
一氧化氮对植物重金属胁迫抗性的影响研究进展
2015-02-08 05:33夏海威施国新
生态学报 2015年10期
夏海威,施国新,黄 敏,吴 娟
南京师范大学生命科学学院 江苏省生物多样性与生物技术重点实验室, 南京 210023
一氧化氮对植物重金属胁迫抗性的影响研究进展
夏海威,施国新*,黄 敏,吴 娟
南京师范大学生命科学学院 江苏省生物多样性与生物技术重点实验室, 南京 210023
一氧化氮(NO)作为一种重要的信号分子,在调节植物重金属胁迫抗性方面上起着非常重要的作用。综述了NO在植物体内的产生途径,重金属胁迫下植物体内内源NO含量的变化以及外源NO与内源NO对植物重金属胁迫抗性的影响。大量研究表明外源NO能够增强植物对重金属胁迫的抗性,一方面是通过增强植物细胞的抗氧化系统或直接清除活性氧,另一方面是通过影响植物对重金属的吸收以及重金属在植物细胞内的分布。然而内源NO在调节植物重金属胁迫抗性上的功能角色仍存在争议。有些研究表明内源NO是有益的,能够缓解重金属胁迫诱导的毒性;但是也有证据表明内源NO是有害的,能够通过促进植物对重金属的吸收以及对植物螯合素进行S-亚硝基化弱化其解毒功能,从而参与重金属诱导的毒害反应和细胞凋亡过程。
外源NO; 内源NO; 重金属胁迫; 抗性
一氧化氮(Nitric Oxide, NO)是一种广泛存在于生物体内的水溶性和脂溶性气体小分子信号物质,在植物体内参与多种生理过程,如诱导种子萌发[1]、抑制种子休眠[2], 调节植物光合作用[3]、花的形成[4]以及植物的根向地性生长[5],延缓植物的衰老过程[6]等。同时,NO也参与调节植物对一系列非生物和生物胁迫的抗性,在非生物胁迫抗性方面,NO能够增强植物抗盐性,抗旱性,抗涝性,抗极端温度,抗紫外线辐等[7];在生物胁迫抗性方面,NO能通过增加3′5′-环-磷酸鸟苷(cGMP)和水杨酸(SA)的水平来增强植物的抗病性[8]。
对重金属胁迫抗性而言,已有大量文献表明NO能够调节植物对重金属胁迫抗性,但对NO在植物重金属胁迫抗性方面的具体机制做系统总结和概括的研究较少并且对内源NO在调节植物重金属胁迫抗性上的功能角色也很少涉及。本文结合国内外最新的研究进展,综述了NO调节植物重金属胁迫抗性的相关机制,为研究植物重金属胁迫抗性机制提供了参考。
1 植物体内NO的产生途径
1.1 酶促反应途径
1.1.1 一氧化氮合酶(NOS)途径
在动物体内,NO主要通过NOS以L-精氨酸、O2及NADPH为底物催化而成,FAD、FMN、血红素、四氢叶酸、Ca2+/CaM和Zn2+为NOS的辅基[9]。在植物体内也有类似的NOS,Neill等[7]发现植物能够通过氧化L-精氨酸形成瓜氨酸而产生NO,并且动物NOS抑制剂L-硝基精氨酸甲酯(L-NAME)能够抑制拟南芥NOS活性并减少NO的产生[10],另外人们已经在植物组织如豌豆的根、茎、叶中[11]以及各种细胞器如过氧化物酶体[12]和叶绿体[13]中都检测到了NOS活性。然而植物NOS的基因和蛋白序列与动物却并不相同,如最近发现的绿藻(Ostreococcustauri)中的NOS蛋白序列已经被鉴定出来,结果其与动物的NOS蛋白序列仅有45%的相似性,而其结构模型与动物的反应区域却高等相似[14],另外Guo等发现拟南芥中AtNOS1基因编码的蛋白与蜗牛中参与NO合成的蛋白有相似序列,但这一蛋白与典型的动物NOS蛋白序列没有相似性[15],并且体外重组的AtNOS1也没有NOS活性[16],实际上AtNOS1是一种GTPase而不是NOS,因而命名为AtNOA1[17],该蛋白可能参与线粒体核糖体的生物合成以及翻译过程,并在此过程中间接参与NO的合成[16]。
1.1.2 硝酸还原酶(NR)和亚硝酸还原酶(Ni-NOR)途径
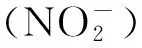
1.1.3 其他酶促途径
另外,植物体内还有其他的酶也参与NO的产生,如辣根过氧化物酶[21]、黄嘌呤氧化酶(XOR)以及黄嘌呤脱氢酶(XDH)[22]、细胞色素P450[23]等。
1.2 非酶促途径
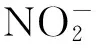
2 重金属胁迫下植物体内内源NO含量的变化
重金属胁迫下,植物细胞内源NO含量往往会发生显著变化,然而内源NO含量的变化却有众多的影响因素。
如100 μmol/L的铜(Cu)、镉(Cd)、锌(Zn)短期处理24 h以后,豌豆根内的内源NO含量升高[27],而100 μmol/L的Cu、Cd、Zn长期处理14 d以后,豌豆根内的内源NO含量却下降[28],说明重金属处理时间的长短能够影响植物体内内源NO含量的变化。
Leterier等用不同浓度的砷(As)对拟南芥处理7 d以后发现,当砷浓度小于250 μmol/L时,拟南芥根内的NO含量没有增加,甚至在250 μmol/L时出现了下降;而当砷浓度超过500 μmol/L时,拟南芥根内的NO含量大幅增加,并且在500 μmol/L时达到最大[29],表明植物体内内源NO含量的变化与重金属的浓度有关。
