
文峪河上游华北落叶松林的种子雨、种子库与幼苗更新
2015-02-07 05:50高润梅石晓东郭跃东樊兰英
生态学报 2015年11期
高润梅,石晓东,郭跃东,樊兰英
1 山西农业大学林学院, 太谷 030801 2 山西省林业科学研究院, 太原 030012
文峪河上游华北落叶松林的种子雨、种子库与幼苗更新
高润梅1,*,石晓东1,郭跃东1,樊兰英2
1 山西农业大学林学院, 太谷 030801 2 山西省林业科学研究院, 太原 030012
华北落叶松林下更新不良,为探究其制约因素,开展了山西省文峪河上游5个华北落叶松林分的种子雨、土壤种子库和幼苗更新的研究。结果表明:(1) 华北落叶松种子主要集中于9—10月散落。2011年为华北落叶松种子丰年:种子产量高,种子雨密度达(961.93±377.40)粒/m2;种子质量高,完整种子占(89.31±16.13)%。2012年为种子平年,种子产量低,种子雨密度为(252.73±115.12)粒/m2。华北落叶松种子雨主要源于毗邻树木,华北落叶松纯林和落叶松云杉林的种子雨密度显著高于其他3个针阔混交林。(2) 土壤种子库主要由上年种子雨组成,2012年4月的土壤种子库密度为(695.18±297.23)粒/m2,完整种子占(59.73±9.56)%。种子自然萌发前,约(78.98±24.76)粒/m2具发芽力,基本可满足更新需要。但种子活力保持期少于2 a,只能形成短期持久土壤种子库。(3) 华北落叶松更新不良,种子年后仍难以实现幼苗建成,当年生幼苗的出现频度平均为1.6%,且林下难以存活。幼苗发生与种子储量关联性不强,种源条件不是制约华北落叶松更新的主要因素。
更新; 华北落叶松; 种子雨; 土壤种子库; 文峪河
自然条件下,多数针叶树种通过种子实现更新,足够的种源是林木实现更新的必备条件之一。在更新动态中,种子雨的生产、扩散和土壤种子库动态等环节都可能成为更新的限制因素[1- 3]。落叶松属(Larix)已有的更新研究表明,低劣的种子质量是造成美国西北部与加拿大西部L.occidentalis天然更新失败的主要原因[4];蒙古Khentey山脉西部的新疆落叶松(L.sibirica)更新困难,与其母树缺少球果有关[5];兴安落叶松(L.gmelinii)与长白落叶松(L.olgensis)的种子仅能保持1a的生活力,难以形成连续的土壤种子库[6- 8]。种子雨与土壤种子库的可靠调查与评估在一定程度上有助于有效预测现有群落的潜在更新能力[9- 10]。
华北落叶松(L.principis-rupprechtii)是华北地区中高山寒温性针叶林的主要建群种之一,也是重要的用材林、水源涵养林和景观风景林树种,具有较高的利用和保护价值。山西省是华北落叶松最适生长区,构成了我国华北落叶松的主体,对于维持区域的景观多样性,维护该区及下游地区的环境质量具有举足轻重的作用。然而,该种林下更新不良,幼苗幼树缺乏[11- 12]。虽已有该区华北落叶松土壤种子库的报道[13- 14],但缺乏对其种子雨和种子库输出动态—幼苗萌发的追踪报道,而已有研究表明,落叶松可在种子年后实现幼苗建成[15],因此,本文拟通过开展华北落叶松种子雨、种子年后土壤种子库及幼苗发生等的研究,探寻幼苗更新与种源条件之间的可能联系,以期有效预测现有华北落叶松群落的更新潜力与未来发展方向,为经营、管理与保护现有华北落叶松天然林提供更充分的理论依据。
1 研究地区自然概况
文峪河上游位于山西省吕梁山脉中段的庞泉沟国家级自然保护区,地理位置111°21′—111°37′E,37°45′—37°59′N,本区属暖温带大陆性山地气候,年均温4.2 ℃,年均降水量822.6 mm,6—8月3个月的降水量占全年降水量的59.4%,年均相对湿度70.9%,年均蒸发量1100—1500 mm,无霜期年际变幅较大,平均为100—130 d;年日照时数1900—2200 h。地带性土壤为山地棕壤。
