
北京东灵山森林植物多样性的网络结构特征
2015-02-07 06:51陈禹舟马克明张育新牛树奎
生态学报 2015年11期
陈禹舟, 马克明, 张育新, 张 霜, 牛树奎
1 中国科学院生态环境研究中心城市与区域生态国家重点实验室, 北京 100085 2 北京林业大学森林培育与保护省部共建教育部重点实验室, 北京 100083
北京东灵山森林植物多样性的网络结构特征
陈禹舟1,2, 马克明1,*, 张育新1, 张 霜1, 牛树奎2
1 中国科学院生态环境研究中心城市与区域生态国家重点实验室, 北京 100085 2 北京林业大学森林培育与保护省部共建教育部重点实验室, 北京 100083
一个群落可以看作是由物种相互连接的复杂系统,刻画其网络结构有助于深入揭示系统性质以及结构与功能之间的关系。在生态学中,复杂网络理论已被成功应用于食物网与互利网络的结构研究,但尚未检验其刻画生物多样性格局的能力。采用复杂网络理论研究了北京东灵山森林乔木层、灌木层、草本层植物关联关系的网络结构特征及其差异。结果表明,植物物种的共同出现是非随机的,并表现出一定的小世界模式;乔木层、灌木层、草本层植物共同出现的网络在结构特征上存在明显差异,草本层网络比乔木层和灌木层网络更松散且平均路径更长,灌木层和草本层网络的聚类系数高于乔木层且存在度的幂律分布。皆表明复杂网络理论具备反映不同层植物多样性格局差异的能力。
群落联结; 分布格局; 复杂网络; 共有物种
种间联结(物种联结性)是指不同物种在空间分布上的相互关联性,通常是由于群落生境的差异影响了物种的分布而引起的[1]。这样种间联结性就是对各个物种在不同生境中相互影响相互作用所形成的有机联系的反映。研究群落种间联结和分布格局不但有助于更深刻地认识群落的结构、功能、演替和分类,而且对于维持群落稳定性、生物多样性和制订珍稀濒危物种保护措施均有重要理论价值[2]。
目前,针对种间联结性的研究集中在种间关系上,主要是通过统计学的手段考察某两个物种共同出现的或正或负的相关性[3- 8]。然而具有相互作用的个体联系起来也可以看作是一个系统。复杂系统理论认为,大量个体(更典型的是具有适应性的主体)所组成的复杂系统,在没有中心控制、非完全信息、仅仅存在局域相互作用的条件下,通过个体之间的非线性相互作用,可以在宏观层次上表现出一定的结构和功能,产生涌现特性。就像生命从无机物中涌现,意识从动物群体中涌现,社会组织从个体行为中涌现[9]。把群落看作是一个复杂系统有助于发掘一些传统方法尚未注意到的特性,并能从更深的层面探讨物种之间的相互作用。