100 μmol/L的Cd处理24 h以后,水稻根内的NO含量下降[30],而在同样的100 μmol/L的Cd处理24 h以后豌豆根内的NO却增加[27];另外,Chen等在研究大麦的两种基因型weishuobuzhi和dong17对5 μmol/L的Cd胁迫反应时发现,两种基因型大麦体内的NO含量变化不同,其中weishuobuzhi型大麦根内的NO含量在第1天达到最大值,而后随着处理时间的延长,NO含量迅速下降,而对Cd敏感的dong17型大麦根内的NO含量在第10天达到最大,另外,weishuobuzhi和dong17叶内的NO含量都在第1天达到最大[31]。这些发现说明重金属胁迫下,植物体内内源NO含量的变化与植物的种类以及基因型和植物组织类型有关。
尽管大量研究证明植物在重金属胁迫下内源NO含量的变化受植物种类与基因型,重金属浓度以及处理时间等因素的影响,然而对植物悬浮细胞而言,重金属胁迫下其内源NO含量总会增加[32- 35],出现这一现象的原因可能是植物悬浮细胞缺少细胞与细胞之间的网络调控。
3 外源NO增强植物对重金属胁迫的抗性
3.1 外源NO缓解重金属对植物细胞造成的氧化胁迫

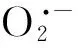
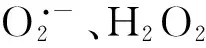
金属硫蛋白(Metallothioneins, MTs)是一种小分子的富含半胱氨酸的金属结合蛋白,通过巯基与重金属结合形成无毒或低毒络合物,从而避免有害重金属对植物体的潜在毒性[59]。在动物细胞中,有研究发现在肝细胞中添加NO的供体V-PYRRO/NO,能够大幅增强MTs基因的表达,从而减弱Cd的毒性,增强肝细胞对Cd胁迫的抗性[60]。原因可能是NO能够替换与MTs结合的重金属[61- 62],并且释放的重金属能够进一步促进MTs基因的表达[62]。在植物细胞中可能也存在类似的机制,Wang等研究发现外源NO能够提高MTs的含量并增强番茄对Cu胁迫的抗性,而对MTs敏感型番茄而言,外源NO并不能明显地增强番茄对Cu胁迫的抗性,表明MTs在介导NO缓解重金属胁迫上起着至关重要的作用[42]。
3.2 外源NO影响植物对重金属的吸收以及重金属在植物体内的分布
研究发现外源NO能够影响植物对重金属的吸收,从而调节植物对重金属胁迫的抗性。如外源NO能够减少苜蓿对Cd的吸收[47],减少小麦和豌豆对Zn的过量积累[63],抑制决明对铝(Al)的吸收[64]。但也有研究表明外源NO缓解重金属胁迫并不是通过抑制植物对重金属的吸收。如外源NO缓解过量Cu对小麦种子萌发的抑制作用,并且通过增强SOD和CAT的活性缓解过量Cu对小麦种子造成的氧化胁迫,然而外源NO并不能抑制小麦种子对Cu的吸收[65];外源NO能够缓解过量Cu对人参造成的毒害作用,但并不能显著降低人参根细胞对Cu的吸收[46];外源NO供体硝普钠(SNP)处理能够缓解铅(Pb)对拟南芥的毒害,但不能减少拟南芥对Pb的积累[66]。Xu等在研究中发现,对Cd的超富集植物龙葵而言,Cd胁迫能够诱导显著的生长抑制,促进H2O2的积累并破坏膜的完整性,而添加外源的NO供体亚硝基谷胱甘肽(GSNO)能够增强SOD和CAT的活性,增加Pro含量,抑制H2O2的积累,提高膜的完整性,从而缓解Cd对龙葵的毒害,并且添加NO清除剂c-PTIO后能够加重Cd对龙葵的毒害作用,这表明外源NO能够缓解Cd胁迫对龙葵造成的伤害,增强龙葵对Cd胁迫的抗性。但另一方面添加NO清除剂c-PTIO能够减少龙葵对Cd的吸收,添加外源的NO供体GSNO能够增加龙葵对Cd的吸收,说明NO促进龙葵根部对Cd的吸收[67],这些研究暗示了外源NO可能存在相应的调节机制调节重金属在植物体内的分布。
面对重金属胁迫,植物细胞也有一系列的防御机制,以最大程度减少重金属可能造成的伤害,其中作为抗重金属胁迫的第一道屏障,细胞壁对吸收的重金属具有束缚作用[68]。研究表明进入水稻细胞的Al 80%—85%分布在细胞壁,从而有效地阻止Al进入水稻细胞的细胞质[69],因此重金属在植物细胞壁的沉积是一种十分重要的重金属抗性机制。纤维素、半纤维素和果胶等是细胞壁的主要组成成分,研究发现外源NO能够调节植物细胞细胞壁成分的代谢,如低浓度的NO供体SNP能够促进番茄根部细胞壁纤维素的合成[70],增加土豆嫩叶胼胝质的积累和沉积,而高浓度的NO却起相反的效果[71];另外外源NO能够减少烟草BY- 2悬浮细胞细胞壁果胶、半纤维素和纤维素的含量[72],这些研究都暗示了外源NO可能通过调节植物细胞壁成分含量进而调节植物细胞对重金属胁迫的抗性。