华北落叶松是构成该区天然次生林的主要建群种,在海拔高于1800 m的亚高山地带形成纯林,或与云杉(Piceawilsonii,P.meyeri)混交形成以华北落叶松为主的落叶松云杉混交林,或于山脊线同白桦(Betulaplatyphylla)、山杨(Populusdavidiana)或红桦(B.albo-sinensis)等混交。林下灌木主要有土庄绣线菊(Spiraeapubescens)、山刺玫(Rosadavurica)、金花忍冬(Lonicerachrysantha)、陕西荚蒾(Viburnumschensianum)、毛榛(Corylusmandshurica)等,盖度30%;草本主要有披针苔草(Carexlanceolata)、小红菊(Dendranthemachanetii)、细叶鸢尾(Iristenuifolia)、东方草莓(Fragariaorientalis)、鹿蹄草(Pyrolacalliantha)、花荵(Polemoniumcoeruleum)、蒿类(Artemisiaspp.)、禾草类(Poaspp.)等,盖度90%。
2 研究方法
2.1 样地设置
根据已有的调查数据和航片资料,结合实地踏查,在研究地区选择5种林龄约60年的华北落叶松林设置样地,面积为100 m×100 m。对每个样地进行立木调查,并记录样地的立地条件。各林分概况见表1。

表1 林分概况
2.2 种子雨
华北落叶松球果9—10月成熟,2011年8月在华北落叶松种子自然散播之前,分别在以上5个林分沿“W”型曲线随机设立15个接种点[16],为尽可能减少动物对种子雨结果的影响,种子收集器离地1.4—1.5 m。每个收集器框口面积为50 cm × 50 cm,框口四边以1.5 cm宽的等长木条固定,尼龙网(网眼2 mm × 2 mm)铺置网底。四角拧设细铁环,牵引尼龙绳,分别绑缚在毗邻的树干上,以固定种子收集器。内放大小适中的石块,以防种子落下后反弹出框或被风吹走。
2011—2013年8月10日至11月30日,每10 d收集1次种子。先将收集器内的枯枝落叶和其它植物的种子等杂物分离出去,按完整粒(种子饱满、颜色鲜明、无啃食痕迹)、被取食粒(种子有明显啃食痕迹)和干瘪与腐烂粒等3类分别统计,后两类记为不完整种子[7]。同时,分离收集器中的落叶并称重,用于分析种子雨落种过程与其枯叶降落量之间的关系。2012—2013年产种量小,只对种子雨总量进行统计。
2.3 土壤种子库
2012年4月下旬,在上述5个林分毗邻每个接种点设置 20 cm × 20 cm 的小样方,进行土样采集。多数落叶松种子留存于枯落物层,受动物活动与地表径流等影响,小部分种子向下发生迁移[7],但10 cm的取样深度足以反映森林土壤种子库的储量[16],同时借鉴已有的落叶松种子库的取样方法[7],区分枯落物层、0—5 cm和 5—10 cm的土壤层采集土样,分别装入小布袋,共计225个土样。土样带回室内人工挑拣华北落叶松种子,按照种子雨的分类标准,分别记录完整粒、被取食粒和干瘪与腐烂粒的数量[7]。
将每种林分挑选的完整种子分为两份,一份用于室内发芽试验进行活力测定;另一份区分样方装袋后,于5月中旬埋于原有林分,埋土深度约5 cm,上面覆盖与毗邻环境厚度相近的枯落物[16]。2013年4月,取出布袋,同法测定种子活力。种子发芽试验在山西农业大学林学院苗圃温室进行,取林下土壤装入直径约20 cm的圆形发芽盘,浇湿,均匀播撒落叶松种子后,覆盖约1 cm的林土,隔天洒水,以保持土壤湿润。光源为高压钠灯(400 W),光/暗周期12/12 h, 光周期23.5—25 ℃;暗周期18.5—20 ℃。发芽盘每周调换两次位置,以减弱发芽盘位置对发芽的影响作用。每天9:00—10:00时观察记录萌发幼苗的数量,持续5d无幼苗出现时结束试验。
2.4 林下更新幼苗
2012年5月在上述林分每个土样采集点附近设置3—4个50 cm × 50 cm的小样方,每个林分设置50个小样方。6—9月每月定期调查华北落叶松当年生幼苗的数量、频度、地径与株高等。幼茎尚未木质化或木质化程度弱的幼苗被计为当年生幼苗[2]。
2.5 数据处理
采用SPSS11.5软件对不同林分的种子雨、种子库储量及幼苗数量等的差异性进行单因素方差分析(one-way ANOVA)。在方差分析前,对分析数据进行方差齐性检验(Levene-test),以邓肯(Duncan)多重检验法检验数据之间的差异显著性,显著水平P=0.05;进行种子雨密度与落叶量、种子雨千粒重与完整种子比例、种子雨总量与土壤种子库密度、当年生幼苗数量与种子库完整种子之间的相关性分析,显著水平P=0.01。
3 结果与分析
3.1 种子雨
8—11月定期收集华北落叶松种子,室内人工挑拣后,分析种子雨扩散过程、质量及总量年际变化。结果如下:
3.1.1 种子雨扩散
