
濒危植物天目铁木和羊角槭的光合及蒸腾特性日动态比较
2015-02-24 11:16:05王晓燕杨淑贞赵明水崔敏燕李媛媛
华东师范大学学报(自然科学版) 2015年2期
关键词:光合作用
王晓燕, 杨淑贞, 赵明水, 罗 远, 崔敏燕, 李媛媛
(1. 华东师范大学 生态与环境科学学院,浙江天童国家森林生态系统野外观测研究站,上海 200241;
2. 浙江天目山国家级自然保护区管理局,浙江临安 311311;
3. 台州学院 生命科学学院,浙江台州 318000)
濒危植物天目铁木和羊角槭的光合及蒸腾特性日动态比较
王晓燕1, 3,杨淑贞2,赵明水2,罗远2,崔敏燕1,李媛媛1
(1. 华东师范大学 生态与环境科学学院,浙江天童国家森林生态系统野外观测研究站,上海200241;
2. 浙江天目山国家级自然保护区管理局,浙江临安311311;
3. 台州学院 生命科学学院,浙江台州318000)
摘要:分别对浙江西天目山濒危植物天目铁木(Ostrya rehderiana)和羊角槭(Acer yangjuechi)进行叶片气体交换参数日动态测定,探讨其光合与蒸腾作用的影响因素,比较光能利用效率和水分利用效率.结果显示,天目铁木的净光合速率日动态有午休现象;当光合有效辐射没有显著差异时,天目铁木的净光合速率显著大于羊角槭(p<0.05),与其林冠层阳生环境相适应;二者光能利用效率日变化无明显差异,但弱光条件下,羊角槭的光能利用效率高于天目铁木(p<0.05),与其林下层耐阴植物的特性相符;二者的水分利用效率日动态不同,羊角槭高于天目铁木(p<0.05),表明羊角槭蒸腾活动降低而导致.
关键词:天目铁木;羊角槭;光合作用;蒸腾;日动态

第一作者:王晓燕,女,博士,研究方向为分子生态学. Email: wxy3470117@163.com.
0引言
光合作用和蒸腾作用是植物基本的生理过程,其特征可解释植物关键的生态需求[1],揭示在特定地理范围内的生长状况和耐受性[2].在自然植物群落中,处于不同高度层次的树种接受的光照强度存在差异,形成与之相适应的光合特性,主要体现在阳生和耐阴植物叶片结构和功能的分化.阳生植物和耐阴植物也因此进化出多种不同途径的光合、呼吸、蒸腾特征以最大程度的适应环境[3-6].尽管存在环境可塑性,但分析影响不同高度层植物的光合、蒸腾作用的环境特征,可为确定植物适宜的生长条件提供依据.对濒危植物而言,研究其光合和蒸腾作用及其影响因素,对于正确评估最优栖息地条件更为重要[7].在迁地保护时,还可正确地判断其生长所需最适的光照、水分等环境因素,便于更好的选择最优栽培地[8].
为制定濒危植物有效的保护措施,近年来,国内外开展了不少濒危植物光合和蒸腾特征的研究.研究表明,裂叶沙参(Adenophoralobophylla)、金花茶(Camellianitidissima)等濒危植物的光合、呼吸和蒸腾等生理代谢速率比对照种在相同条件下低[9-10],即使光合特性相似,濒危物种的光合产物也远远低于未处于濒危状态的植物[11].在不利环境中,濒危植物比近缘的非濒危植物有较高的蒸腾速率,而表现出较低的光合速率、水分利用率和生物量积累[12].但对于处在不同群落高度的阳生和耐阴濒危物种的光合、蒸腾特征少有报道.