实际上,Xiong等证实了在Cd胁迫下,外源NO能够增加水稻根细胞细胞壁果胶和半纤维素的含量,增加Cd在根和茎细胞壁中的积累,减少Cd在叶片可溶性成分中的分布,从而增强水稻对Cd胁迫的抗性[73];Zhang等也发现外源NO能够逆转Al胁迫诱导的水稻根细胞细胞壁果胶和半纤维素含量的增加,并且显著降低Al在水稻根尖和水稻幼苗细胞壁中的积累,从而减少Al胁迫诱导的根的生长抑制和膜脂过氧化,增强水稻对Al胁迫的抗性[74];另外外源NO能够降低小麦和黑麦根尖细胞细胞壁对Al的吸附,增强Al胁迫抗性[75]。因此上述的外源NO缓解重金属胁迫,但并不减少植物对重金属的吸收可能是因为外源NO能够调节植物细胞细胞壁成分,促进了重金属在细胞壁的分布,减少了在细胞可溶性成分中的分布,但总体上并没有减少植物体内的重金属,但也可能是因为存在其他的调节机制。
4 内源NO参与调节植物重金属胁迫抗性
尽管大量的研究表明低浓度的外源NO能够缓解重金属胁迫对植物造成的伤害,增强植物对重金属胁迫的抗性,然而内源NO在调节植物重金属胁迫抗性上的功能角色仍存在争议。有研究发现内源NO是有益的,能够缓解重金属胁迫诱导的毒性,如Tian等发现Al胁迫能够减少芙蓉葵体内的内源NO含量,并且抑制芙蓉葵根的伸长,添加NO的供体SNP能够缓解Al对根的伸长抑制,而NO清除剂c-PTIO、NOS抑制剂L-NAME以及NR抑制剂钨酸盐(Tungstate)却能够加重Al的抑制作用或者抑制外源NO的缓解作用,表明内源NO能够促进根的伸长,增强Al胁迫抗性[76];Qiu等研究发现微波处理能够提高小麦抗氧化酶SOD、POD、CAT、APX、GPX和GR活性以及GSH、ASA和Car含量,缓解Cd胁迫对小麦的氧化伤害,而NO清除剂c-PTIO则能够逆转微波的缓解效果,表明内源NO能够增强小麦体内的抗氧化系统,参与微波增强植物对重金属胁迫的抗性反应[77];Talukdar等发现内源NO与外源NO都能缓解As对菜豆造成的生长抑制和氧化胁迫,增强菜豆As胁迫抗性[78];在荠菜中也有类似的发现,即外源NO与内源NO都能够缓解Cd诱导的膜质过氧化,增强荠菜对Cd胁迫的抗性[79]。
植物螯合素(Phytochelatins, PCs)是一种由半胱氨酸、谷氨酸和甘氨酸组成的含巯基螯合多肽,GSH是植物螯合素的前体,在植物螯合素合成酶催化下合成植物螯合素,植物螯合素能够与进入细胞内的重金属离子螯合,并把重金属隔离在液泡内,从而减轻重金属对细胞质中活性物质的毒害,增强植物对重金属胁迫的抗性[59]。NO能够促进γ-ecs和gshs基因的表达,从而增加苜蓿根内GSH的含量[82],而GSH是植物螯合素的前体,因此可以推断出NO也可能促进植物螯合素的合成,但事实并非如此,在100 μmol/L和150 μmol/L Cd胁迫下,拟南芥悬浮细胞内的植物螯合素含量大幅升高,而添加NOS抑制剂L-NAME后,植物螯合素含量进一步升高,表明内源NO抑制植物螯合素含量的升高,进一步研究发现是因为Cd诱导的内源NO能够通过s-亚硝基化作用与植物螯合素的半胱氨酸残基(Cys)结合形成NO-PC2、NO-PC3和NO-PC4,从而弱化植物螯合素对Cd的解毒作用,促进拟南芥悬浮细胞的细胞凋亡[33],Elviri等也证实了NO与植物螯合素的s-亚硝基化结合[83]。由此可见,内源NO能够使植物螯合素s-亚硝基化,减弱植物螯合素对重金属胁迫的解毒功能,从而促进重金属对植物的毒害。
5 研究展望
尽管国内外已对NO在调节植物重金属胁迫抗性方面做了大量研究,但是NO的具体作用机制仍然不是很清楚。建议未来相关研究应该加强以下几个方面的工作:
(1) 重视NO与其他信号分子如Ca2+、茉莉酸(JA)、水杨酸(SA)和乙烯(ET)等存在的交叉调控。研究表明其他的信号分子也参与调节植物重金属胁迫抗[84],并且NO可能与其他信号分子共同作用来调节植物重金属胁迫抗性,因此加强NO与其他信号分子的交叉调控尤为必要。
(2) 加强对NO靶标分子的研究。NO作为一种信号分子,必然通过刺激靶标分子而发挥作用,而NO正是通过靶标分子进而调节植物对重金属胁迫的抗性,因此加强对NO靶标分子的研究很有意义。
(3) 加强对内源NO作用机制的探讨。重金属胁迫下,内源NO对植物重金属胁迫抗性起着更为重要的作用,并且研究发现内源NO对重金属胁迫具有双重作用,因此内源NO在重金属胁迫抗性上具体的功能角色有待研究。
[1] Zheng C F, Jiang D, Liu F L, Dai T B, Liu W C, Jing Q, Cao W X. Exogenous nitric oxide improves seed germination in wheat against mitochondrial oxidative damage induced by high salinity. Environmental and Experimental Botany, 2009, 67(1): 222- 227.