如果把所有具有关联关系的群落看作是一个复杂系统,就不能仅仅依靠常规统计分析手段来进行研究。为了理解系统的行为,需要理解系统的拓扑结构。而复杂网络理论近年来的发展,使得其可以被用于理解系统的拓扑结构。复杂网络理论在21世纪以来取得了十分重大的进展。其中最为重要的发现就是,在现实世界中,许多网络虽然节点代表的对象不同,却体现出了相似的统计特性[10]。这个发现之所以重要有两个原因:首先,这说明所有的复杂系统可能存在一个共有的结构模式。其次,结构决定功能,弄清楚系统的结构模式有助于人们更进一步了解系统的运行机制。复杂网络理论已经作为一种新方法在各个领域发挥着作用。例如,分子生物学家曾受制于一次只能研究少数几个基因的作用,而现在他们可以做出描述基因相互依赖关系的图谱来实现例如基因活化这一类的基因控制[11]。生态学家通过将复杂网络应用于食物网研究,发现了“二度分割”现象[12],即在食物网中节点的平均路径长度是2。也就是说,在食物网中,一个物种平均只需要通过两个物种就能对另一个物种产生影响。还有人在食物网中发现了“小世界”模式[13],即在大的食物网中存在着若干个相对致密的子网络。这是由于节点自身特性导致其倾向于跟同一类的节点发生连接,形成所谓的“小世界”。这些发现都说明食物网中的各个物种都处于紧密连接的状态[14]。而在对互利网络的研究中也发现了其具有高度的“嵌套式”结构[15]。种间关联的研究也有着类似的问题,即只局限于个别几个物种之间的关系研究。可见,通过引入复杂网络理论,有助于从整体上了解种间关联遵循什么样的模式。
这种模式即体现为物种分布格局。研究认为,物种分布格局的形成受物种生物学特性和环境条件的影响[16- 17]。它们将导致物种可能呈现集群或者随机分布[18]。Diamond曾通过零模型预言了共有物种的分布格局呈现非随机分布[19],随后Gotelli对96个种间关联矩阵进行Meta-Analysis验证了Diamond的预言[20]。Smith认为只有在环境条件均匀的少数地区才可能出现随机分布[21]。目前,对于物种分布格局的探讨往往局限于群落优势种或者珍稀种的分布[22- 26],其结论也就局限于所研究的物种。而将研究对象根据生活型分为草本、灌木、乔木三层,分别刻画其网络结构特征,将有助于了解网络分析方法刻画植物多样性格局的能力以及不同层植物多样性格局的差异性。
1 研究区域与调查方法
1.1 研究区域
研究地点位于北京市门头沟区东灵山小龙门林场的中国科学院北京森林生态系统定位研究站(图1)。年降雨量为 500—650 mm。研究区域为该地的一个小流域,位于39°56′58″—39°57′28″ N, 115°25′18″—115°25′55″ E,海拔高度范围在1252—1530 m,面积约为1km2。暖温带落叶阔叶林是典型地带性植被,主要类型包括辽东栎林(Quercuswutaishanica)、山杨林(Populusdavidiana)、桦木林(Betulaspp.),其主要伴生树种包括槭树(Acerspp.)、椴树(Tiliaspp.)、大叶白腊(Fraxinusrhynchophylla)以及核桃楸(Juglansmandshurica)等。
1.2 调查方法