天目铁木(OstryarehderianaChun) (2n=16[13])为桦木科(Betulaceae)铁木属(Ostrya)落叶乔木,国家一级保护植物,生于常绿落叶阔叶混交林的林冠层,目前全世界仅存5株野生植株.羊角槭(AceryangjuechiFang et P.L.Chiu)隶属槭树科(Aceraceae)槭属(Acer),为常绿落叶阔叶混交林下木层的落叶乔木或灌木种,国家二级保护植物,世界仅存2株[14].这两种野生植物仅分布于浙江省西北部临安县的西天目山.对于个体数极少的濒危植物,迁地栽培是必要的保护方法[15],已用于这两种濒危植物的保护.但是,天目铁木花粉寿命短,种子发芽率低[16],幼苗不易生长[17];羊角槭也存在种子败育率高,种子活力低等状况[18].因此,适宜的异地栽培条件对有幸存活幼苗的生长尤为重要.研究显示,耐阴植物羊角槭较阳生植物天目铁木的光补偿点和光饱和点低[19],与阴生植物的特征相符,本文选择这两种植物,分析光合及蒸腾日动态特征,探讨濒危机制,为更好地制定就地和迁地保护策略提供理论依据.
本文从叶片气体交换日动态特征方面对天目铁木和羊角槭进行分析,目的是:①比较处于林冠层天目铁木和林下层羊角槭的光合及蒸腾作用特征;②揭示影响光合及蒸腾作用的主要影响因素,为两种濒危植物的有效保护和异地栽培提供建议.
1材料与方法
1.1 研究地点和对象
研究地点位于野生天目铁木和羊角槭分布的浙江省西天目山(191°27′ E,30°17′ N).天目山区属北亚热带气候,四季分明,气候温和,雨量充沛,光照适宜.从山麓至山顶年均温为14.8~8.8°C,最冷月1月气温3.4~2.6°C,最热月7月气温28.1~19.9°C.年降水量1 390~1 870 mm,年蒸发量1 150~820 mm,年日照时数1 550~2 000 h,无霜期235~209 d,土壤随着海拔升高由亚热带红壤向温带型棕黄壤过渡[20].
已知仅存的5株野生天目铁木位于西天目山南坡海拔约250 m的山麓林缘处.其中,1株位于公路旁,树高约21 m,胸径达1 m,年龄约100 a.另外4株在西边相距约200 m处,树高高于20 m,胸径40~50 cm,生境状况基本一致.残存的2株野生羊角槭位于西天目山南坡海拔约870 m处,生于以紫楠(Phoebesheareri)、绵椆(Lithocarpusharlandii)、香果树(Emmenopteryshenryi)为优势种的常绿落叶阔叶混交林内.然而,由于冰冻雪灾的影响,此2株生长状况不良.通过种子繁殖的部分羊角槭栽植于西天目山山坡上,选择其中1株进行测量,该树木高约10 m,胸径约40 cm.
1.2 研究方法
在两种植物的生长旺盛期,选择晴朗少云天气(2008年9月1日至9月4日),从6:30~17:30,用LI-6400便携式光合作用测定系统(LI-COR Inc.,Lincoln,NE,USA)对野生天目铁木和羊角槭叶片气体交换参数和日动态特征进行测定,每1 h测定1次,两物种分别从树冠边缘选取3个枝条,每个枝条选取健康、长势良好的向阳成熟叶片3-4片进行测定,每个叶片重复读取5-7组数据取平均值.为了对天目铁木进行准确的在体测定,在树干旁边搭建约10 m的工作台;羊角槭的在体测定借助树下的墙壁和梯子.测定前对光合仪进行系统校正,测定时尽量使叶片保持自然着生角度.
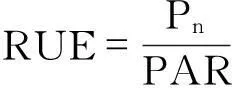
1.3 数据处理
对天目铁木和羊角槭的净光合速率(Pn)、光能利用效率(RUE)、水分利用效率(WUE)以及光合有效辐射(PAR)等参数的比较采用配对t检验,如数据不符合正态分布则采用非参数Wilcoxon配对检验.分别对Pn和蒸腾速率(Tr)与其它各指标进行相关分析及多元逐步回归分析,相关分析采用非参数Spearman相关系数.统计分析使用SPSS 17.0软件完成.