由图1可知,华北落叶松8月下旬开始落种,9月中旬至10月中旬为落种高峰期,11月下旬结束。根据散种量的差异,可将种子扩散过程分为3个阶段:(1)起始期:8月20日至9月10日,散种量较小,平均为(3.24±3.82)粒 m-2d-1;(2)高峰期:9月20日至10月20日,散种量急剧上升,平均为(19.14±10.64)粒 m-2d-1,9月末至10月初增至(24.62±11.95)粒 m-2d-1,后散种量逐渐下降;(3)末期:10月30日至11月20日,散种量减少至停止,平均为(3.28±3.08) 粒 m-2d-1。
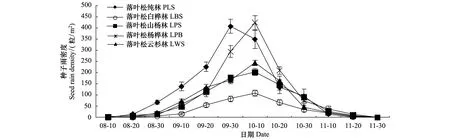
图1 2011年华北落叶松种子雨散落动态
落叶动态见图2。由图2可知,枯叶降落量在种子雨扩散期也呈现先增后减的趋势,9月中旬至10月中旬落叶量较大,平均为(1.80±0.73)g m-2d-1,此后逐渐减少,与种子雨的散落高峰期高度吻合。华北落叶松种子雨密度与其落叶量显著相关(r=0.91,P<0.01)。
3.1.2 种子雨质量
由表2知,华北落叶松种子雨以完整种子居多,平均占种子雨总量的(89.31±16.13)%。在散种过程中,完整种子的比例先升后降:起始期为(73.97±20.68)%;高峰期达(97.41±2.14)%;散落末期,这一比例下降为(77.78±24.40)%。高峰期的完整种子比例显著高于起始期和末期(P<0.05)。

图2 2011年落叶动态

表2 华北落叶松2011年不同散种期的完整种子比例/%(平均值±标准差)
由表3知,华北落叶松种子雨的千粒重平均为(4.92±1.08)g。在种子散落过程中,种子千粒重先升后降:起始期最小,为(3.61±0.71)g;高峰期达(5.71±0.64)g;末期降至(3.95±0.88)g,高峰期的种子雨千粒重显著高于起始期和末期(P<0.05)。
种子雨千粒重随散种期的变化趋势与完整种子比例相似,相关分析表明,二者显著相关(r=0.44,P<0.01)。

表3 华北落叶松2011年不同散种期的千粒重/g(平均值±标准差)
3.1.3 种子雨总量
由表4知,2011年的华北落叶松种子雨总量为(961.93±377.40)粒/m2,完整种子(867.73±358.26)粒/m2,约占种子雨的(89.63±16.13)%;2012年的种子雨总量为(252.73±115.12)粒/m2,完整种子(91.07±45.78)粒/m2,约占种子雨的(36.04±4.26)%;2011年的种子雨总量和完整种子数量均极显著高于2012年(P<0.01)。2013年基本没有收集到华北落叶松种子,为种子歉年。
落叶松纯林和落叶松云杉林的种子雨总量显著高于其他3个针阔混交林(P<0.05,表4)。各林分的种子雨大小与林分落叶松密度成显著正相关(r=0.98,P<0.01)。华北落叶松的不完整种子主要为发育不良或败育的干瘪粒,2011年不完整种子的比例低于10%,2012年这一比例为60%—70%。

表4 华北落叶松2011与2012年种子雨/(粒/m2)(平均值±标准差)
3.2 土壤种子库
2012年4月下旬雪融后在5个林分采集土样,进行土壤种子库分析.
3.2.1 种子库密度
由表5知,2012年的华北落叶松土壤种子库密度在不同林分间存在显著差异(P<0.05):落叶松纯林和落叶松云杉林的种子库储量较大,分别为(1041.00±98.15)粒/m2和(1063.67±215.51)粒/m2;落叶松白桦林和落叶松山杨林较接近,分别为(507.60±75.04)粒/m2和(615.00±39.15)粒/m2;落叶松杨桦林的土壤种子库密度最低,为(373.67±46.48)粒/m2。5 个林分的土壤种子库密度平均为(695.18±297.23)粒/m2,约占种子雨总量的(76.05±13.79)%,二者紧密相关(r=0.88,P<0.01)。
土壤种子库的完整种子数为(415.24±176.62)粒/m2,占种子库总量的(59.73±9.56)%;不完整种子占(40.03±9.56)%,其中干瘪与腐烂种子(57.06±58.89)粒/m2,被取食种子(222.88±108.46)粒/m2,根据被取食种子的种壳形状和啃食痕迹,推测取食动物主要为鼠类。多于85%的种子留存于枯落物层,5.50—12.87%的种子出现于0—5 cm的土壤层;5—10 cm土壤层的留种量极小,仅有0.09—0.47%。

表5 2012年华北落叶松土壤种子库密度/(粒/m2)(平均值±标准差)
3.2.2 种子库完整种子活力