[2] Bethke P C, Libourel I G L, Jones R L. Nitric oxide reduces seed dormancy inArabidopsis. Journal of Experimental Botany, 2006, 57(3): 517- 526.
[3] Takahashi S, Yamasaki H. Reversible inhibition of photophosphorylation in chloroplasts by nitric oxide. FEBS Letters, 2002, 512(1/3): 145- 148.
[4] He Y K, Tang R H, Yi H, Stevens R D, Cook C W, Ahn S M, Jing L, Yang Z, Chen L, Guo F, Fiorani F, Jackson R B, Crawford N M, Pei Z M. Nitric oxide represses theArabidopsisfloral transition. Science, 2004, 305(5692): 1968- 1971.
[5] Hu X, Neill S J, Tang Z, Cai W. Nitric oxide mediates gravitropic bending in soybean roots. Plant Physiology, 2005, 137(2): 663- 670.
[6] Guo F Q, Crawford N M.Arabidopsisnitric oxide synthase1 is targeted to mitochondria and protects against oxidative damage and dark-induced senescence. The Plant Cell, 2005, 17(12): 3436- 3450.
[7] Neill S J, Desikan R, Hancock J T. Nitric oxide signalling in plants. New Phytologist, 2003, 159(1): 11- 35.
[8] Hong J K, Yun B W, Kang J G, Raja M U, Kwon E, Sorhagen K, Chu C, Wang Y, Loake G J. Nitric oxide function and signalling in plant disease resistance. Journal of Experimental Botany, 2008, 59(2): 147- 154.
[9] Nathan C, Xie Q W. Nitric oxide synthases: roles, tolls, and controls. Cell, 1994, 78(6): 915- 918.
[10] Asai S, Ohta K, Yoshioka H. MAPK signaling regulates nitric oxide and NADPH oxidase-dependent oxidative bursts inNicotianabenthamiana. The Plant Cell, 2008, 20(5): 1390- 1406.
[11] Corpas F J, Barroso J, Carreras A, Valderrama R, Palma J, León A, Sandalio L, Del Río L A. Constitutive arginine-dependent nitric oxide synthase activity in different organs of pea seedlings during plant development. Planta, 2006, 224(2): 246- 254
[12] Del Rio L A, Corpas F J, Sandalio L M, Palma J M, Gomez M, Barroso J B. Reactive oxygen species, antioxidant systems and nitric oxide in peroxisomes. Journal of Experimental Botany, 2002, 53(372): 1255- 1272.
[13] Jasid S, Simontacchi M, Bartoli C G, Puntarulo S. Chloroplasts as a nitric oxide cellular source. Effect of reactive nitrogen species on chloroplastic lipids and proteins. Plant Physiology, 2006, 142(3): 1246- 1255.
[14] Eckardt N A. A functional nitric oxide synthase inOstreococcustauri. The Plant Cell, 2010, 22(11): 3507- 3507.
[15] Guo F Q, Okamoto M, Crawford N M. Identification of a plant nitric oxide synthase gene involved in hormonal signaling. Science, 2003, 302(5642): 100- 103.
[16] Zemojtel T, Frohlich A, Palmieri M C, Kolanczyk M, Mikula I, Wyrwicz L S, Wanker E E, Mundlos S, Vingron M, Martasek P, Durner J. Plant nitric oxide synthase: a never-ending story?. Trends in Plant Science, 2006, 11(11): 524- 525.
[17] Moreau M, Lee G I, Wang Y, Crane B R, Klessig D F. AtNOS/AtNOA1 is a functionalArabidopsisthalianacGTPase and not a nitric-oxide synthase. The Journal of Biological Chemistry, 2008, 283(47): 32957- 32967.
[18] de la Haba P, Agüera E, Benitez L, Maldonado J M. Modulation of nitrate reductase activity in cucumber (Cucumissativus) roots. Plant Science, 2001, 161(2): 231- 237.
[19] Rockel P, Strube F, Rockel A, Wildt J, Kaiser W M. Regulation of nitric oxide (NO) production by plant nitrate reductaseinvivoandinvitro. Journal of Experimental Botany, 2002, 53(366):103- 110.