2结果
2.1 主要环境因子日变化
天目铁木的光合有效辐射(PAR)日变化呈“单峰”曲线,11:30至14:30辐射最强,羊角槭PAR的日变化呈“平台型”,8:30至13:30基本处于高峰,13:30后快速下降(见图1a).两种植物测定时全天的PAR有显著差异(p<0.05,见表1),但当以辐射最大的时刻14:00为界,之前和之后测定2种植物的PAR都无显著差异(p>0.05,见图1a).两种植物测定时的大气CO2浓度(Ca)、大气温度(Tair)、叶片温度(Tleaf)和大气相对湿度(RH)日变化趋势相似,但天目铁木测定时Ca在午间11:30有较大的升高(见图1和表1).叶片温度(Tleaf)和Tair变化趋势一致(见图1d和表1),相关分析表明二者呈极显著正相关(p<0.01),说明Tleaf主要随气温而变化,RH总体上清晨较高.

注:虚线表示以最大光合有效辐射的时刻(14:00)为界前后分别比较;p>0.05无显著差异,p<0.05

表1 天目铁木和羊角槭主要环境因子日均值
注:平均值±标准差Means±S.D.;PAR,光合有效辐射,Ca,大气CO2浓度,Tair:大气温度;Tleaf,叶片温度;RH,大气相对湿度;字母不同代表有显著差异(p<0.05)
2.2 两种植物光合作用特征比较
两物种净光合速率(Pn)日变化格局明显不同(见图2a和表1),天目铁木呈“双峰”曲线. 11:30和14:30的 Pn分别达到7.49和6.85 μmol·m-2·s-1,期间12:30有明显的光合“午休”现象,Pn降为5.58 μmol·m-2·s-1.羊角槭的Pn日变化则呈现“平台型”曲线,8:30至13:30都处于平台期 (4.66~5.50 μmol·m-2·s-1).净光合速率与环境因子的相关分析表明(见表3):两个物种的Pn都与PAR呈极显著正相关,PAR是影响光合作用的主要因子.此外,羊角槭的Pn还与Tair呈显著正相关,与Ca呈显著负相关,但逐步回归分析表明这两种环境因子不是主要影响因素.两物种PAR在13:30之前不存在显著差异(p>0.05,见图1a),但天目铁木Pn(5.69 μmol·m-2·s-1)显著高于羊角槭(4.30 μmol·m-2·s-1) (p< 0.05,见图2a).

注:虚线表示以前后参数差别大的时刻为界分别比较,p > 0.05无显著差异,p < 0.05有显著差异
导致植物Pn日变化呈早晚低,午间高的原因有气孔限制因素和非气孔限制因素[22].当两种植物的午后Pn降低时,相应的胞间CO2浓度(Ci)增大(见图2b),气孔导度(Gs)(见图2c)和气孔限制值(Ls)减小(见图2d),说明午后Pn降低是由非气孔限制因素导致的.但是,天目铁木的Pn在12:30降低的同时(见图2a),Gs下降(见图2c),Ls增大(见图2d),阻止了CO2的供应,导致Ci降低(见图2b),说明“午休”现象由气孔限制因素导致.
天目铁木和羊角槭的瞬时光能利用效率(RUE)也呈现早晚高、午间低的趋势(见图2e),两种植物日变化无显著差异(p>0.05, 见表2),但15:30以后随着光照强度减弱,羊角槭的RUE显著大于天目铁木(p<0.05,见图2e).天目铁木和羊角槭瞬时水分利用效率(WUE)都是清晨和下午较高、中午较低(见图2f),表明午间的蒸腾作用比较大,羊角槭显著大于天目铁木(p<0.05, 见表2).

表2 天目铁木和羊角槭叶片主要气体交换参数日均值
注:平均值±标准差 Means±S.D.;Pn,净光合速率;Ci,胞间CO2浓度;Gs,气孔导度;Ls,气孔限制值;RUE,光能利用效率;WUE,水分利用效率;Tr,蒸腾速率; VPD,叶面饱和蒸气压亏缺;字母不同代表有显著差异(p<0.05)
2.3 两种植物蒸腾作用特征日变化
两物种的蒸腾速率(Tr)格局不同(见图3a),天目铁木的Tr出现2个峰值,一个是清晨6:30最高,Tr为4.28 mmol·m-2·s-1,7:30突然下降后逐渐回升至13:30又达到峰值(4.32 mmol·m-2·s-1),以后又逐渐下降.羊角槭的Tr为单峰曲线,中午10:30和11:30达到最高值,分别为2.80和2.81 mmol·m-2·s-1.