区分不同斑块,对该小流域中的全部森林类型进行群落调查。采取20 m × 20 m样方调查方式,根据群落组成,随机设置共76个标准样方,包括不同坡位及坡向。逐一记录各乔木树种的名称、树高、冠幅、胸径等;在每个乔木样方内设置3个5 m × 5 m灌木样方沿主对角线方向等间距分布,记录各灌木物种的名称、高度、冠幅、盖度等;在每个灌木样方内3个1 m × 1 m 草本样方,同样沿主对角线方向等间距设置,分别记录物种名、株高、盖度。研究区域的植被及样地分布如图1所示。

图1 研究区位置、植被及样地分布,图中原点代表样地
2 数据处理方法
为了研究该区域物种共同出现的结构模式,将76个标准样地视为76个节点。再逐一对比各个样地中物种的关联情况,分别考察乔木层、灌木层、及草本层。如果两个样地间具有共有物种,则视为具有连接,并以共有物种的数目来定义连接强度。以此分别构建其乔木层、灌木层、以及草本层3个网络。为了使各个层的结构能够更为清晰显现,在不影响研究结果的前提下,按照乔灌草最大连线值之比,删除了连线值低于6、8、10的连线所对应的节点。然后再分别考察3个网络的属性和结构特征,包括:(1)平均路径长度、(2)平均聚类系数、和(3)度分布。
曾有研究通过统计学方法证明了生态网络的不均匀性对于维持系统稳定的重要性[27]。分析这些网络属性,可以帮助我们检测复杂网络理论是否可用于种间关系研究。通过与随机网络的结构属性进行对比,了解种间关联的网络是否也具有不均匀性。
2.1 平均路径长度[28]
在复杂网络理论中,对于路径长度的定义是从一个节点到另一个节点所经历的最少的边的数目,通常表示为dij。网络的平均路径长度的定义为任意两个点之间路径长度的平均值,即:
(1)
式中,N为网络的节点数目。就共有物种网络而言,网络的平均路径长度衡量了样地间的相似性。
2.2 聚类系数
聚类系数[28]是用来衡量网络中相对密集情况的系数。在共有物种网络中,相对密集的子网络中的点代表了一类具有共性的点相互连接所体现出的特征。通过分析聚类系数可进一步从整体上了解节点的关联性,即样地中共有物种的关联程度。假设一个网络中的节点i有ki个节点直接与其相连。这ki个节点就称为i的邻点。而在这ki个节点之间最多可能有k(k-1)/2条边。而这ki个节点之间实际存在的边数Ei比上总的可能的边数k(k-1)/2就是节点i的聚类系数Ci,即:
Ci=2Ei/ki(ki-1)
(2)
从几何上看,上式也可等价理解为:
(3)
而与点i相连的三元组表示的是点i以及与其相连的两个节点。它们应该至少含有两条边。
整个网络的聚类系数C就是单个节点的聚类系数Ci相加之后取的平均值:
(4)
2.3 度与度分布[29]
节点i的度ki的定义为该节点连接的其它节点的数目。在共有物种网络中,一个节点的度越大则说明该节点所代表的样地越能够体现样地之间的共有物种。网络中所有节点i的度ki的平均值称为网络的(节点)平均度,记为
对于随机网络而言,度分布函数P(k)服从泊松分布,记为:
(5)
其形状在远离均值