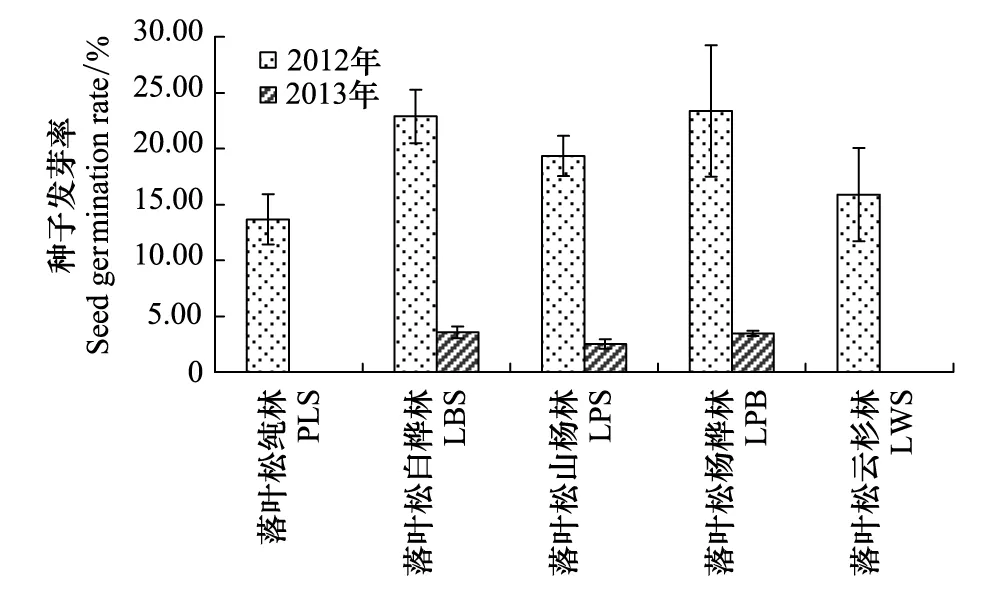
图3 华北落叶松2012年种子库完整种子的发芽率
2012年4月完整种子的发芽率平均为(19.02±5.00)%(图3),即约(78.98±24.76)粒/m2具发芽潜力;针阔混交林完整种子的发芽率显著高于落叶松纯林和落叶松云杉林(P<0.05)。完整种子在林下土壤继续留存1年,至2013年4月时,种子发芽率大幅降低。落叶松纯林和落叶松云杉林的种子库没有种子萌发,其余3个林分种子的萌发率降至(1.90±0.57)%(图3),即约(7.89±2.53)粒/m2种子具萌发力。
3.3 幼苗更新
2012年6—9月对5个华北落叶松林分进行更新调查。结果表明,当年生幼苗6月开始出现,至7月底,共2个林分4个样方发现10株当年生幼苗:落叶松白桦林8株,出现频度6%;落叶松杨桦林2株,出现频度2%;5个林分当年生幼苗的出现频度平均为1.6%,此后幼苗数量不再增加,至9月底,10株幼苗全部死亡。挖取死亡幼苗观察,发现死亡幼苗的根茎相接处缢缩变黑,推测为幼苗水分缺失所致。由于幼苗数量少,难以进行其与种子雨或种子库的相关分析,但10株幼苗并非发生于种子储量较为丰富的林分,足以说明幼苗发生与种子储量关联性不强。由此可知,华北落叶松种子年后仅出现少量幼苗,且林下不能正常生长与存活,难以实现幼苗建成。
4 讨论与结论
4.1 讨论
4.1.1 华北落叶松种子雨
华北落叶松种子雨从8月下旬持续至11月中旬,多数种子集中于9—10月散落,散种时间与同属的长白落叶松相似[7]。扩散高峰期的种子质量高于起始期和末期,利用前期的低劣种子消耗取食者的部分取食潜力,以降低高峰期优质种子被捕食的风险;且种子扩散动态与落叶动态高度吻合,通过落叶覆盖,降低种子被动物取食的可能性。这是多数树木不断适应环境而形成的种子扩散策略和自我繁殖保护机制[2]。
落叶松纯林和落叶松云杉林的种子雨密度高于其他3个针阔混交林林分。华北落叶松种子散落量与林分母树密度显著正相关,而且郁闭林冠可以有效拦截种子[17- 18]。此外,虽然落叶松种子小且具翅,可以随风扩散到林冠外,但由于受自身重力影响,多数种子主要降落在林冠下;且自然条件下,落叶松种子的1次扩散距离最远为树高的1.5倍,即距离母树约16—32 m之处[7,19],因此收集器内的种子主要源于毗邻的多株母树[13,20]。针阔混交林中,相当比例的阔叶树不仅对落叶松种子雨毫无贡献,且已进入主林层的庞大树冠和枝叶会阻挡落叶松种子扩散,进而缩短扩散距离,相对减少落叶松种子来源,因此针阔混交林的华北落叶松种子雨密度相对较低。
落叶松属结实有丰歉年之分,丰歉年的产种量差异较大,一般认为种子丰歉年的周期为3—5 a[4,7- 8]。根据2011—2013年种子雨的年际差异判断,2011年是研究地区华北落叶松结实丰年,种子产量高:种子雨密度平均达(961.93±377.40)粒/m2;种子质量高:千粒重(4.92±1.08)g,完整种子达(867.73±358.26)粒/m2,占种子雨的89.31%。2012年为华北落叶松种子平年,种子产量低:种子雨密度为(252.73±115.12)粒/m2;种子质量低劣:完整种子(91.07± 45.78)粒/m2,约占种子雨的36.03%。2013年没有收集到落叶松种子,为种子歉年。种子年的发生是不少树种实现更新的先决条件,多数树木只有在种子年后才能实现幼苗建成[3,15,21]。
4.1.2 华北落叶松土壤种子库
2012年4月下旬的土壤种子库主要源于2011年的种子雨。因此,5个林分的种子库储量排序与种子雨相同:落叶松纯林与落叶松云杉林的种子库储量最大,多于1000 粒/m2;落叶松红桦林和落叶松山杨林居中,为500—900 粒/m2;落叶松杨桦林最低,为400—450 粒/m2。华北落叶松的土壤种子库密度平均为(695.18±297.23)粒/m2,其中完整种子(415.24±176.62) 粒/m2,与郭晋平的研究结果相近[13],但显著高于2006年的种子库[14],这在于土壤种子库间的年际差异[16],2005年可能为种子歉年。
多于85%的华北落叶松种子留存于枯落物层,0—5 cm的土壤层留存有5.50%—12.87%的种子,土壤层5 cm以下基本没有种子。与其它树种的研究结果相似[7,22],大量的落叶松种子散落后,难以穿越林下枯枝落叶层或根系盘结层构成的隔离带;仅有少量种子在自身重力、下渗水以及动物储食等的作用下逐渐下移,到达土壤表层[7]。华北落叶松种子库消耗以动物取食为主,占种子库总量的32.06%,取食动物主要为鼠类,与对长白落叶松[7]和兴安落叶松[15]的研究结果相似。研究地区冬春季节降水少,林地环境干燥,仅有少量的华北落叶松种子(<8.21%)发生腐烂。