蒸腾速率与环境和生理因子的相关分析表明(见表3):当去除天目铁木Tr在6:30的极高值后,两种植物的Tr与PAR、Tair呈极显著的正相关,PAR增强可提高Tair,从而使Tr加快(见图1a、1c和图3a).羊角槭的Tr还与Ca呈显著的负相关关系(见图1b和图3a),与其Pn对Ca的反应一致.两物种的Tr与湿度均无显著相关关系(见表3).生理因子中,两物种的Tr都与Gs和VPD呈显著正相关(见表3、图2c和图3b),当天目铁木和羊角槭Tr达到最高值时,VPD也分别于13:30和10:30时达最大值,说明蒸腾作用受叶面饱和蒸气压作用下气孔开闭的影响.对Tr的逐步回归也看出,PAR和Tair是影响Tr的主要环境因子.

表3 光合、蒸腾速率与环境因子和生理因子的相关系数
注:天目铁木蒸腾速率和气孔导度的相关系数计算时去除了6:30的极高值;*表示在0.05水平显著相关,**表示在0.01水平上显著相关;Pn,净光合速率;Tr,蒸腾速率;Gs,气孔导度;Ci,胞间CO2浓度;VPD,叶面饱和蒸气压亏缺;PAR,光合有效辐射;Ca,大气CO2浓度;Tair,大气温度;RH,大气相对湿度

图3 天目铁木和羊角槭蒸腾速率(Tr, a)和叶面饱和蒸气压亏缺(VPD, b)的日变化
3讨论
3.1 影响光合及蒸腾作用的主要因素
植物光合作用直接或间接地受到多种因素影响[23],但光能的吸收是相对独立的因素.天目铁木和羊角槭的Pn日变化与分别与PAR的变化趋势一致,呈现极显著相关,说明影响两种植物Pn的主要环境因子是PAR.但是,当接受的PAR没有差异时,两种植物的净光合速率却不同,天目铁木的Pn显著大于羊角槭(p<0.05).这与天目铁木处于林冠层有关,在相同光强下处于林冠层树种的光合性能往往大于林下层,有更强的合成有机物的能力[24].
天目铁木和羊角槭的Tr主要受PAR和Tair的影响,PAR的增加和Tair升高促使蒸腾作用增强.此外,Tr都与VPD呈极显著正相关,虽然大气湿度与羊角槭的蒸腾作用没有显著关系,但较低的大气湿度使叶片和大气间的蒸气压差(VPD)增大,潜在的导致较高的Tr[25,26],并且VPD是Gs的直接响应因素[27],影响蒸腾作用.因此,两种植物的蒸腾作用都最终受到植物内外蒸气压差的强烈影响.
3.2 两物种的光能利用效率
光能利用效率(RUE)能体现植物捕获光强的能力[28],两种植物瞬时RUE日平均分别为(0.014 2±0.004 5) mol·mol-1和(0.019 3±0.007 1) mol·mol-1,二者没有显著差异(p>0.05).与其它植物相比较,与长柄扁桃(AmygdaluspedunculataPall.)0.011 mol·mol-1[29]、禾雀花(Mucunabirdwoodiana)0.007 mol·mol-1[30]、以及3种针叶树种0.02 mol·mol-1[31]的瞬时日平均光能利用效率相当.
虽然在光强相同的时候,两种植物的RUE没有差异(p>0.05),但在弱光下RUE却明显不同.在下午15:30光照强度急剧减弱后,羊角槭的RUE大于天目铁木(p<0.05),表明羊角槭对弱光利用能力好,这是长期生长在林下层弱光条件下适应性的表现.叶绿体荧光参数测定也表明,天目铁木在弱光下生长受到影响而羊角槭则能正常生长[19].长期处于弱光下的植物常常通过改变叶绿体格局及结构,如叶绿体变大、叶绿素含量增加、叶绿体内基粒数目增多、基粒的类囊体排列紧密等方面,从而提高弱光下的RUE[32].而天目铁木作为林冠层种类,则对强光下的光抑制现象(photo-inhibition)有更强的抵抗作用[6, 33],表现出一定的自身保护机制.正午出现的光合“午休”现象是气孔限制因素所导致[22],此时PAR和Tair均达到一天中的最高值,天目铁木通过缩小气孔张开幅度对强光有积极的应对反应.另外,天目铁木正午蒸腾作用强烈,强烈的蒸腾有可能导致气孔保卫细胞周围的脱落酸增多,限制了气孔过度张开[34],也可能是强光下的自我调节反应.