而近年来的研究表明,许多现实网络包括互联网、社会网、食物网、新陈代谢等网络的度分布都不同于泊松分布而呈现幂律分布,记为:
P(k)=cx-α(c,α为正数)
(6)
通过对上式两端进行对数变换可以得到
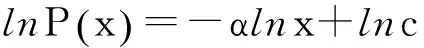
(7)
可以看出,在双对数坐标下幂律分布则表现为一条直线。通过将实验数据在双对数坐标下做线性回归分析,可以判断其是否服从幂律分布[30]。
服从幂律分布是“小世界”模式网络的主要特征,而明确有别于随机网络。除此之外,具有“小世界”模式的网络具有较高的聚类现象,而任意两个节点的距离一般相对较短。而对于随机网络(由节点按照一定概率随机连接而成)而言,节点距离同样较短,但不存在聚类现象。因此,通过跟随机网络的特征进行比较,可以判断共有物种形成的网络是遵循某种模式,还是随机连接构成。
分别计算乔、灌、草3层不同的植物所构成的共有物种网络的属性之后,再用Pajek软件针对每一层生成节点数相同的随机网络各1000个,得到它们的网络属性之后取平均值,再与共有物种网络进行比较。
3 结果
将图2和图3进行比较可以看出,共有物种网络的平均路径长度和随机网络接近。但是草本层和灌木层的聚类系数远高于随机网络,其聚类系数之比分别为3.37和2.90;而乔木层的聚类系数跟随机网络相比则较为接近一些,聚类系数比为1.67。
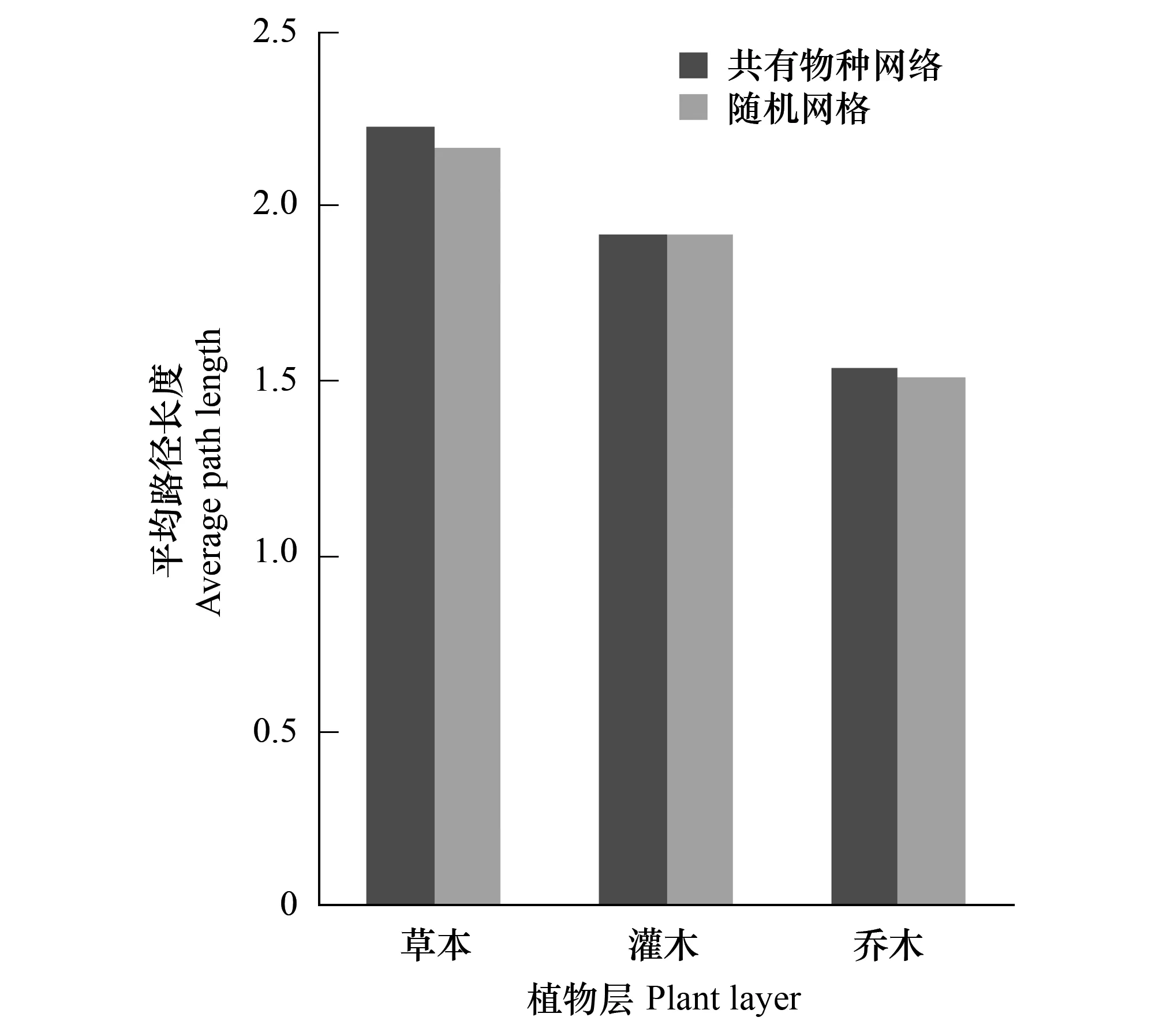
图2 共有物种网络和相应随机网络的平均路径
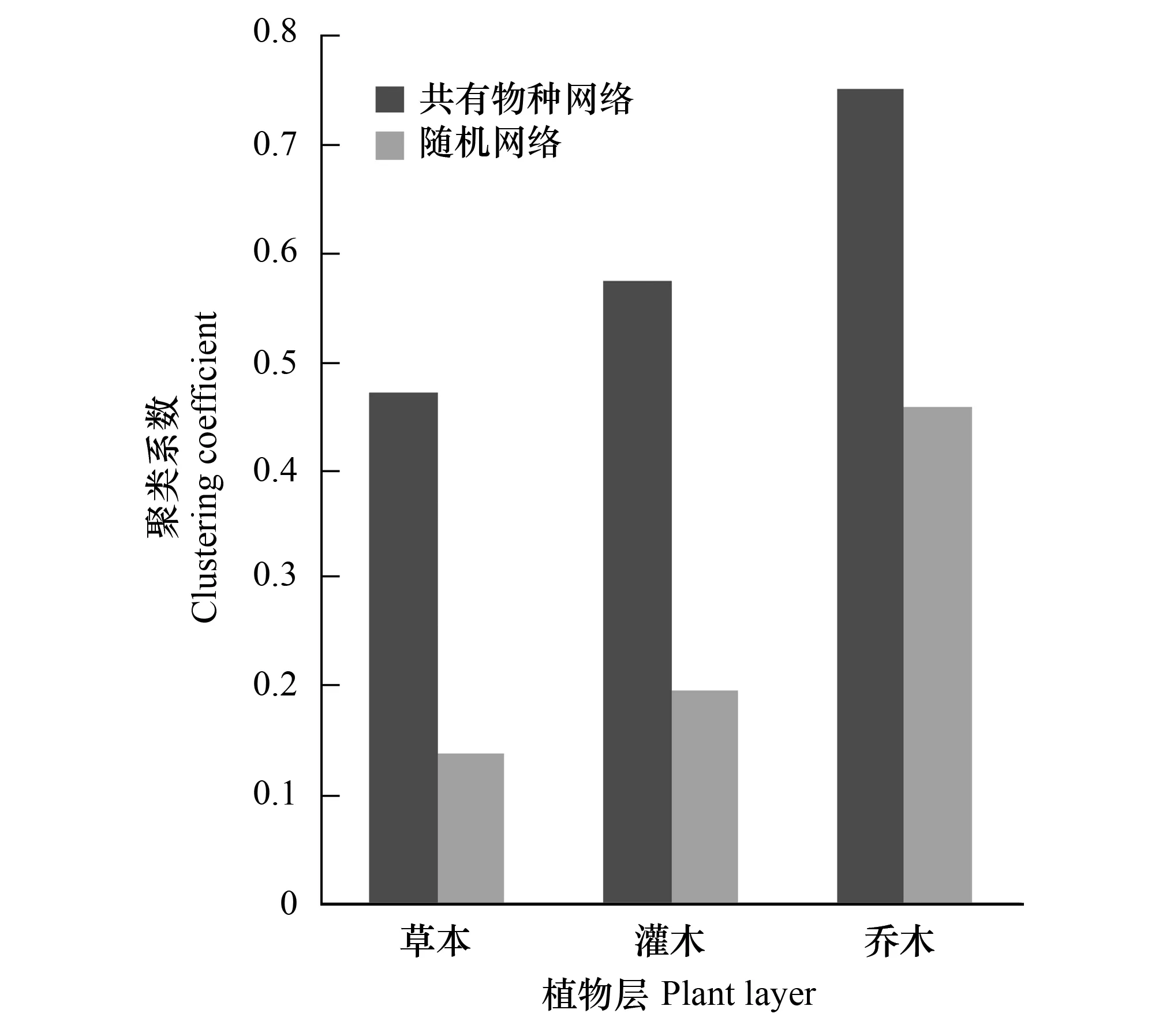
图3 共有物种网络和相应随机网络的聚类系数
对k和P(k)取对数以后进行的回归分析表明,草本层和灌木层服从幂律分布。而乔木层不服从(图4和表1)。
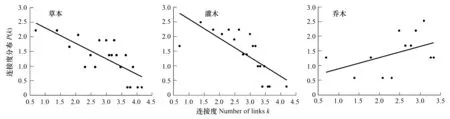
图4 不同层植物连接度分布

表1 不层植物幂律分布拟合方程
横向比较乔、灌、草三层的网络属性则可以发现,这3个网络的结构特征跟其节点的属性有关。从乔木到灌木再到草本,这三类不同层的植物,个体寿命和大小依此递减,平均路径长度和聚类系数却分别呈现递增和递减的分布。
4 讨论
复杂网络理论用于物种多样性分布格局(种间关联)研究,可以获得以往研究所不能发现的新结果,具有一定理论价值。通过比较同一区域中乔灌草三层的网络特性,可以从拓扑学的角度揭示不同生活型的植物在相同的环境条件下所表现出的不同的物种多样性格局特征。
首先,从不同层植物多样性的关联网络的平均路径递增和聚类系数递减的结果可以看出,在共有物种网络中存在“小世界”模式。
通过将共有物种网络跟随机网络进行比较可以发现,草本植物和灌木植物组成的共有物种网络表现出一定的“小世界”特性。其平均路径距离跟随机网络接近而聚类系数远大于随机网络。这说明网络中存在相对密集的子网络,节点之间体现出了关联的偏好。而对于乔木而言,其平均路径最低且聚类系数最大。但是整个网络中的点都是紧密连接在一起的,节点的关联偏好并不如草灌明显。