华北落叶松种子年后的种源不是其更新不良的限制因素,种子库室内萌发试验表明,华北落叶松种子年后的翌年春季,在种子自然萌发前,约(78.98±24.76) 粒/m2种子具萌发力,基本可满足更新需要,与兴安落叶松[6]和长白落叶松[7]的研究结果一致。但种子在林下土壤中继续留存1年后,最多有(7.89±2.53)粒/m2可在室内萌发。土壤种子库研究工作中,大约90%采用萌发法[23],但由于室内和林地环境的巨大差异,萌发法所得结果可能不同于林地萌发率。种子发芽主要依赖于水分的干湿循环[24],充足的水分是土壤中种子萌发的必要因素[25- 26]。研究地区冬春季节降水少,5月平均降水量小于75 mm,推断自然生境下华北落叶松种子的萌发率和活力保持期将低于室内萌发法的结果。因此,华北落叶松种子的活力持续期少于2 a,只能形成短期持久种子库[14],与云杉属、冷杉属(Abies)和水青冈属(Fagus)等的研究结果相似[9,14,20- 22]。
4.1.3 华北落叶松幼苗更新及可能影响因素
足够的种源是林木实现更新的基本条件之一,华北落叶松种子年后的翌年春季,留存于林地的活力种子基本可以满足更新需要,但当年生幼苗发生率低,出现频度平均为1.6%,且幼苗在林下不能正常存活,难以实现幼苗建成,与对长白落叶松的研究结果一致[7,27]。该区华北落叶松天然更新种群结构与空间格局的研究也表明,5个20 m×20 m的样地中,4个样地缺失幼苗(DBH≤2.5 cm),仅1个样地发现5株幼苗[12]。这些研究反映了华北落叶松天然更新不良的客观现实,并将导致该种发展为衰退种群[11- 12],森林面积不断减少,引发逆行演替,使华北落叶松从原有分布区彻底消失[5]。
除了种源条件,种子萌发与幼苗存活的适宜生境也是影响更新的重要因素[8,16]。水分是种子发芽的直接驱动力[25- 26],针叶树种遭遇夏季干旱等气候变化时难以实现更新定居[28]。北方森林中,新疆落叶松的种子萌发对土壤干旱高度敏感[29],山地森林林下幼苗完全缺失,而在泛滥平原(flood plain)仍有更新[5]。研究地区近50 a以来干旱频发[30],林木种子萌发率降低[31]。因此,干燥的林地环境可能是限制华北落叶松种子萌发的主要原因。另外,林下枯枝落叶层阻断种子与土壤的有效接触,也是树种天然更新最直接的障碍因素[32- 33],对落叶松等小粒种子的抑制作用更加明显[27,34]。该区华北落叶松林的年凋落物总量为19.15 t hm-2a-1,分解半衰期1.5—2 a[35],而本研究表明,自然萌发前,多于85%的华北落叶松种子仍留存于枯落物层,因此即使部分种子得以萌发,幼苗根系由于枯落物阻隔,也难以深入土壤,养分和水分供给困难,导致幼苗死亡[33]。这可以从本研究中10株当年生幼苗全部死亡的事实得到佐证。今后应开展林下生境,尤其是土壤水分条件和枯落物阻断对华北落叶松更新影响的研究。
4.2 结论
通过对文峪河上游5个华北落叶松林分种子雨、土壤种子库和幼苗更新的研究,得到如下结论:
(1) 华北落叶松种子雨从8月下旬持续至11月下旬,主要集中于9—10月,散种期和散种量与枯叶的散落规律一致。种子为枯枝落叶覆盖,将减少被动物取食的机会。2011年种子产量高,密度达(961.93±377.40)粒/m2,完整种子占(89.31±16.13)%,为种子丰年;2012年种子产量低,为种子平年;2013年为种子歉年。
(2) 华北落叶松种子雨主要源于毗邻的多株母树。散种量与林分密度显著正相关,华北落叶松纯林和落叶松云杉林的种子雨密度显著高于其他3个针阔混交林。2012年4月的土壤种子库主要由种子雨组成,华北落叶松纯林和落叶松云杉林的种子库储量也显著高于其他3个针阔混交林。
(3) 2012年4月的土壤种子库密度为(695.18±297.23)粒/m2,完整种子占(59.73±9.56)%。85%以上的种子散落于枯落物层,5.50—12.87%的种子留存于0—5 cm的土壤层,5 cm以下的土壤层基本没有华北落叶松种子。种子自然萌发前,约(78.98±24.76)粒/m2具萌发力,基本可满足更新需要。但林下种子活力持续期少于2 a,仅可形成短期持久土壤种子库。