3.3 两物种的水分利用效率
水分利用效率(WUE)与光合作用呈正比,与蒸腾作用呈反比.天目铁木日均WUE为(2.236 1±0.351 9) μmol·mmol-1,与同为桦木科的白桦(Betulaplatyphylla)日平均值相当(2.39 μmol·mmol-1)[35].羊角槭WUE为(2.929 0±0.273 5) μmol·mmol-1,与同为槭属的茶条槭(Aerginnala)的日均值(约2.5 μmol·mmol-1)[36]相差不大.但羊角槭的WUE大于天目铁木(p<0.05),表明羊角槭的蒸腾作用较天目铁木弱.天然羊角槭生长于林下层,耐阴特性进化出弱的蒸腾作用,以便将更多的水分用于光合作用,使耐阴植物能更有效地利用光能和水分资源.而生长于林冠层的天目铁木,在充足的光照下蒸腾作用较强,表现出较低的水分利用效率.
3.4 对两种濒危植物的保护建议
植物对光照需求范围是迁地保护时选择合适地点的重要条件[2].羊角槭具有耐阴特性,没有表现出对强光照的自我保护反应,目前天目山仅存的2株较大的羊角槭分布在房舍旁边,周围没有高大的树木遮挡,应采取适当措施如设置遮阳网等,防止强光的抑制作用.天目山保护区引种的其它羊角槭栽种在山谷中,具有较好的遮阳条件,较适合羊角槭生长.天然天目铁木是处于林冠层的阳生树种,对强光有一定的自我保护能力,在迁地栽植时应考虑充足的光照条件,目前天目山保护区栽种的天目铁木幼苗分布于山谷的坡面上,光照条件不甚充足,可能使得生长所需的光照条件受限.
天目铁木在清晨相对弱光和低温时有非常高的Gs和蒸腾作用,推测其可能属于夜间蒸腾植物,但这需要进一步验证,具体情况及生理意义还应深入研究.
[参考文献]
[1]ADAMEC L. Photosynthetic characteristics of the aquatic carnivorous plantAldrovandavesiculosa[J]. Aquatic Botany, 1997, 59: 297-306.
[2]ALERIC K M, KIRKMAN L K. Growth and photosynthetic responses of the federally endangered shrub,Linderamelissifolia(Lauraceae), to varied light environments [J]. American Journal of Botany, 2005, 92: 682-689.
[3]MURCHIE E H, HORTON P. Acclimation of photosynthesis to irradiance and spectral quality in British plant species: chlorophyll content, photosynthetic capacity and habitat preference [J]. Plant, Cell and Environment, 1997, 20: 438-448.
[4]NOGUCHI K, GO C S, MIYAZAWA S I, et al. Costs of protein turnover and carbohydrate export in leaves of sun and shade species [J]. Australian Journal of Plant Physiology, 2001, 28: 37-47.
[5]NEILL S O, GOUKD K S, KILMARTIN P A, et al. Antioxidant capacities of green and cyanic leaves in the sun species,Quintiniaserrata[J]. Function Plant Biology, 2002, 29: 1437-1443.
[7]SMITH M, WU Y, GREEN O. Effect of light and water-stress on photosynthesis and biomass production inBoltoniadecurrens(Asteraceae), a threatened species [J]. American Journal of Botany, 1993, 80: 859-864.
[8]SMITH M, HOUPIS J L J. Gas exchange responses of the wetland plantSchoenoplectushalliito irradiance and vapor pressure deficit [J]. Aquatic Botany, 2004, 79: 267-275.
