区一般对他人的心理状态敏感,且被视为刺激驱动性注意的中枢;而左侧颞顶连接区在某些心理状态推断任务下甚至不会激活,但它却被证明在信念理解中不可或缺(Samson,Apperly,Chiavarino,& Humphreys,2004)。这种左右颞顶连接区转化很可能体现了由心理状态推测向高级信念加工的转变,值得未来研究予以验证。
参考文献
王美芳,陈会昌.(2009).儿童行为与心理水平的特质推理的发展.心理学报,47(10),947-957.
Baetens,K.,der Cruyssen,L.V.,Achtziger,A.,Vandekerckhove,M.,& Van Overwalle,F.(2011).N400 and LPP in spontaneous trait inferences.BrainResearch,1418,83-92.
Bargh,J.A.,Schwader,K.L.,Hailey,S.E.,Dyer,R.L.,& Boothby,E.J.(2012).Automaticity in social-cognitive processes.TrendsinCognitiveSciences,16,593-605.
Ferreira,M.B.,Garcia-Marques,L.,Hamilton,D.,Ramos,T.,Uleman,J.S.,& Jerónimo,R.(2012).On the relation between spontaneous trait inferences and intentional inferences:An inference monitoring hypothesis.JournalofExperimentalSocialPsychology,48,1-12.
Gilron,R.,& Gutchess,A.H.(2012).Remembering first impressions:Effects of intentionality and diagnosticity on subsequent memory.Cognitive,Affective,&BehavioralNeuroscience,12,85-98.
Hahn,B.,Ross,J.T.,& Stein,A.E.(2006).Neuroanatomical dissociation between bottom-up and top-down processes of visuospatial selective attention.Neuroimage,32(2),842-853.
Hartwright,C.E.,Apperly,I.A.,& Hansen,P.C.(2014).Representation,control,or reasoning?Distinct functions for theory of mind within the medial prefrontal cortex.JournalofCognitiveNeuroscience,26,685-698.
Lau,E.F.,Phillips,C.,& Poeppel,D.(2008).A cortical network for semantics:[De]constructing the N400.NatureReviewsNeuroscience,9(12),920-933.
Ma,N.,Baetens,K.,Vandekerckhove,M.,Kestemont,J.,Fias,W.,& Van Overwalle,F.(2014).Traits are represented in the medial Prefrontal Cortex:An fMRI adaptation study.SocialCognitiveandAffectiveNeuroscience,9(8),1185-1192.
Ma,N.,Vandekerckhove,M.,Baetens,K.,Van Overwalle,F.,Seurinck,R.,& Fias,W.(2012).Inconsistencies in spontaneous and intentional trait inferences.SocialCognitiveandAffectiveNeuroscience,7(8),937-950.
Ma,N.,Vandekerckhove,M.,Van Overwalle,F.,Seurinck,R.,& Fias,W.(2010).Spontaneous and intentional trait inferences recruit a common mentalizing network to a different degree:Spontaneous inferences activate only its core areas.SocialNeuroscience,6,123-138.
Na,J.,& Kitayama,S.(2011).Spontaneous trait inference is culture-specific behavioral and neural evidence.PsychologicalScience,22(8),1025-1032.
Ochsner,K.N.,& Lieberman,M.D.(2001).The emergence of social cognitive neuroscience.TheAmericanPsychologist,56(9),717-734.
Samson,D.,Apperly,I.A.,Chiavarino,C.,& Humphreys,G.W.(2004).Left temporoparietal junction is necessary for representing someone else’s belief.NatureNeuroscience,7,499-500.
Skowronski,J.J.,Carlston,D.E.,Mae,L.,& Crawford,M.T.(1998).Spontaneous trait transference:Communicators take on the qualities they describe in others.JournalofPersonalityandSocialPsychology,74(4),837-848.
Uleman,J.S.,Rim,S.,Adil Saribay,S.,& Kressel,L.M.(2012).Controversies,questions,and prospects for spontaneous social inferences.SocialandPersonalityPsychologyCompass,6(9),657-673.
Uleman,J.S.,Adil Saribay,S.,& Gonzalez,C.M.(2008).Spontaneous inferences,implicit impressions,and implicit theories.AnnualReviewofPsychology,59,329-360.
Van Duynslaeger,M.,Van Overwalle,F.,& Verstraeten,E.(2007).Electrophysiological time course and brain areas of spontaneous and intentional trait inferences.SocialCognitiveandAffectiveNeuroscience,2,174-188.
Van Overwalle,F.(2009).Social cognition and the brain:A meta-analysis.HumanBrainMapping,30,829-858.
Van Overwalle,F.(2011).A dissociation between social mentalizing and general reasoning.Neuroimage,54,1589-1599.
Van Overwalle,F.,& Baetens,K.(2009).Understanding others’ actions and goals by mirror and mentalizing systems:A meta-analysis.NeuroImage,48,564-584.
Van Overwalle,F.,Van den Eede,S.,Baetens,K.,& Vandekerckhove,M.(2009).Trait inferences in goal-directed behavior:ERP timing and localization under spontaneous and intentional processing.SocialCognitiveandAffectiveNeuroscience,4,177-190.
Varnum,M.E.W.,Na,J.,Murata,A.,& Kitayama,S.(2012).Social class differences in N400 indicate differences in spontaneous trait inference.JournalofExperimentalPsychology:General,141,518-526.
Wells,B.M.,Skowronski,J.J.,Crawford,M.T.,Scherer,C.R.,& Carlston,D.E.(2011).Inference making and linking both require thinking:Spontaneous trait inference and spontaneous trait transference both rely on working memory capacity.JournalofExperimentalSocialPsychology,47,1116-1126.
The Neuroscience Bases of the Spontaneous Trait Inferences:
A Model Structured by Mirror and Mentalizing Systems
Jia Lei1,Zhang Changjie1,Zhang Qinglin2
(1.Department of Psychology,Zhejiang University of Technology,Hangzhou 310014;
2.Faculty of Psychology,Southwest University,Chongqing 400715)
Abstract:The spontaneous trait inference refers to an automatic social cognitive processing which infers others’ traits from behavior descriptions without specific purpose or under any awareness.It has intimate relationship with the mind of theory(ToM)and preconscious automaticity.This research reviewed the neuroscience bases of the spontaneous trait inferences based on the Mirror-Mentalizing System model suggested by Van Overwalle et al.(2009).The roles of Mirror and Mentalizing Systems in behavior coding and trait representation were discussed,and their associations with the sematic memory network and center control system were also analyzed.
Key words:spontaneous and intentional trait inference(STI & ITI);Mentalizing System;Mirror System
中图分类号:B8409
文献标识码:A
文章编号:1003-5184(2015)05-0393-07
通讯作者:张庆林,E-mail:zhangql@swu.edu.cn。
基金项目:*国家自然科学基金(31400900)项目资助。
猜你喜欢
当代党员(2020年20期)2020-11-06
养生月刊(2020年10期)2020-10-14
影像研究与医学应用(2020年17期)2020-08-20
小康(2018年23期)2018-08-23
读者(2016年18期)2016-08-23
小康(2015年4期)2015-03-31
中国医学科学院学报(2013年3期)2013-03-11
意林(2009年8期)2009-02-11
cookie world(2008年11期)2008-12-12