[20] Stöhr C, Ullrich W R. Generation and possible roles of NO in plant roots and their apoplastic space. Journal of Experimental Botany, 2002, 53(379): 2293- 2303.
[21] Huang J, Sommer E M, Kim-Shapiro D B, King S B. Horseradish peroxidase catalyzed nitric oxide formation from hydroxyurea. Journal of the American Chemical Society, 2002, 124(13): 34733480.
[22] Harrison R. Structure and function of xanthine oxidoreductase: Where are we now? Free Radical Biology and Medicine, 2002, 33(6): 774- 797.
[23] Mansuy D, Boucher J L. Oxidation of N-hydroxy-guanidines by cytochromes P450 and NO-synthases and formation of nitric oxide. Drug Metabolism Reviews, 2002, 34(3): 593- 606.
[24] Cooney R V, Harwood P J, Custer L J, Franke A A. Light-mediated conversion of nitrogen dioxide to nitric oxide by carotenoids. Environmental Health Perspectives, 1994, 102(5): 460- 462.
[25] Wendehenne D, Durner J, Klessig D F. Nitric oxide: a new player in plant signalling and defence responses. Current Opinion in Plant Biology, 2004, 7(4): 449- 455.
[26] Tun N N, Santa-Catarina C, Begum T, Silveira V, Handro W, Floh E I S, Scherer G F E. Polyamines induce rapid biosynthesis of nitric oxide (NO) inArabidopsisthalianaseedlings. Plant and Cell Physiology, 2006, 47(3): 346- 354.
[27] Bartha B, Kolbert Z, Erdei L. Nitric oxide production induced by heavy metals inBrassicajunceaL. Czern. andPisumsativumL.. Acta Biologica Szegediensis, 2005, 49(1/2): 9- 12.
[28] Rodríguez-Serrano M, Romero-Puertas M C, Pazminö D M, Testillano P S, Risueo M C, Del Río L A, Sandalio L M. Cellular response of pea plants to cadmium toxicity: cross talk between reactive oxygen species, nitric oxide, and calcium. Plant Physiology, 2009, 150(1): 229- 243.
[29] Leterrier M, Airaki M, Palma J M, Chaki M, Barroso J B, Corpas F J. Arsenic triggers the nitric oxide (NO) andS-nitrosoglutathione (GSNO) metabolism inArabidopsis. Environmental Pollution, 2012, 166: 136- 143.
[30] Besson-Bard A, Gravot A, Richaud P, Auroy P, Duc C, Gaymard F, Taconnat L, Renou J P, Pugin A, Wendehenne D. Nitric oxide contributes to cadmium toxicity inArabidopsisby promoting cadmium accumulation in roots and by up-regulating genes related to iron uptake. Plant Physiology, 2009, 149(3): 1302- 1315.
[31] Chen F, Wang F, Sun H Y, Cai Y, Mao W H, Zhang G P, Vincze E, Wu F B. Genotype-dependent effect of exogenous nitric oxide on Cd-induced changes in antioxidative metabolism, ultrastructure, and photosynthetic performance in barley seedlings (Hordeumvulgare). Journal of Plant Growth Regulation, 2010, 29(4): 394- 408.
[32] Kopyra M, Stachoń-Wilk M, Gwóz′dz′ E A. Effects of exogenous nitric oxide on the anti-oxidant capacity of cadmium-treated soybean cell suspension. Acta Physiologiae Plantarum, 2006, 28(6):525- 536.
[33] Michele R D, Vurro E, Rigo C, Elviri L, Valentin M D, Careri M, Zottini M, Sanità di Toppi L, Schiavo F L. Nitric oxide is involved in cadmium-induced programmed cell death inArabidopsissuspension cultures. Plant Physiology, 2009, 150(1): 217- 228.
[34] Balestrazzi A, Macovei A, Testoni C, Raimondi E, Donà M, Carbonera D. Nitric oxide biosynthesis in white poplar (PopulusalbaL.) suspension cultures challenged with heavy metals. Plant Stress, 2009, 3(1): 1- 6.
[35] Ma W W, Xu W Z, Xu H, Chen Y S, He Z Y, Ma M. Nitric oxide modulates cadmium influx during cadmium-induced programmed cell death in tobacco BY- 2 cells. Planta, 2010, 232(2): 325- 335.
[36] Mittler R, Vanderauwera S, Gollery M, Breusegem F V. Reactive oxygen gene network of plants. Trends in Plant Science, 2004, 9(10): 490- 498.
[37] Ammar W B, Nauairi I, Zarrouk M, Jamel F. Cadmium stress induces changes in the lipid composition and biosynthesis in tomato (LycopersiconesculentumMill.) leaves. Plant Growth Regulation, 2007, 53(2): 75- 85.
[38] Romero-Puertas M C, Palma J M, Gomez M, Del Rio L A, Sandalio L M. Cadmium causes the oxidative modification of proteins in pea plants. Plant Cell and Environment, 2002, 25(5): 677- 686.
[39] Britt A B. Molecular genetics of DNA repair in higher plants. Trends in Plant Science, 1999, 4(1): 20- 25.