共有物种网络所产生的聚类现象是因为样地所具有的相似特性,以共有物种的数量表现出来,共有物种越多说明越相似。现实网络中的聚类现象在人类社会中就体现在“物以类聚,人以群分”,在一个大网络下形成若干相对密集的子网络。在一个具有“小世界”模式的大肠杆菌的新陈代谢网络中,聚类系数可以达到随机网络系数的17倍以上[11]。但是本研究中的共有物种网络只有3倍多,差异程度不如其它非生态网络明显。
其次,共有物种网络的连接度服从幂律分布说明植物多样性分布的不均匀特性。
服从幂律分布从直观上来看就是大部分节点跟其它节点连接较少,而少数节点却跟大多数节点都有连接。幂律分布的发现者Barabasi和Albert把这种服从幂律分布的网络又叫做“无标度(scale-free)网络”[29],因为在这个分布中的点在横跨几个标度的数量级下依然保持增长特性,而明显有别于随机网络服从的泊松分布。在随机网络的泊松分布中,节点的度是在平均值
这种不均匀特性产生的一个问题就是,在物种节点是遵循了怎样的连接原则使得其能够在不同标度下都产生了相同的特性?这种不均匀的网络特征是如何形成的?Watts和Strogatz在模拟“小世界”网络的动态过程中提出的“优先连接”模型,可能是一个潜在的解决方案。这个算法设计的原则就是节点会优先连接那些点度高的节点[28]。这种模型已经被证明可以用来解释技术网络以及新陈代谢网络的形成机制[29- 30]。
但是这个原则跟生态系统的规律有不容的地方。按照生态位理论[31],一个区域的生态位是有限的,能够容纳的物种数量也是有限的。当区域中一部分物种率先占领了各自的生态位以后,新加入的物种就会在这些生态位上形成竞争。而竞争将限制新的物种的涌入,于是物种可能会选择出现在其它区域。在这个规律的限制线下,物种的数目不可能突破几个数量级增长。因此,从本研究数据的表现上来看就是幂指数的值小于非生态网络的值。在共有物种网络中幂律指数的值是0.53和0.65,而通常的非生态网络幂指数的值在2到3之间[32]。因此未来研究应该结合种间关联的数据,考虑种间竞争或互利作用来进一步探讨幂律分布是如何被影响的。
同时乔木层的网络度分布既不服从幂律分布也不服从泊松分布,点度高的点出现的概率也高,且乔木网络具有最高的聚类系数,说明乔木虽然种类有限,但是在各个样地中都能占到一席之地。草本灌木相对乔木而言,对于环境变化的指示作用更为明显。
最后,在共有物种网络中,不同层植物由于其自身特性而产生了结构上的差异。这一发现对于深入理解生物多样性格局与群落演替过程、物种生态位、种子传播等之间的关系提供了新角度。