(4) 华北落叶松林下更新不良,种子年后仍不能实现幼苗建成,仅2个林分出现当年生幼苗,幼苗出现频度平均为1.6%,且林下难以存活。幼苗发生与种子储量关联性不强,种源条件不是制约华北落叶松更新的主要因素。
致谢:感谢加拿大Lakehead University自然资源管理系骆咏博士后对论文写作的帮助。
[1] Batkhuu N O, Lee D K, Tsogtbaatar J, Park Y D. Seed quality of Siberian larch (LarixsibiricaLdb.) from geographically diverse seed sources in Mongolia. Scandinavian Journal of Forest Research, 2010, 25(Suppl 8): 101- 108.
[2] Du X J, Guo Q F, Gao X M, Ma K P. Seed rain, soil seed bank, seed loss and regeneration ofCastanopsisfargesii(Fagaceae) in a subtropical evergreen broad-leaved forest. Forest Ecology and Management, 2007, 238(1/3): 212- 219.
[3] Yin H J, Liu Q, He H. Seed rain and soil seed bank ofPiceaasperataMast. in subalpine coniferous forest of western Sichuan, China. Plant Biosystems, 2007, 141(3): 314- 322.
[4] Stoehr M U. Seed production of western larch in seed-tree systems in the southern interior of British Columbia. Forest Ecology and Management, 2000, 130(1/3): 7- 15.
[5] Dulamsuren C, Hauck M, Khishigjargal M, Leuschner H H, Leuschner C. Diverging climate trends in Mongolian taiga forests influence growth and regeneration ofLarixsibirica. Oecologia, 2010, 163(4): 1091- 1102.
[6] 徐化成, 班勇. 大兴安岭北部兴安落叶松种子在土壤中的分布及其种子库的持续性. 植物生态学报, 1996, 20(1): 28- 34.
[7] 刘足根, 朱教君, 袁小兰, 王贺新, 谭辉. 辽东山区长白落叶松(Larixolgensis)种子雨和种子库. 生态学报, 2007, 27(2): 579- 587.
[8] 朱教君, 刘足根, 王贺新. 辽东山区长白落叶松人工林天然更新障碍分析. 应用生态学报, 2008, 19(4): 695- 703.
[9] Bossuyt B, Honnay O. Can the seed bank be used for ecological restoration? An overview of seed bank characteristics in European communities. Journal of Vegetation Science, 2008, 19(6): 875- 884.
[10] Abella S R, Chiquoine L P, Vanier C H. Characterizing soil seed banks and relationships to plant communities. Plant Ecology, 2013, 214(5): 703- 715.
[11] 石晓东, 韩有志. 庞泉沟自然保护区寒温性针叶林的种群结构与分布格局. 武汉植物学研究, 2008, 26(5): 489- 494.
[12] 武小钢, 郭晋平. 关帝山华北落叶松天然更新种群结构与空间格局研究. 武汉植物学研究, 2009, 27(2): 165- 170.