[40] Gill S S, Tuteja N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiology and Biochemistry, 2010, 48(12): 909- 930.
[41] Papadakis A K, Roubelakis-Angelakis K A. Polyamines inhibit NADPH oxidase-mediated superoxide generation and putrescine prevents programmed cell death induced by polyamine oxidase-generated hydrogen peroxide. Planta, 2005, 220(6): 826- 837.
[42] Wang L N, Yang L M, Yang G J, Li X G, Song Y P, Wang X F, Hu X Y. Involvements of H2O2and metallothionein in NO-mediated tomato tolerance to copper toxicity. Journal of Plant Physiology, 2010, 167(15): 1298- 1306.
[43] Panda P, Nath S, Chanu T T, Sharma G D, Panda S K. Cadmium stress-induced oxidative stress and role of nitric oxide in rice (OryzasativaL.). Acta Physiologiae Plantarum, 2011, 33(15): 1737- 1747.
[44] Laspina N V, Groppa M D, Tomaro M L, Benavides M P. Nitric oxide protects sunflower leaves against Cd-induced oxidative stress. Plant Science, 2005, 169(2): 323- 330.
[45] Kazemi N, Khavari-Nejad R A, Fahimi H, Saadatmand S, Nejad-Sattari T. Effects of exogenous salicylic acid and nitric oxide on lipid peroxidation and antioxidant enzyme activities in leaves ofBrassicanapusL. under nickel stress. Scientia Horticulturae, 2010, 126(3): 402- 407.
[46] Tewari R K, Hahn E J, Paek K Y. Modulation of copper toxicity-induced oxidative damage by nitric oxide supply in the adventitious roots ofPanaxginseng. Plant Cell Reports, 2008, 27(1): 171- 181.
[47] Xu J, Wang W Y, Yin H X, Liu X J, Sun H, Min Q. Exogenous nitric oxide improves antioxidative capacity and reduces auxin degradation in roots ofMedicagotruncatulaseedlings under cadmium stress. Plant and Soil, 2010, 326(1/2): 321- 330.
[48] Filippou P, Antoniou C, Fotopoulos V. The nitric oxide donor sodium nitroprusside regulates polyamine and proline metabolism in leaves ofMedicagotruncatulaplants. Free Radical Biology and Medicine, 2013, 56: 172- 183.
[49] Groppa M D, Rosales E P, Iannone M F, Benavides M P. Nitric oxide, polyamines and Cd-induced phytotoxicity in wheat roots. Phytochemistry, 2008, 69(14): 2609- 2615.
[50] Zhang L P, Mehta S K, Liu Z P, Yang Z M. Copper-Induced proline synthesis is associated with nitric oxide generation inChlamydomonasreinhardtii. Plant and Cell Physiology, 2008, 49(3): 411- 419.
[51] Singh H P, Batish D R, Kaur G, Arora K, Kohli R K. Nitric oxide (as sodium nitroprusside) supplementation ameliorates Cd toxicity in hydroponically grown wheat roots. Environmental and Experimental Botany, 2008, 63(1/3): 158- 167.
[52] Singh H P, Kaur S, Batish D R, Sharma V P, Sharma N, Kohli P K. Nitric oxide alleviates arsenic toxicity by reducing oxidative damage in the roots ofOryzasativa(rice). Nitric Oxide, 2009, 20(4): 289- 297.
[53] Srivastava S, Dubey R S. Nitric oxide alleviates manganese toxicity by preventing oxidative stress in excised rice leaves. Acta Physiologiae Plantarum, 2012, 34(2): 819- 825.
[54] Hsu Y T, Kao C H. Cadmium toxicity is reduced by nitric oxide in rice leaves. Plant Growth Regulation, 2004, 42(3): 227- 238.
[55] Caro A, Puntarulo S. Nitric oxide decreases superoxide anion generation by microsomes from soybean embryonic axes. Physiologia Plantarum, 1998, 104(3): 357- 364.
[56] Delledonne M, Zeier J, Marocco A, Lamb C. Signal interactions between nitric oxide and reactive oxygen intermediates in the plant hypersensitive disease resistance response. Proceedings of the National Academy of Sciences of the United States of America, 2001, 98(23): 13454- 13459.
[57] Martinez G R, Mascio P D, Bonini M G, Augusto O, Briviba K, Sies H, Maurer P, Röthlisberger U, Herold S, Koppeno W H. Peroxynitrite does not decompose to singlet oxygen (1ΔgO2) and nitroxyl (NO-). Proceedings of the National Academy of Sciences of the United States of America, 2000, 97(19): 10307- 10312.
[58] Thompson J E, Legge R L, Barber R F. The role of free radicals in senescence and wounding. New Phytologist, 1987, 105(3): 1044- 1047.
[59] Zhang H Y, Xu W Z, Guo G B, He Z Y, Ma M. Coordinated responses of phytochelatins and metallothioneins to heavy metals in garlic seedlings. Plant Science, 2005, 169(6): 1059- 1065.
[60] Qu W, Liu J, Fuquay R, Shimoda R, Sakurai T, Saavedra J E, Keefer L K, Waalkes M P. The nitric oxide prodrug, V-PYRRO/NO, protects against cadmium toxicity and apoptosis at the cellular level. Nitric Oxide, 2005, 12(2): 114- 120.