在同一区域中,寿命较长和个体较大的乔木倾向于出现有限的几个种,而灌木和草本相对来说则更为多样化一些。乔木和灌木相比草本在适应气候变化的速度上更慢,草本的演替速度是木本的2到10倍[33]。因为年龄越大的木本植物生活繁殖周期就越长,许多树木需要较长的营养生长时间之后,才能进行生殖生长,于是其累积基因变化的速率也就越慢。而这样就导致木本植物相比草本植物的突变率较低,产生的种类就比草本植物要少。又由于草本的数量多且更新速度快,在漫长的演替过程中,当环境发生变化时,往往也是草本植物更能适应环境,于是有更多的种类保存下来。即使是在相对稳定的环境,由于乔灌木体积和生物量较大,对生境质量的要求相对较高。
除此之外,乔木的生存还涉及到对共生生物的要求,比如真菌以及传播种子的动物。相比而言,能够容纳草本植物的生态位则更为多样。比如树木的阴影,岩石缝隙或者土壤养分较为贫乏的区域,虽然无法满足乔灌木的生长条件,却可能让某一种草本植物留存下来。又由于草本植物大多通过风媒传播,扩散速度更快,传播范围也大于果实体积较大的乔木(例如辽东栎)。这就导致一个区域内的草本植物能够散播在更多的地方,占领更多乔木无法占领的生态位,同时其它区域的草本植物也更容易突破地理条件的限制来到这片区域中。
总之,共有物种网络中表现出的小世界模式,说明物种多样性分布格局是非随机的。相比以往研究,本研究将物种关联关系看做是一个复杂网络,是对关联关系从全局上更为清晰的界定,并且在此基础上揭示了小世界模式和幂律分布规律。把复杂系统看做是一个网络进行研究,不仅可以发掘其中的模式,还能在此基础上对其动态行为进行预测或者研究其崩溃的信号。这也是复杂网络理论相比传统研究方法的优势所在。
5 问题与展望
本研究成功刻画了小流域森林群落共有物种的网络结构,发现其做为复杂系统所存在的特性:小世界模式和度的幂律分布,也发现了不同层的植物网络结构所存在的差异,说明复杂网络方法具有描述物种多样性分布格局的能力。但本研究仍存在不足之处。因计算方法限制,本研究对于网络属性的探讨是删除了部分强度较低的连线之后进行的,而这些连线对网络属性的影响尚缺乏定量研究。同时,样本数量,在其它度的幂律分布的网络中,比如人类社会或者互联网的节点数量往往是达到数万甚至上亿,在探索“小世界”模式的食物网的节点数量也都有100个左右,而本研究中网络节点只有76个,这在揭示幂律分布的“无标度”特性上是否有所限制,也有待进一步研究。
目前,复杂网络还是一门新兴的交叉学科,小世界模式和度的幂律分布这两个特征同时也在出现在其它领域的复杂网络研究中。这种不同领域间的研究所发现的共性,说明各学科正在从不同角度接近更为本质的问题。不同学科从各自的角度进入网络科学可以互为补充,又可以深化各自的研究问题。未来研究应该在此基础上,通过建模探讨网络演化,进一步阐明系统性质以及结构与功能之间的关系。这需要将物种间的竞争或互利作用以及环境因子的影响引入,最终实现对系统行为进行预测和调控的目的。同时,未来还应将复杂网络方法更多地应用于生态学研究中,这一探索过程也是逐步完善复杂网络理论的过程。
[1] 彭少麟, 周厚诚, 郭少聪, 黄忠良. 鼎湖山地带性植被种间联结变化研究. 植物学报, 1999, 41(11): 1239- 1244.