[13] 郭晋平, 薛俊杰, 李世广, 王俊田. 庞泉沟自然保护区华北落叶松土壤种子库的研究. 武汉植物学研究, 1998, 16(2): 131- 136.
[14] 高润梅, 郭晋平, 郭跃东. 山西文峪河上游河岸林的土壤种子库与树种更新特征. 植物科学学报, 2011, 29(5): 580- 588.
[15] 班勇, 徐化成. 原始老龄林内兴安落叶松种子命运的试验研究. 生态学报, 1996, 16(5): 541- 547.
[16] Csontos P. Seed banks: ecological definitions and sampling considerations. Community Ecology, 2007, 8(1): 75- 85.
[17] de Andres E G, Camarero J J, Martinez I, Coll L. Uncoupled spatiotemporal patterns of seed dispersal and regeneration in Pyrenean silver fir populations. Forest Ecology and Management, 2014, 319: 18- 28.
[18] Paluch J G. Ground seed density patterns under conditions of strongly overlapping seed shadows inAbiesalbaMill. stands. European Journal of Forest Research, 2011, 130(6): 1009- 1022.
[19] Yan Q L, Zhu J J, Gang Q. Comparison of spatial patterns of soil seed banks between larch plantations and adjacent secondary forests in Northeast China: implication for spatial distribution of larch plantations. Trees, 2013, 27(6): 1747- 1754.
[20] Berger T W, Sun B, Glatzel G. Soil seed banks of pure spruce (Piceaabies) and adjacent mixed species stands. Plant and Soil, 2004, 264(1/2): 53- 67.
[21] Rossi S, Morin H, Gionest F, Laprise D. Episodic recruitment of the seedling banks in balsam fir and white spruce. American Journal of Botany, 2012, 99(12): 1942- 1950.
[22] 尹华军, 刘庆. 川西米亚罗亚高山云杉林种子雨和土壤种子库研究. 植物生态学报, 2005, 29(1): 108- 115.
[23] 李洪远, 莫训强, 郝翠. 近30年来土壤种子库研究的回顾与展望. 生态环境学报, 2009, 18(2): 731- 737.
[24] Leck M A, Brock M A. Ecological and evolutionary trends in wetlands: evidence from seeds and seed banks in New South Wales, Australia and New Jersey, USA. Plant Species Biology, 2000, 15(2): 97- 112.
[25] Grombone-Guaratini M T, Leitao Filho H F, Kageyama P Y. The seed bank of a gallery forest in Southeastern Brazil. Brazilian Archives of Biology and Technology, 2004, 47(5): 793- 797.
[26] Walck J L, Hidayati S N, Dixon K W, Thompson K, Poschlod P. Climate change and plant regeneration from seed. Global Change Biology, 2011, 17(6): 2145- 2161.
[27] Zhu J J, Yang K, Yan Q L, Liu Z G, Yu L Z, Wang H X. Feasibility of implementing thinning in even-agedLarixolgensisplantations to develop uneven-aged larch-broadleaved mixed forests. Journal of Forest Research, 2010, 15(1): 70- 81.
[28] Kazantseva O, Bingham M, Simard S W, Berch S M. Effects of growth medium, nutrients, water, and aeration on mycorrhization and biomass allocation of greenhouse-grown interior Douglas-fir seedlings. Mycorrhiza, 2009, 20(1): 51- 66.
[29] Dulamsuren C, Wommelsdorf T, Zhao F J, Xue Y Q, Zhumadilov B Z, Leuschner C, Hauck M. Increased summer temperatures reduce the growth and regeneration ofLarixsibiricain southern boreal forests of eastern Kazakhstan. Ecosystems, 2013, 16(8): 1536- 1549.
[30] 马洁华, 刘园, 杨晓光, 王文峰, 薛昌颖, 张晓煜. 全球气候变化背景下华北平原气候资源变化趋势. 生态学报, 2010, 30(14): 3818- 3827.
[31] 董丽佳, 桑卫国. 模拟增温和降水变化对北京东灵山辽东栎种子出苗和幼苗生长的影响. 植物生态学报, 2012, 36(8): 819- 830.
[32] Bueno A, Baruch Z. Soil seed bank and the effect of needle litter layer on seedling emergence in a tropical pine plantation. Revista de Biologia Tropical, 2011, 59(3): 1071- 1079.
[33] 陈永富. 森林天然更新障碍机制研究进展. 世界林业研究, 2012, 25(2): 41- 45.
[34] 刘足根, 朱教君, 袁小兰, 谭辉. 辽东山区长白落叶松天然更新调查. 林业科学, 2007, 43(1): 42- 49.
[35] 郭晋平, 丁颖秀, 张芸香. 关帝山华北落叶松林凋落物分解过程及其养分动态. 生态学报, 2009, 29(10): 5684- 5695.