[9]张文辉,祖元刚,刘国彬. 十种濒危植物的种群生态学特征及致危因素分析[J]. 生态学报, 2002, 22: 1512-1520.
[10]WEI X, JIANG Y S, JIANG S Y, et al. Photosynthetic characteristics of an endangered speciesCamellianitidissimaand its widespread congenerCamelliasinensis[J]. Photosynthetica, 2008, 46: 312-314.
[11]CHANG J, GE Y, LU Y J, et al. A comparison of photosynthesis in endangered and non-endangered plantsChangiumsmyrnioidesandAnthriscussylvestris[J]. Photosynthetica, 2002, 40: 445-447.
[12]GE Y, CHANG J, LI W C, et al. Effect of soil moisture on the gas exchange ofChangiumsmyrnioidesandAnthriscussylvestris[J]. Biologia Plantarum, 2003, 47: 605-608.
[13]孟爱平,何子灿,李建强,等. 桦木科两种濒危植物的染色体数目[J]. 武汉植物学研究, 2004, 22: 171-173.
[14]顾云春. 中国国家重点保护野生植物现状[J]. 中南林业调查规划, 2003, 22: 1-7.
[15]KRAUSS S L, DIXON B, DIXON K W. Rapid genetic decline in a translocated population of the endangered plantGrevilleascapigera[J]. Conservation Biology, 2002, 16: 986-994.
[16]张若蕙,张金谈,邹达明,等. 天目铁木的花及花粉形态[J]. 浙江林学院学报, 1988(5): 93-96.
[17]管康林,陶银周. 濒危树种——天目铁木的现状和繁殖[J]. 浙江林学院学报, 1988(5): 90-92.
[18]许小连,金荷仙,陈香波,等. 濒危树种羊角槭种子基本生物学特征[J]. 林业科技开发, 2012, 26(3): 46-49.
[19]钟泰林,李根有,石柏林. 3种浙江特产濒危植物气体交换特征和叶绿素荧光特性研究[J]. 上海交通大学学报:农业科学版, 2009, 27: 149-152,176.
[20]丁炳扬,李根有,傅承新,等. 天目山植物志(第一卷)[M]. 杭州: 浙江大学出版社, 2010.
[21]李扬,黄建辉. 库布齐沙漠中甘草对不同水分和养分供应的光合生理响应[J]. 植物生态学报, 2009, 33: 1112-1124.
[22]FARQUHAR G D, SHARKEY T D. Stomatal conductance and photosynthesis [J]. Annual Review of Plant Physiology, 1982, 33: 317-345.
[23]KIRSCHBAUM M U F. Direct and indirect climate change effects on photosynthesis and transpiration [J]. Plant Biology, 2004(6): 242-253.
[24]KENZO T, ICHIE T, WATANABE Y, et al. Changes in photosynthesis and leaf characteristics with tree height in five dipterocarp species in a tropical rain forest [J]. Tree Physiology, 2006, 26: 865-873.
[25]ASSMANN S M, GRANTZ D A. Stomatal response to humidity in sugarcane and soybean: effect of vapour pressure difference on the kinetics of the blue light response [J]. Plant, Cell and Environment, 1990, 13: 163-169.
[26]MOTT K A, PARKHURST D F. Stomatal responses to humidity in air and helox [J]. Plant, Cell and Environment, 1991, 14: 509-515.
[27]许大全. 光合作用测定及研究中一些值得注意的问题[J]. 植物生理学通讯, 2006, 42: 1163-1167.
[28]PENUELAS J, FILELLA I, LLUSIA J, et al. Comparative field study of spring and summer leaf gas exchange and photobiology of the Mediterranean treesQuercusilexandPhillyrealatifolia[J]. Journal of Experimental Botany, 1998, 49: 229-238.
[29]罗树伟,郭春会,张国庆,等. 沙地植物长柄扁桃光合特性研究[J]. 西北农林科技大学学报:自然科学版, 2010, 38(1): 125-132.
[30]林植芳,吴彤,孔国辉,等. 8种城市绿化攀援植物的光合作用和水分关系特性[J]. 热带亚热带植物学报, 2007, 15: 473-481.