[2] 林勇明, 洪滔, 吴承祯, 洪伟, 胡喜生, 范海兰, 宋萍. 桂花次生林群落主要树种种间关联及其对混交度的响应. 应用与环境生物学报, 2007, 13(3): 327- 332.
[3] 郭志华, 卓正大, 陈洁, 吴梅凤. 庐山常绿阔叶、落叶阔叶混交林乔木种群种间联结性研究. 植物生态学报, 1997, 21(5): 424- 432.
[4] 娄彦景, 赵魁义. 三江平原毛苔草群落近30年演替过程中的种间联结性分析. 生态学杂志, 2008, 27(4): 509- 513.
[5] 李建民, 谢芳, 陈存及, 张思玉, 肖日红, 赵大洲. 光皮桦天然林群落优势种群的种间联结性研究. 应用生态学报, 2001, 12(2): 168- 170.
[6] 林永慧, 何兴兵, 胡文勇, 田启建, 何平. 濒危植物缙云卫矛群落种间联结分析. 植物科学学报, 2011, 29(1): 58- 63.
[7] 郭忠玲, 马元丹, 郑金萍, 刘万德, 金哲峰. 长白山落叶阔叶混交林的物种多样性、种群空间分布格局及种间关联性研究. 应用生态学报, 2004, 15(11): 2013- 2018.
[8] 孙中伟, 赵士洞. 长白山北坡椴树阔叶红松林群落木本植物种间联结性与相关性研究. 应用生态学报, 1996, 7(1): 1- 5.
[9] Auyang S Y. Foundations of Complex-system Theories. USA: Cambridge University Press, 1998: 7- 9.
[10] Albert R, Jeong H, Barabási A L. Error and attack tolerance of complex networks. Nature, 2000, 406(6794): 378- 382.
[11] Luscombe N M, Babu M M, Yu H, Snyder M, Teichmann S A, Gerstein M. Genomic analysis of regulatory network dynamics reveals large topological changes. Nature, 2004, 431(7006): 308- 312.
[12] Williams R J, Berlow E L, Dunne J A, Barabási A L, Martinez N D. Two degrees of separation in complex food webs. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99(20): 12913- 12916.
[13] Montoya J M, Solé R V. Small world patterns in food webs. Journal of Theoretical Biology, 2002, 214(3): 405- 412.
[14] Montoya J M, Pimm S L, Solé R V. Ecological networks and their fragility. Nature, 2006, 442(7100): 259- 264.
[15] Pires M M, Guimarães P R Jr, Araújo M S, Giaretta A A, Costa J C, dos Reis S F. The nested assembly of individual-resource networks. The Journal of Animal Ecology, 2011, 80(4): 896- 903.
[16] 郑元润. 大青沟森林植物群落主要木本植物种群分布格局及动态的研究. 植物学通报, 1998, 15(6): 52- 58.
[17] 张炜银, 李鸣光, 梁士楚, 王伯荪, 昝启杰, 王勇军. 外来杂草薇甘菊种群分布格局研究. 广西植物, 2003, 23(4): 303- 306.
[18] Diamond J M, Gilpin M E. Examination of the “null” model of Connor and Simberloff for species co-occurrences on islands. Oecologia, 1982, 52(1): 64- 74.
[19] Gotelli N J, McCabe D J. Species co-occurrence: a meta-analysis of J. M. Diamond′s assembly rules model. Ecology, 2002, 83(8): 2091- 2096.
[20] 苏日古嘎, 张金屯, 程佳佳, 张斌. 东灵山糠椴林优势种种群结构与分布格局. 生态学杂志, 2009, 28(7): 1253- 1258.