[61] Misra R R, Hochadel J F, Smith G T, Cook J C, Waaalkes M P, Wink D A. Evidence that nitric oxide enhances cadmium toxicity by displacing the metal from metallothionein. Chemical Research in Toxicology, 1996, 9(1): 326- 332.
[62] Katakai K, Liu J, Nakajima K, Keefer L K, Waalkes M P. Nitric oxide induces metallothionein (MT) gene expression apparently by displacing zinc bound to MT. Toxicology Letters, 2001, 119(2): 103- 108.
[63] Abdel-Kader D E Z. Role of nitric oxide, glutathione and sulfhydryl groups in zinc homeostasis in plants. American Journal of Plant Physiology, 2007, 2(2): 59- 75.
[64] Wang Y S, Yang Z M. Nitric oxide reduces aluminum toxicity by preventing oxidative stress in the roots ofCassiatoraL.. Plant and Cell Physiology, 2005, 46(12): 1915- 1923.
[65] Hu K D, Hu L Y, Li Y H, Zhang F Q, Zhang H. Protective roles of nitric oxide on germination and antioxidant metabolism in wheat seeds under copper stress. Plant Growth Regulation, 2007, 53(3): 173- 183.
[66] Phang I C, Leung D W M, Taylor H H, Burritt D J. The protective effect of sodium nitroprusside (SNP) treatment onArabidopsisthalianaseedlings exposed to toxic level of Pb is not linked to avoidance of Pb uptake. Ecotoxicology and Environmental Safety, 2011, 74(5): 1310- 1315.
[67] Xu J, Wang W J, Sun J H, Zhang Y, Ge Q, Du L G, Yin H X, Liu X J. Involvement of auxin and nitric oxide in plant Cd-stress responses. Plant and Soil, 2011, 346(1/2): 107- 119.
[68] Lux A, Martinka M, Vaculk M, White P J. Root responses to cadmium in the rhizosphere: a review. Journal of Experimental Botany, 2011, 62(1): 21- 37.
[69] Yang J L, Li Y Y, Zhang Y J, Zhang S S, Wu Y R, Wu P, Zheng S J. Cell wall polysaccharides are specifically involved in the exclusion of aluminum from the rice root apex. Plant Physiology, 2004, 146(2): 602- 611.
[70] Correa-Aragunde N, Lombardo C, Lamattina L. Nitric oxide: an active nitrogen molecule that modulates cellulose synthesis in tomato roots. New Phytologist, 2008, 179(2): 386- 396.
[71] París R, Lamattina L, Casalongue C A. Nitric oxide promotes the wound-healing response of potato leaflets. Plant Physiology and Biochemistry, 2007, 45(1): 80- 86.
[72] Pacoda D, Montefusco A, Piro G, Dalessandro G. Reactive oxygen species and nitric oxide affect cell wall metabolism in tobacco BY- 2 cells. Journal of Plant Physiology, 2004, 161(10): 1143- 1156.
[73] Xiong J, An L Y, Lu H, Zhu C. Exogenous nitric oxide enhances cadmium tolerance of rice by increasing pectin and hemicellulose contents in root cell wall. Planta, 2009, 230(4): 755- 765.
[74] Zhang Z Y, Wang H H, Wang X M, Bi Y R. Nitric oxide enhances aluminum tolerance by affecting cell wall polysaccharides in rice roots. Plant Cell Reports, 2011, 30(9): 1701- 1711.
[75] 何虎翼, 何龙飞, 黎晓峰, 顾明华. 铝胁迫下硝普钠对黑麦和小麦根尖细胞壁铝吸附的影响. 广西农业生物科学, 2007, 26(3): 235- 239, 249- 249.
[76] Tian Q Y, Sun D H, Zhao M G, Zhang W H. Inhibition of nitric oxide synthase (NOS) underlies aluminum-induced inhibition of root elongation inHibiscusmoscheutos. New Phytologist, 2007, 174(2): 322- 331.
[77] Qiu Z B, Guo J L, Zhang M M, Lei M Y, Li Z L. Nitric oxide acts as a signal molecule in microwave pretreatment induced cadmium tolerance in wheat seedlings. Acta Physiologiae Plantarum, 2013, 35(1): 65- 73.
[78] Talukdar D. Arsenic-induced oxidative stress in the common bean legume,PhaseolusvulgarisL. seedlings and its amelioration by exogenous nitric oxide. Physiology and Molecular Biology of Plants, 2013, 19(1): 69- 79.
[79] Verma K, Mehta S K, Shekhawat G S. Nitric oxide (NO) counteracts cadmium induced cytotoxic processes mediated by reactive oxygen species (ROS) in Brassica juncea: cross-talk between ROS, NO and antioxidant responses. Biometals, 2013, 26(2):255- 269.
[80] Ye Y, Lin Z, Xing D. Nitric oxide promotes MPK6-mediated caspase- 3-like activation in cadmium-inducedArabidopsisthalianaprogrammed cell death. Plant, Cell and Environment, 2013, 36(1): 1- 15.