Seed rain, soil seed bank and regeneration ofLarixprincipis-rupprechtiistands in the Upper Reaches of Wenyuhe
GAO Runmei1,*, SHI Xiaodong1, GUO Yuedong1, FAN Lanying2
1ForestryCollegeofShanxiAgriculturalUniversity,Taigu030801,China2ShanxiAcademyofForestrySciences,Taiyuan030012,China
Larixprincipis-rupprechtiiis one of dominant deciduous coniferous tree species in subalpine temperate forests of North China. It plays significant roles in water conservation, ecotourism and biodiversity maintenance in its distributional areas. It is also one of major economic tree species in this region due to its fast growth and good-quality timber. Previous studies suggest thatL.principis-rupprechtiiexperiences natural regeneration failure, as indicated by low proportion of saplings and seedlings in natural conditions. The failure of regeneration is a threat to persistence ofL.principis-rupprechtiiforests in its distributional areas. However, the causes of natural regeneration failure are still poorly understood. This study was to examine whether the natural regeneration failure of this species was driven by seed limitation. We investigated seed rain, soil seed bank and seedling regeneration in five typical larch stand types, i.e., pureL.principis-rupprechtiistand,L.rincipis-rupprechtii+Betulaplatyphyllastand,L.principis-rupprechtii+Populusdavidianastand,L.principis-rupprechtii+P.davidiana+B.platyphyllastand,L.principis-rupprechtii+Piceawilsoniistand. Fifteen 50 cm × 50 cm seed traps were deployed in each plot for seed rain collecting. For each plot, 15 quadrats of 20 cm×20 cm with 10cm depth and 50 quadrats of 1 m × 1 m were examined nearby seed traps for soil seed bank and recruitment survey, respectively. We found that most of seeds were dispersed from September to October. In 2011, which was a seed year ofL.principis-rupprechtii, seed production was amounted to (961.93±377.40) seeds/m2, (89.31±16.13)% of which was full seeds. In 2012, a common seed year, both seed production and full seeds proportion were lower than those in previous year, with (252.73±115.12) seeds/m2and (36.04±4.26)%, respectively. Furthermore, seed rain ofL.principis-rupprechtiiwas mainly from the surrounding mother trees. Seed rain densities of pureL.principis-rupprechtiistand andL.principis-rupprechtii+P.wilsoniistand were higher than those of the other three stand types, which were mixed with broad-leave trees. Thirdly, soil seed bank ofL.principis-rupprechtiiwas mainly from the seed rain that produced in previous year. Seed bank in April of 2012 was (695.18±297.23) seeds/m2, accounting for (76.05±13.79)% of seed rain in autumn of 2011. More than 85% of seeds were found in the litter layer. About (59.73±9.56)% of the seed bank was full seeds. There were (78.98±24.76) seeds/m2remained viable to germinate before natural regeneration, accounting for (19.02±5.00)% of full seeds in soil seed bank. The germination capacity decreased with time being in forest floor, hence, forming a short-term persistent soil seed bank forL.principis-rupprechtii. Lastly,the emergence of seedlings uncoupled with the seed storage. The frequency of seedling emergence was 1.6%. Seedlings could hardly survive understory. Collectively, this study demonstrates that seed source, especially in seed year, is not the limiting factor for the poor natural regeneration ofL.principis-rupprechtii. Our study suggests that further studies should be carried out to examine the detrimental or limiting factors, such as effects of soil moisture and litter, for the natural regeneration failure of the species.
regeneration;Larixprincipis-rupprechtii; seed rain; soil seed bank; Wenyuhe
国家自然科学基金项目(31400536); 山西省自然科学基金项目(2012011033- 6);山西农业大学作物学博士后流动站基金项目(125376); 山西农业大学2010年引进人才科研启动项目(2010008)资助
2014- 06- 18;
2014- 11- 27
10.5846/stxb201406181268
*通讯作者Corresponding author.E-mail: sxndgrm@163.com
高润梅,石晓东,郭跃东,樊兰英.文峪河上游华北落叶松林的种子雨、种子库与幼苗更新.生态学报,2015,35(11):3588- 3597.
Gao R M, Shi X D, Guo Y D, Fan L Y.Seed rain, soil seed bank and regeneration ofLarixprincipis-rupprechtiistands in the Upper Reaches of Wenyuhe .Acta Ecologica Sinica,2015,35(11):3588- 3597.
猜你喜欢
南方农业(2021年5期)2021-12-27
今日农业(2021年10期)2021-11-27
今日农业(2021年10期)2021-11-27
今日农业(2020年17期)2020-10-27
山东林业科技(2019年2期)2019-06-03
现代营销(创富信息版)(2018年2期)2018-08-15
大陆桥视野(2016年18期)2016-12-27
大陆桥视野(2016年18期)2016-12-27
中国火炬(2015年12期)2015-07-31
中国火炬(2013年3期)2013-07-24