[21] Greig-Smith P. Pattern in vegetation. Journal of Ecology, 1979, 67(3): 755- 779.
[22] 金则新. 浙江天台山七子花种群结构与分布格局研究. 生态学杂志, 1997, 16(4): 15- 19.
[23] 张文辉, 王延平, 康永祥, 刘祥君. 太白山太白红杉种群空间分布格局研究. 应用生态学报, 2005, 16(2): 207- 212.
[24] 张继义, 赵哈林. 科尔沁沙地草地植被恢复演替进程中群落优势种群空间分布格局研究. 生态学杂志, 2004, 23(2): 1- 6.
[25] 李先琨, 黄玉清, 苏宗明. 元宝山南方红豆杉种群分布格局及动态. 应用生态学报, 2000, 11(2): 169- 172.
[26] 金则新. 浙江仙居俞坑森林群落优势种群结构与分布格局研究. 武汉植物学研究, 2000, 18(5): 383- 389.
[27] May R M C. Stability and Complexity in Model Ecosystems. USA: Princeton University Press, 2001: 347- 352.
[28] Watts D J, Strogatz S H. Collective dynamics of ‘small-world’ networks. Nature, 1998, 393(6684): 440- 442.
[29] Barabási A L, Albert R. Emergence of scaling in random networks. Science, 1999, 286(5439): 509- 512.
[30] 方爱丽, 高齐圣, 张嗣瀛. 引文网络的幂律分布检验研究. 统计与决策, 2007, (14): 22- 25.
[31] Odum E P. 生态学基础. 北京:人民教育出版社, 1981: 89- 94.
[32] Wagner A, Fell D A. The small world inside large metabolic networks. Proceedings of the Royal Society of London. Series B: Biological Sciences, 2001, 268(1478): 1803- 1810.
[33] Smith S A, Beaulieu J M. Life history influences rates of climatic niche evolution in flowering plants. Proceedings of the Royal Society B: Biological Sciences, 2009, 276(1677): 4345- 4352.
Network structure of plant diversity of Dongling Mountain, Beijing
CHEN Yuzhou1,2, MA Keming1,*, ZHANG Yuxin1, ZHANG Shuang1, NIU Shukui2
1StateKeyLaboratoryofUrbanandRegionalEcology,ResearchCenterforEco-EnvironmentalScience,ChineseAcademyofSciences,Beijing100085,China2TheKeyLaboratoryforSilvicultureandConservationofMinistryofEducation,BeijingForestryUniversity,Beijing100083,China
Community can be regard as a complex system which is built with different connecting species. Exploring the pattern of the structure is the basis of system modeling, which contributes to our understanding of the system property and the relationship between structure and function of the system. Complex network theory has been used in studying food webs and mutualistic networks, but the use of network theory in describing the pattern of forest plant diversity hasn′t been tested. In this study, using the network theory, we analyzed the network structure of species diversity pattern of woody, shrub, and herb in a warm temperate deciduous broad-leaved forest of Dongling Mountain, Beijing. The results showed that the pattern of species co-occurrence is non-random, showed an so called “small world” pattern. The co-occurrence species networks of herb, shrub and wood are significant different. The net work for herb is looser, it has longer average path length than the shrub and wood layer. These results indicate that the network theory is useful in analyzing the diversity pattern of different layers of species among associated communities.
communities associate; distribution pattern; complex network; co-occurrence species
国家自然科学基金青年基金(31300368)
2013- 09- 25;
2014- 08- 22
10.5846/stxb201309252357
*通讯作者Corresponding author.E-mail: mkm@rcees.ac.cn
陈禹舟, 马克明, 张育新, 张霜, 牛树奎.北京东灵山森林植物多样性的网络结构特征.生态学报,2015,35(11):3702- 3709.
Chen Y Z, Ma K M, Zhang Y X, Zhang S, Niu S K.Network structure of plant diversity of Dongling Mountain, Beijing.Acta Ecologica Sinica,2015,35(11):3702- 3709.
猜你喜欢
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
建筑与预算(2022年6期)2022-07-08
农村实用技术(2020年1期)2020-12-16
蔬菜(2018年5期)2018-05-17
高原山地气象研究(2016年1期)2016-11-10
浙江大学学报(工学版)(2016年9期)2016-06-05
杂草学报(2015年2期)2016-01-04
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
河南科技(2014年16期)2014-02-27
测绘科学与工程(2014年6期)2014-02-27