[81] Arasimowicz-Jelonek M, Floryszak-Wieczorek J, Deckert J, Rucińska-Sobkowiak R, Gzyl J, Pawlak-Spradaa S, Abramowskib D, Jelonekc T, Gwóz′dz′a E A. Nitric oxide implication in cadmium-induced programmed cell death in roots and signaling response of yellow lupine plants. Plant Physiology and Biochemistry, 2012, 58: 124- 134.
[82] Innocenti G, Pucciariello C, Gleuher M L, Hopkins J, De Stefano M, Delledonne M, Puppo A, Bandouin E, Frendo P. Glutathione synthesis is regulated by nitric oxide inMedicagotruncatularoots. Planta, 2007, 225(6): 1597- 1602.
[83] Elviri L, Speroni F, Careri M, Mangia A, Sanità di Toppi L, Zottini M. Identification ofinvivonitrosylated phytochelatins inArabidopsisthalianacells by liquid chromatography-direct electrospray-linear ion trap-mass spectrometry. Journal of Chromatography A, 2010, 1217(25): 4120- 4126.
[84] Maksymiec W. Signaling responses in plants to heavy metal stress. Acta Physiologiae Plantarum, 2007, 29(3):177- 187.
Advances on effects of nitric oxide on resistances of plants to heavy metal stress
XIA Haiwei, SHI Guoxin*, HUANG Min, WU Juan
JiangsuKeyLaboratoryofBiodiversityandBiotechnology,CollegeofLifeScience,NanjingNormalUniversity,Nanjing210023,China
Heavy metal pollution has become an increasingly serious environmental problem because heavy metal can be easily taken up by plants, leading to inhibition of plant growth and development. Hence, it is necessary to investigate resistances of plants to heavy metal stress. As an important signaling molecule, nitric oxide (NO) is involved in the regulation of multiple plant responses to a variety of abiotic and biotic stresses. Recently, an increasing number of articles have reported the effects of NO on resistances of plants to heavy metal stress. However, studies which systematically summarize the molecular mechanisms of NO on resistances of plants to heavy metal stress are quite limited. This research mainly reviews the pathways of NO production, changes of endogenous NO contents under heavy metal stress and influences of exogenous and endogenous NO on resistances to heavy metal stress. The sources of NO production in plants involve not only enzymatic reaction pathways which include nitric oxide synthase (NOS), nitrate reductase (NR), nitrite reductase (Ni-NOR) pathways and etc. but also non-enzymatic reaction pathways. Many authors have noted discrepant reports on the effects of heavy metal stress on endogenous NO content in plants and the observed differences in endogenous NO accumulation are frequently ascribed to the use of different duration of treatment, heavy metal concentrations, species and genotypes of plants and varieties of plant tissues. Interestingly, all plant cell suspensions show a visible increase in endogenous NO accumulation under heavy metal stress, which is attributed to the fact that they are lacking of network regulation between cells and cells. Meanwhile, it has been demonstrated that exogenous NO could enhance antioxidant defence system of plant cells, act as an antioxidant promoting direct scavenging of reactive oxygen species, induce metallothioneins (MTs) gene expression by displacing heavy metal bound to MTs, and affect the uptake of heavy metal into plants and the distribution of heavy metal in plant cells though regulating the metabolism of cell wall composition, consequently relieve heavy metal toxicity and enhance resistances of plants to heavy metal stress. However, the functional roles of endogenous NO in regulating resistances of plants to heavy metal stress are controversial. Some research show that endogenous NO is helpful for alleviating heavy metal-induced toxicity. On the contrary, some evidences indicate that endogenous NO is harmful, and participates in heavy metal-induced cell toxicity and programmed cell death through accelerating the absorption of heavy metal andS-nitrosylation of phytochelatins. These conflicting data suggest that NO may have a dual effect on resistances to heavy metal stress, but great efforts are required in order to clarify this speculation. Finally, interrelated perspectives are also discussed to further study the relationship between NO and resistances to heavy metal stress.
exogenous NO; endogenous NO; heavy metal stress; resistances
江苏高校优秀学科建设工程资助项目(164320H106)
2013- 06- 24;
2014- 05- 30
10.5846/stxb201306241770
*通讯作者Corresponding author.E-mail: gxshi@njnu.edu.cn
夏海威,施国新,黄敏,吴娟.一氧化氮对植物重金属胁迫抗性的影响研究进展.生态学报,2015,35(10):3139- 3147.
Xia H W, Shi G X, Huang M, Wu J.Advances on effects of nitric oxide on resistances of plants to heavy metal stress.Acta Ecologica Sinica,2015,35(10):3139- 3147.
猜你喜欢
食品工业科技(2023年4期)2023-02-14
山东科学(2020年3期)2020-06-11
中国食品学报(2019年10期)2019-11-12
浙江工业大学学报(2017年5期)2018-01-22
广东农业科学(2017年5期)2017-08-29
上海农业学报(2016年2期)2016-10-27
华东理工大学学报(自然科学版)(2015年4期)2015-12-01
动物营养学报(2015年10期)2015-12-01
中国洗涤用品工业(2015年2期)2015-02-28
云南中医学院学报(2014年3期)2014-07-31