[31]王军邦,张秀娟,韩海荣,等. 亚热带常绿人工针叶林冠层内光能利用率[J]. 生态学杂志, 2010, 29: 611-616.
[32]战吉宬. 植物弱光逆境生理研究综述[J]. 植物学通报, 2003, 20: 43-50.
[33]FIGUEROA F L, ESCASSI L, PÉREZ-RODRGUEZ E, et al. Effects of short-term irradiation on photoinhibition and accumulation of mycosporine-like amino acids in sun and shade species of the red algal genusPorphyra[J]. Journal of Photochemistry and Photobiology B: Biology, 2003, 69: 21-30.
[34]TALLMAN G. Are diurnal patterns of stomatal movement the result of alternating metabolism of endogenous guard cell ABA and accumulation of ABA delivered to the apoplast around guard cells by transpiration? [J]. Journal of Experimental Botany, 2004, 55: 1963-1976.
[35]杜琳,李永存,穆怀志,等. 四倍体与二倍体白桦的光合特性比较[J]. 东北林业大学学报, 2011, 39: 1-4.
[36]洪丽,卓丽环. 不同季节茶条槭光合特性的差异[J]. 东北林业大学学报, 2008, 36: 16-18.
(责任编辑张晶)
Comparative diurnal variations in photosynthesis and transpiration of endangered plant species,OstryarehderianaandAceryangjuechi
WANG Xiao-yan1, 3,YANG Shu-zhen2,ZHAO Ming-shui2,LUO Yuan2,
CUI Min-yan1,LI Yuan-yuan1
(1.TiantongNationalFieldObservationStationforForestEcosystem,SchoolofEcologicalandEnvironmentalSciences,
EastChinaNormalUniversity,Shanghai200241,China;
2.ManagementBureauofTianmushanNationalNatureReserve,Lin′anZhejiang311311,China;
3.SchoolofLifeScience,TaizhouUniversity,TaizhouZhejiang318000,China)
Abstract:In order to compare the differences of gas exchange between sun-tolerant and shade-tolerant plants, diurnal variations in photosynthetic and transpiration indices were measured using LI-6400 portable photosynthesis system in two endangered species, Ostrya rehderiana Chun (sun-tolerance) and Acer yangjuechi Fang et P. L. Chiu (shade-tolerance). Our results showed that O. rehderiana was a bimodal curve in diurnal change of photosynthesis rate (Pspan), while A. yangjuechi was a unimodal curve. A. yangjuechi had a higher light use efficiency (RUE) than O. rehderiana under a relatively weak light (p<0.05), suggesting A. yangjuechi had an increased net photosynthesis rate (Pspan) in weaker light conditions, e.g. under a long-term shade circumstance. The water use efficiency (WUE) was also higher in A. yangjuechi than in O. rehderiana (p<0.05). This indicated A. yangjuechi had a lower transpiration in shaded conditions.
Key words:Ostrya rehderiana;Acer yangjuechi;photosynthesis;transpiration;diurnal variation
通信作者:李媛媛,女,博士,副教授,研究方向为保护生物学和分子生态学. E-mail: yyli@des.ecnu.edu.cn.
基金项目:浙江省临安市科委项目
收稿日期:2014-01
中图分类号:Q16
文献标识码:A
DOI:10.3969/j.issn.1000-5641.2015.02.014
文章编号:1000-5641(2015)02-0113-09
猜你喜欢
科学(2022年4期)2022-10-25 02:43:00
发明与创新(2019年43期)2019-11-16 06:59:44
学苑创造·A版(2018年5期)2018-05-28 12:26:30
青苹果·教育研究版(2016年7期)2016-12-12 05:19:52
求学·理科版(2016年4期)2016-09-02 04:04:04
Coco薇(2016年5期)2016-06-03 09:17:41
考试周刊(2016年6期)2016-03-11 08:14:32
少儿科学周刊·儿童版(2015年10期)2015-11-07 03:45:31
高中生学习·高二版(2014年5期)2014-07-03 14:50:01
青苹果(2014年1期)2014-04-09 20:31:00
